Potential Medicinal Plants for the Treatment of Dengue Fever and Severe Acute Respiratory Syndrome-Coronavirus


Abstract
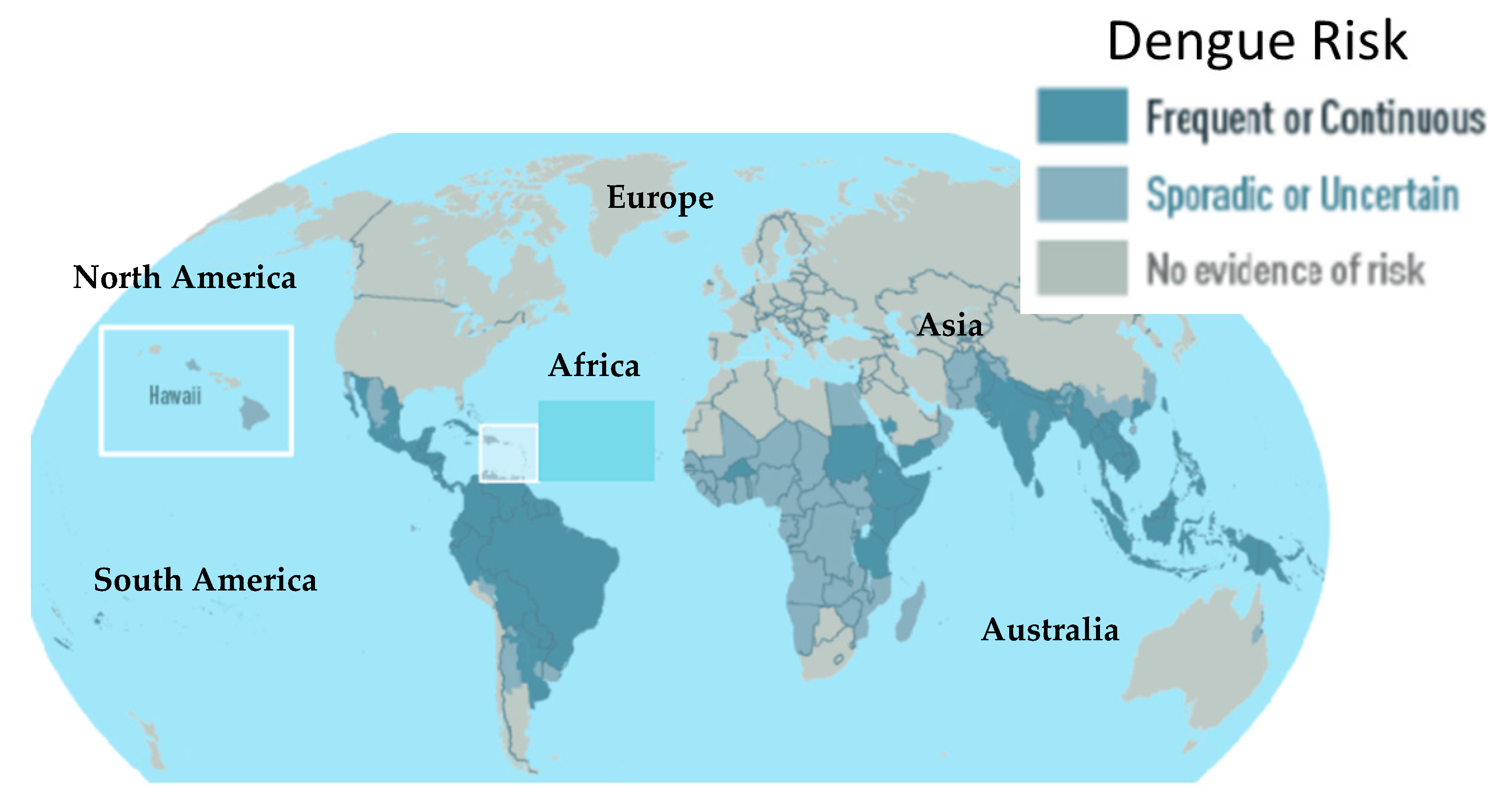
:1. Introduction
2. Evidence Acquisition
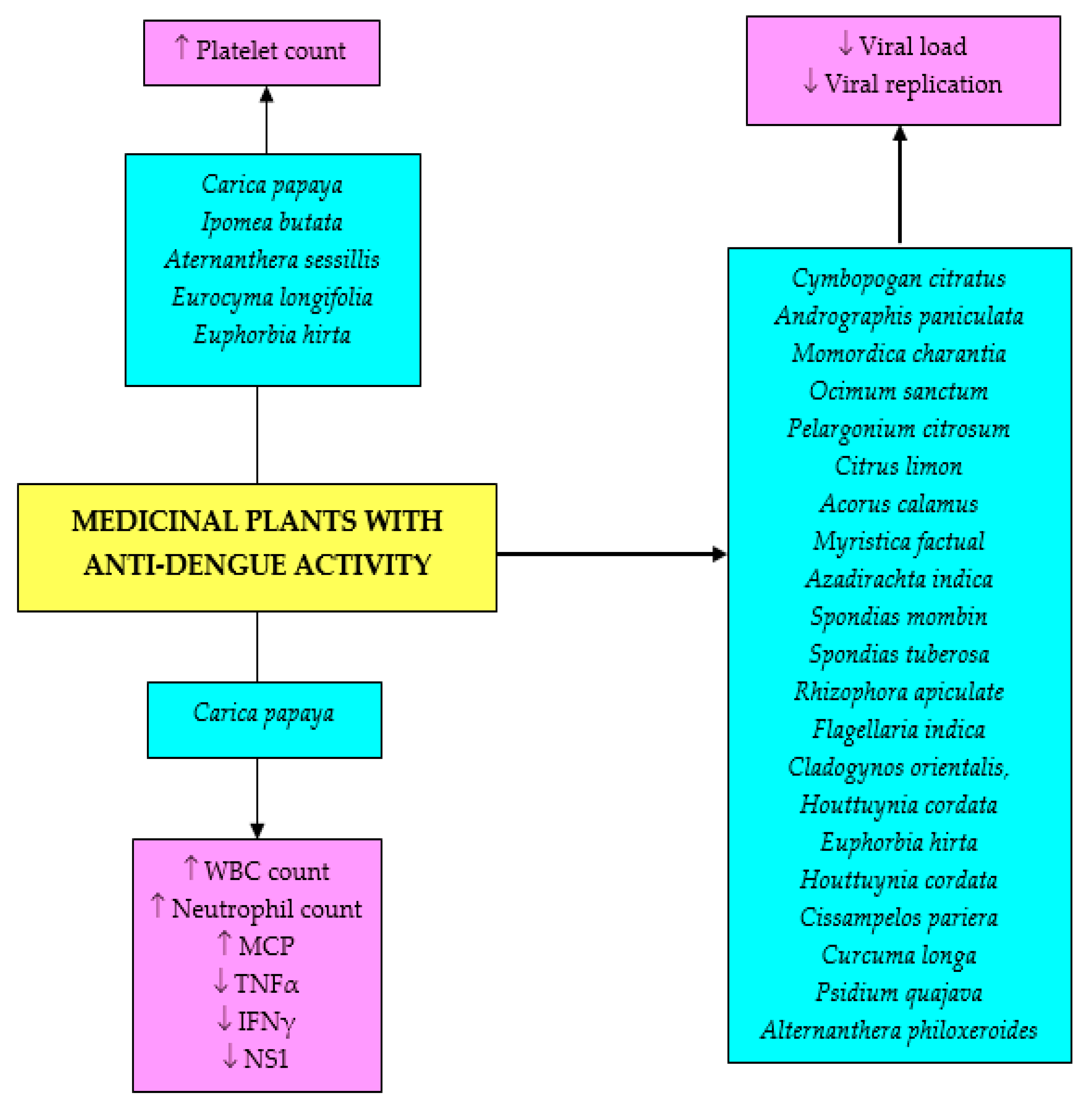
3. Traditional Plants Used for Dengue Treatment
4. Pharmacologically Active Medicinal Plants and Isolated Compounds against Dengue Virus
4.1. In Vivo Studies
4.2. In Vitro Studies
4.3. Clinical Studies
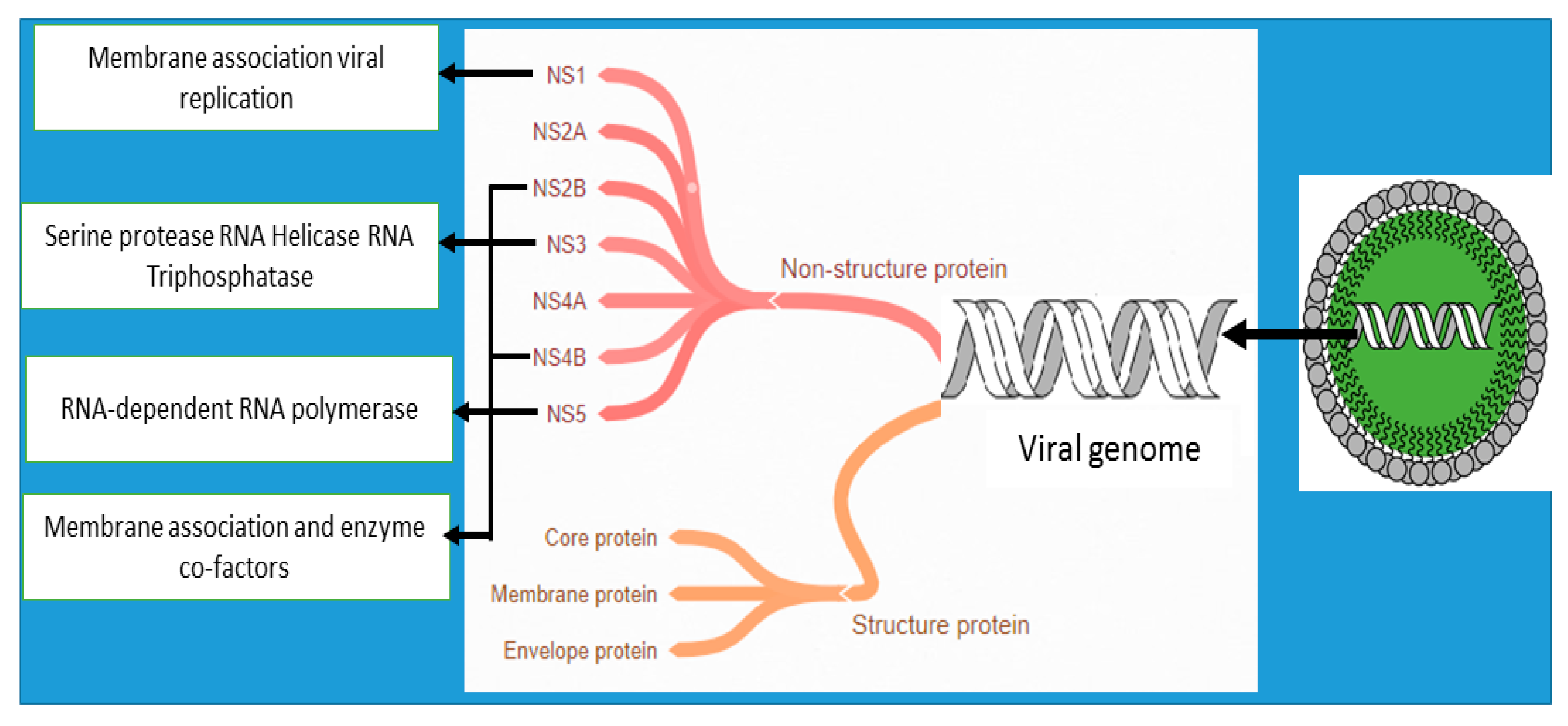
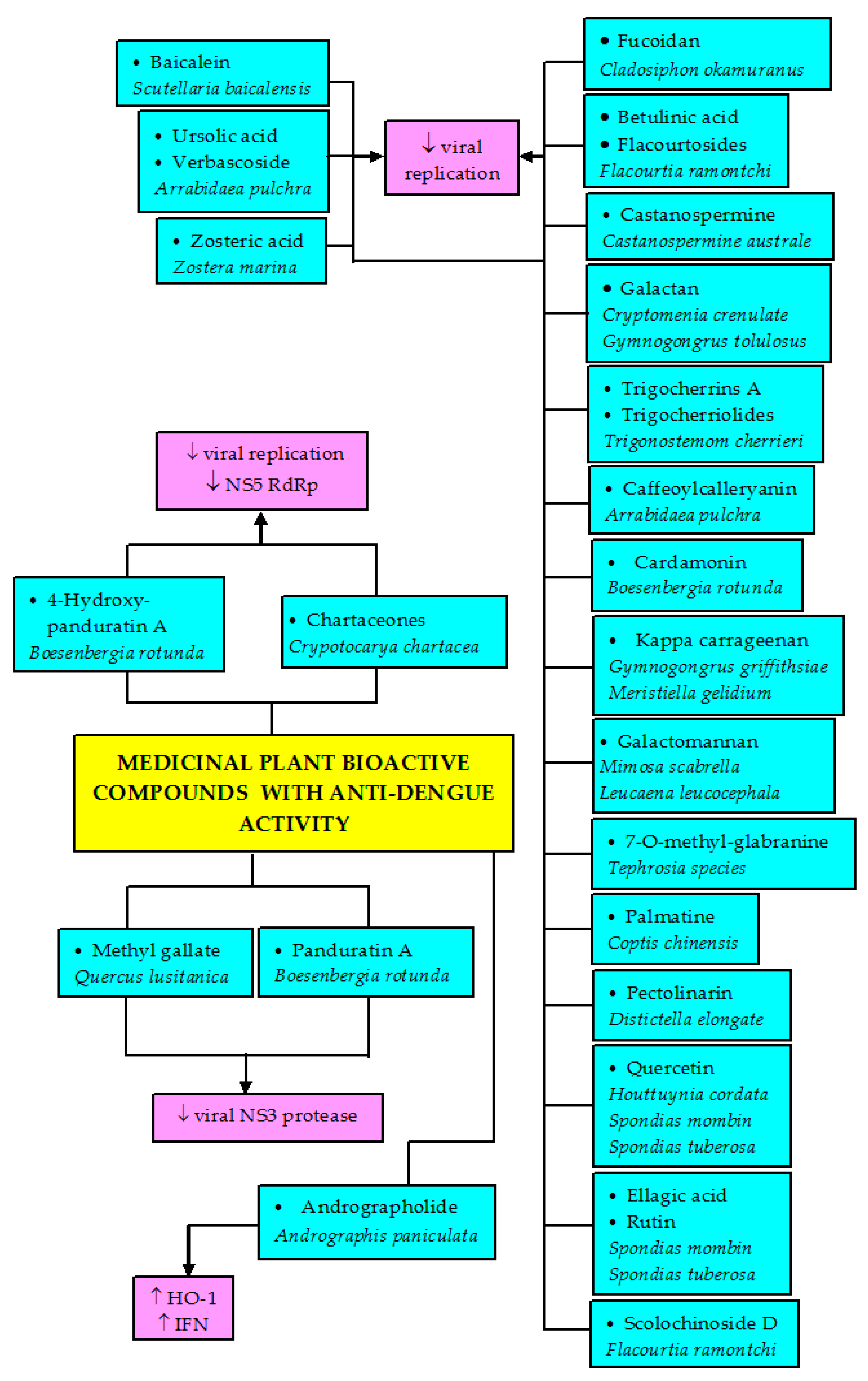
4.4. Anti-Dengue Bioactive Components from Medicinal Plants
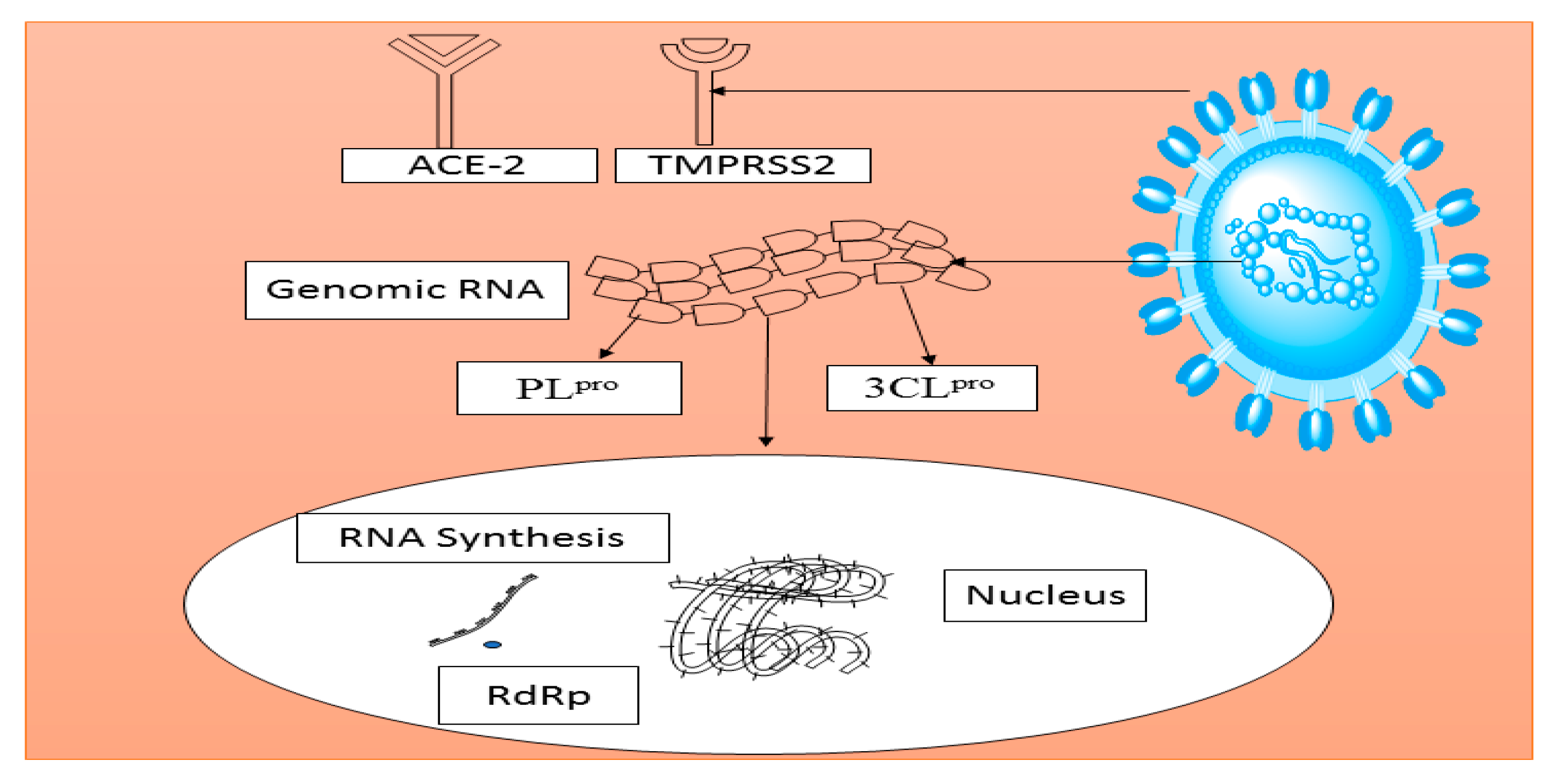
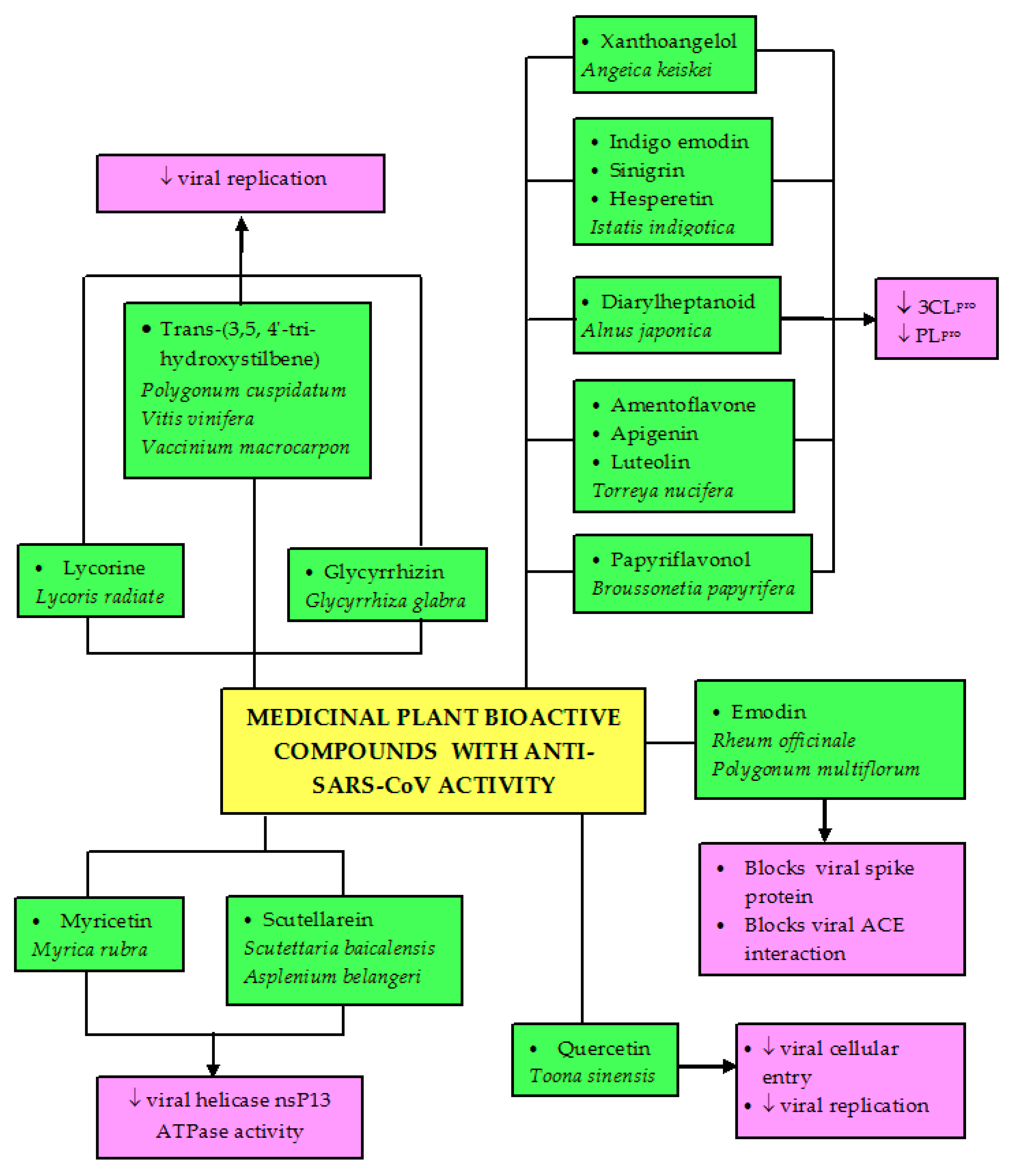
5. Potential Traditional Plants-Based Medicines for SARS-CoV-2 (COVID-19)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Source | Compounds | Mode of Action | Reference |
|---|---|---|---|
| Boesenbergia. rotunda | Panduratin | Inhibited SARS-CoV-2 in E6 cells | [38] |
| Angelica keiskei | Xanthoangelol | Inhibited PLpro in vitro | [152] |
| Alnus japonica | Hirsutenone | Inhibited PLpro in vitro | [166] |
| Hirustanonol | Inhibited PLpro in vitro | [166] | |
| Oregonin | Inhibited PLpro in vitro | [166] | |
| Rubranol | Inhibited PLpro in vitro | [166] | |
| Rubraanoside | Inhibited PLpro in vitro | [166] | |
| Rubranoside A | Inhibited PLpro in vitro | [166] | |
| Broussonetia papyrifera | Broussochalcone B | Inhibited PLpro better than 3CLpro | [153] |
| Broussochalcone A | Inhibits PLpro higher than 3CLpro | [153] | |
| 4-Hydroxyisolonchocarpin | Inhibited PLpro better than 3CLpro | [153] | |
| Kazinol | Inhibited PLpro better than 3CLpro | [153] | |
| Kazinol A | Inhibited PLpro better than 3CLpro | [153] | |
| Kazinol B | Inhibited PLpro better than 3CLpro | [153] | |
| Kazinol F | Inhibited PLpro better than 3CLpro | [153] | |
| Kazinol J | Inhibited PLpro better than 3CLpro | [153] | |
| 3′-(3-methylbut-2-enyl)-3′, 4,7-trihydroxyflavane | Inhibited PLpro better than 3CLpro | [146] | |
| Papyriflavonol | Demonstrated highest inhibitory activity against PLpro (IC50 = 3.7 µM) | [153] | |
| Cinnamomic verum | Gallocatechin gallate | Inhibited wild-type SARS-CoV | [154] |
| Euphoria nerifolia | 3ß-friedelanol | Exhibited potent antiviral activity against SARS-CoV higher than positive control | [149] |
| Istatis indigotica | Hesperetin | Inhibited 3CLpro effect via cell free and cell-based cleavage assays | [151] |
| Sinigrin | Inhibited 3CLpro effect via cell free and cell-based cleavage assays | [151] | |
| Indigo | Inhibited 3CLpro effect via cell free and cell-based cleavage assays | [151] | |
| Emodin | Inhibited 3CLpro effect via cell free and cell-based cleavage assays | [142,155] |
6. Conclusions and Future Prospects
Funding
Conflicts of Interest
References
- WHO Coronavirus Disease (COVID-19). Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019?gclid=CjwKCAjwmf_4BRABEiwAGhDfSfUBiTxQG8b9kYVrWHD9pinxFCOxMnyC7tjIW6bkaoJq9HtVzyyNShoCDbIQAvD_BwE (accessed on 28 July 2020).
- WHO The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 15 September 2020).
- WHO Ebola Virus Disease. Available online: https://www.who.int/news-room/fact-sheets/detail/ebola-virus-disease (accessed on 14 August 2020).
- Mansori, K.; Ayubi, E.; Shadmani, F.K.; Hanis, S.M.; Khazaei, S.; Sani, M.; Moradi, Y.; Khazaei, S.; Mohammadbeigi, A. Estimates of global HIV/AIDS mortality, prevalence and incidence rates, and their association with the Human Development Index. Biomed. Res. Ther. 2017, 4, 1399. [Google Scholar] [CrossRef] [Green Version]
- Gasparini, R.; Amicizia, D.; Lai, P.L.; Panatto, D. Clinical and socioeconomic impact of seasonal and pandemic influenza in adults and the elderly. Hum. Vaccines Immunother. 2012, 8, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nováková, L.; Pavlík, J.; Chrenková, L.; Martinec, O.; Červený, L. Current antiviral drugs and their analysis in biological materials–Part II: Antivirals against hepatitis and HIV viruses. J. Pharm. Biomed. Anal. 2018, 147, 378–399. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Jiang, X.; Kanda, T.; Nakamoto, S.; Saito, K.; Nakamura, M.; Wu, S.; Haga, Y.; Sasaki, R.; Sakamoto, N.; Shirasawa, H.; et al. The JAK2 inhibitor AZD1480 inhibits hepatitis A virus replication in Huh7 cells. Biochem. Biophys. Res. Commun. 2015, 458, 908–912. [Google Scholar] [CrossRef]
- Chung, T.H.; Kim, J.C.; Kim, M.K.; Choi, S.C.; Kim, S.L.; Chung, J.M.; Lee, I.S.; Kim, S.H.; Hahn, K.S.; Lee, I.P. Investigation of Korean plant extracts for potential phytotherapeutic agents against B-virus hepatitis. Phytother. Res. 1995, 9, 429–434. [Google Scholar] [CrossRef]
- Akram, M.; Tahir, I.M.; Shah, S.M.A.; Mahmood, Z.; Altaf, A.; Ahmad, K.; Munir, N.; Daniyal, M.; Nasir, S.; Mehboob, H. Antiviral potential of medicinal plants against HIV, HSV, influenza, hepatitis, and coxsackievirus: A systematic review. Phytother. Res. 2018, 32, 811–822. [Google Scholar] [CrossRef]
- Wang, J.; Peng, Y.; Xu, H.; Cui, Z.; Williams, R.O. The COVID-19 Vaccine Race: Challenges and Opportunities in Vaccine Formulation. AAPS PharmSciTech 2020, 21, 225. [Google Scholar] [CrossRef]
- Thuener, J. Hepatitis A and B infections. Prim. Care Clin. Off. Pract. 2017, 44, 621–629. [Google Scholar] [CrossRef]
- Ma, S.-J.; Li, X.; Xiong, Y.-Q. Combination measles-mumps-rubella-varicella vaccine in healthy children: A systematic review and meta-analysis of immunogenicity and safety. Medicine 2015, 94, e1721. [Google Scholar] [CrossRef]
- Singh, R.; Singh, K.P.; Cherian, S.; Saminathan, M.; Kapoor, S.; Reddy, G.B.M.; Panda, S.; Dhama, K. Rabies–Epidemiology, pathogenesis, public health concerns and advances in diagnosis and control: A comprehensive review. Vet. Q. 2017, 37, 212–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perera, S.D.; Jayawardena, U.A.; Jayasinghe, C.D. Potential use of Euphorbia hirta for dengue: A systematic review of scientific evidence. J. Trop. Med. 2018, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO WHO Epidemiology. Available online: https://www.who.int/denguecontrol/epidemiology/en/ (accessed on 8 August 2020).
- Raja, D.B.; Mallol, R.; Ting, C.Y.; Kamaludin, F.; Ahmad, R.; Ismail, S.; Jayaraj, V.J.; Sundram, B.M. Artificial intelligence model as predictor for dengue outbreaks. Malays. J. Public Health Med. 2019, 19, 103–108. [Google Scholar] [CrossRef]
- Grzybowski, A.; Tiboni, M.; da Silva, M.A.N.; Chitolina, R.F.; Passos, M.; Fontana, J.D. The combined action of phytolarvicides for the control of dengue fever vector, Aedes aegypti. Braz. J. Pharmacogn. 2012, 22, 549–557. [Google Scholar] [CrossRef] [Green Version]
- Stanaway, J.D.; Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Coffeng, L.E.; Brady, O.J.; Hay, S.I.; Bedi, N.; Bensenor, I.M.; Castañeda-Orjuela, C.A.; et al. The global burden of dengue: An analysis from the global burden of disease study 2013. Lancet Infect. Dis. 2016, 16, 712–723. [Google Scholar] [CrossRef] [Green Version]
- Simo, F.B.N.; Bigna, J.J.; Kenmoe, S.; Ndangang, M.S.; Temfack, E.; Moundipa, P.F.; Demanou, M. Dengue virus infection in people residing in Africa: A systematic review and meta-analysis of prevalence studies. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Goel, A.; Patel, D.N.; Lakhani, K.K.; Agarwal, S.B.; Agarwal, A.; Singla, S.; Agarwal, R. Dengue fever—a dangerous foe. J. Indian Acad. Clin. Med. 2004, 5, 247–258. [Google Scholar]
- WHO Dengue and Severe Dengue. Available online: www.who.int/mediacentre/factsheets/fs117/en/index.html/ (accessed on 23 June 2020).
- Gubler, D.J. Dengue/dengue haemorrhagic fever: History and current status. In Proceedings of the Novartis Foundation Symposium; Wiley Online Library: Chichester, UK, 2006; Volume 277, p. 3. [Google Scholar]
- Guha-Sapir, D.; Schimmer, B. Dengue fever: New paradigms for a changing epidemiology. Emerg. Themes Epidemiol. 2005, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Abd Kadir, S.L.; Yaakob, H.; Mohamed Zulkifli, R. Potential anti-dengue medicinal plants: A review. J. Nat. Med. 2013, 67, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Agnandji, S.T. Malaria vaccine: WHO position paper. N. Engl. J. Med. 2011, 356, 1863–1875. [Google Scholar]
- Fatima, K.; Syed, N.I. Dengvaxia controversy: Impact on vaccine hesitancy. J. Glob. Health 2018, 8, 8–11. [Google Scholar] [CrossRef] [PubMed]
- CDC Centers for Disease Control and Prevention. Available online: https://www.cdc.gov/ (accessed on 4 August 2020).
- Wang, C.; Horby, P.W.; Hayden, F.G.; Gao, G.F. A novel coronavirus outbreak of global health concern. Lancet 2020, 395, 470–473. [Google Scholar] [CrossRef] [Green Version]
- Singhal, T. A Review of Coronavirus Disease-2019 (COVID-19). Indian J. Pediatr. 2020, 87, 281–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan-yeung, M.; Xu, R. SARS: Epidemiology. Respirology 2003, 8, S9–S14. [Google Scholar] [CrossRef] [PubMed]
- WHO Coronavirus Disease (COVID-19) Dashboard | WHO Coronavirus Disease (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 4 August 2020).
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Wikipedia Template: Covid-19 Pandemic Data-Wikipedia. Available online: https://en.m.wikipedia.org/wiki/Template:Covid-19_pandemic_data (accessed on 17 August 2020).
- Ridwan, R. COVID-19 and dengue: A deadly duo. Trop. Dr. 2020, 50, 270–272. [Google Scholar] [CrossRef]
- Leardkamolkarn, V.; Sirigulpanit, W.; Phurimsak, C.; Kumkate, S.; Himakoun, L.; Sripanidkulchai, B. The inhibitory actions of Houttuynia cordata aqueous extract on dengue virus and dengue-infected cells. J. Food Biochem. 2012, 36, 86–92. [Google Scholar] [CrossRef]
- Kiat, T.S.; Pippen, R.; Yusof, R.; Ibrahim, H.; Khalid, N.; Rahman, N.A. Inhibitory activity of cyclohexenyl chalcone derivatives and flavonoids of fingerroot, Boesenbergia rotunda (L.), towards dengue-2 virus NS3 protease. Bioorg. Med. Chem. Lett. 2006, 16, 3337–3340. [Google Scholar] [CrossRef]
- Kanjanasirirat, P.; Suksatu, A.; Manopwisedjaroen, S.; Munyoo, B.; Tuchinda, P.; Jearawuttanakul, K.; Seemakhan, S.; Charoensutthivarakul, S.; Wongtrakoongate, P.; Rangkasenee, N.; et al. High-content screening of Thai medicinal plants reveals Boesenbergia rotunda extract and its component Panduratin A as Anti-SARS-CoV-2 agents. Res. Squre. 2020, 10, 19963. [Google Scholar] [CrossRef]
- Lau, K.M.; Lee, K.M.; Koon, C.M.; Cheung, C.S.F.; Lau, C.P.; Ho, H.M.; Lee, M.Y.H.; Au, S.W.N.; Cheng, C.H.K.; Lau, C.B.S.; et al. Immunomodulatory and anti-SARS activities of Houttuynia cordata. J. Ethnopharmacol. 2008, 118, 79–85. [Google Scholar] [CrossRef]
- Derosa, G.; Maffioli, P.; D’Angelo, A.; Di Pierro, F. A role for quercetin in coronavirus disease 2019 (COVID-19). Phytother. Res. 2020, 1–7. [Google Scholar] [CrossRef] [PubMed]
- WHO Traditional, Complementary and Integrative Medicine. Available online: https://www.who.int/health-topics/traditional-complementary-and-integrative-medicine#tab=tab_1 (accessed on 8 August 2020).
- Saleh, M.S.M.; Siddiqui, M.J.; Mat So’ad, S.Z.; Roheem, F.O.; Saidi-Besbes, S.; Khatib, A. Correlation of FT-IR fingerprint and α-glucosidase inhibitory activity of salak (Salacca zalacca) fruit extracts utilizing orthogonal partial least square. Molecules 2018, 23, 1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meneses, R.; Ocazionez, R.E.; Martínez, J.R.; Stashenko, E.E. Inhibitory effect of essential oils obtained from plants grown in Colombia on yellow fever virus replication in vitro. Ann. Clin. Microbiol. Antimicrob. 2009, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rates, S.M.K. Plants as source of new drugs. Toxicon 2001, 39, 603–613. [Google Scholar] [CrossRef]
- Ganjhu, R.K.; Mudgal, P.P.; Maity, H.; Dowarha, D.; Devadiga, S.; Nag, S.; Arunkumar, G. Herbal plants and plant preparations as remedial approach for viral diseases. Virus Dis. 2015, 26, 225–236. [Google Scholar] [CrossRef]
- Betancur-galvis, L.A.; Morales, G.E.; Forero, J.E.; Roldan, J. Cytotoxic and antiviral activities of Colombian medicinal plant extracts of the Euphorbia genus. Mem. Inst. Oswaldo Cruz. 2002, 97, 541–546. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.C.; Du, J.; But, P.P.H.; Deng, X.L.; Zhang, Y.W.; Ooi, V.E.C.; Xu, H.X.; Lee, S.H.S.; Lee, S.F. Antiviral Chinese medicinal herbs against respiratory syncytial virus. J. Ethnopharmacol. 2002, 79, 205–211. [Google Scholar] [CrossRef]
- Rothan, H.; Zulqarnain, M.; Ammar, Y..; Tan, E..; Rahman, N..; Yusof, R. Screening of antiviral activities in medicinal plants extracts against dengue virus using dengue NS2B-NS3 protease assay. Trop. Biomed. 2014, 31, 286–296. [Google Scholar]
- Lee, S.H.; Tang, Y.Q.; Rathkrishnan, A.; Wang, S.M.; Ong, K.C.; Manikam, R.; Payne, B.J.; Jaganath, I.B.; Sekaran, S.D. Effects of cocktail of four local Malaysian medicinal plants (Phyllanthus spp.) against dengue virus 2. BMC Complement. Altern. Med. 2013, 13, 192. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.I.C.; Ling, A.P.K.; Voon, K.G.L. Screening of anti-dengue activity in methanolic extracts of medicinal plants. BMC Complement. Altern. Med. 2014, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.H.; Wu, K.L.; Zhang, X.; Deng, S.Q.; Peng, B. In silico screening of Chinese herbal medicines with the potential to directly inhibit 2019 novel coronavirus. J. Integr. Med. 2020, 18, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Sarmah, S.; Lyndem, S.; Singha Roy, A. An investigation into the identification of potential inhibitors of SARS-CoV-2 main protease using molecular docking study. J. Biomol. Struct. Dyn. 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Macchiagodena, M.; Pagliai, M.; Procacci, P. Identification of potential binders of the main protease 3CLpro of the COVID-19 via structure-based ligand design and molecular modeling. Chem. Phys. Lett. 2020, 750, 137489. [Google Scholar] [CrossRef] [PubMed]
- Aanouz, I.; Belhassan, A.; El-Khatabi, K.; Lakhlifi, T.; El-ldrissi, M.; Bouachrine, M. Moroccan Medicinal plants as inhibitors against SARS-CoV-2 main protease: Computational investigations. J. Biomol. Struct. Dyn. 2020, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fiscal, R.R. Ethnomedicinal plants used by traditional healers in Laguna, Philippines. Asia Pac. J. Multidiscip. Res. 2017, 5, 132–137. [Google Scholar]
- Philippine Medicinal Plants Philippine Medicinal Plants. Available online: http://www.stuartxchange.org/Papaya.html (accessed on 14 August 2020).
- Chand, R.R.; Devi, S.S.; Kumari, S.S.; Kumar, S.S.; Goundar, N.N.; Naranyan, N.; Chandra, P. Traditional use of medicinal plants among selected villages in Fiji Islands: A review. Pac. Med. Stud. J. 2018, 1, 10–20. [Google Scholar]
- Sivasankari, B.; Pitchaimani, S.; Anandharaj, M. A study on traditional medicinal plants of Uthapuram, Madurai District, Tamilnadu, South India. Asian Pac. J. Trop. Biomed. 2013, 3, 975–979. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.; Sharif, R.; Mamun, A.H.; Rayhana, N.; Begum, K.; Tripty, F.; Keya, A.S.; Shetu, S.; Ema, S.P.; Al-Nahain, A. Phytomedicines of traditional health-care professionals in the vicinity of Lawachara Forest Reserve, Moulvibazar district, Bangladesh. Am. Eurasian J. Sustain. Agric. 2014, 8, 51–60. [Google Scholar]
- Gascon, M.G. Traditional ecological knowledge system of the matigsalug tribe in mitigating the effects of dengue and Malaria outbreak. Asian J. Health 2011, 1, 160–171. [Google Scholar] [CrossRef] [Green Version]
- De Guzman, G.Q.; Dacanay, A.T.L.; Andaya, B.A.; Alejandro, G.J.D. Ethnopharmacological studies on the uses of Euphorbia hirta in the treatment of dengue in selected indigenous communities in pangasinan (Philippines). J. Intercult. Ethnopharmacol. 2016, 5, 239–243. [Google Scholar] [CrossRef]
- Rahmatullah, M.; Mollik, M.A.H.; Azam, A.; Islam, M.R.; Chowdhury, M.A.M.; Jahan, R.; Chowdhury, M.H.; Rahman, T. Ethnobotanical survey of the santal tribe residing in Thakurgaon District, Bangladesh. Am. Eurasian J. Sustain. Agric. 2009, 3, 889–898. [Google Scholar]
- Hajdu, Z.; Hohmann, J. An ethnopharmacological survey of the traditional medicine utilized in the community of Porvenir, Bajo Paraguá Indian Reservation, Bolivia. J. Ethnopharmacol. 2012, 139, 838–857. [Google Scholar] [CrossRef] [PubMed]
- Smita, S. Indigenous medicinal plants used for treatment of dengue fever by Tribals of Chhattisgarh (India). Int. J. Pharm. Bio. Sci. 2015, 6, 404–410. [Google Scholar]
- Kandowangko, N.Y.; Latief, M.; Yusuf, R. Short communication: Inventory of traditional medicinal plants and their uses from Atinggola, North Gorontalo District, Gorontalo Province, Indonesia. Biodivers 2018, 19, 2294–2301. [Google Scholar] [CrossRef]
- Rahmatullah, M.; Jahan, R.; Safiul Azam, F.M.; Hossan, S.; Mollik, M.A.H.; Rahman, T. Folk medicinal uses of verbenaceae family plants in Bangladesh. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 53–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, Y.N. Traditional medicine in Fiji: Some herbal folk cures used by Fiji Indians. J. Ethnopharmacol. 1986, 15, 57–88. [Google Scholar] [CrossRef]
- Islam, M.K.; Saha, S.; Mahmud, I.; Mohamad, K.; Awang, K.; Jamal Uddin, S.; Rahman, M.M.; Shilpi, J.A. An ethnobotanical study of medicinal plants used by tribal and native people of Madhupur forest area, Bangladesh. J. Ethnopharmacol. 2014, 151, 921–930. [Google Scholar] [CrossRef]
- Qasim, M.; Abideen, Z.; Adnan, M.Y.; Ansari, R.; Gul, B.; Khan, M.A. Traditional ethno-botanical uses of medicinal plants from coastal areas of Pakistan. J. Coast. Life Med. 2014, 2, 22–30. [Google Scholar] [CrossRef]
- Gruya, G.A.; Roasario, R.D.; Palmes, N.D. Ethnomedicinal plants used by residents in Northern Surigao del Sur, Philippines. Nat. Prod. Chem. Res. 2014, 2, 2–6. [Google Scholar] [CrossRef] [Green Version]
- Miano, R.S.; Picardal, J.P.; Alonso, C.A.G.; Reuyan, D. Ethnobotanical Inventory and assessment of medically-important plant roots in Cebu island, Philippines. Asian J. Biodivers 2011, 2, 81–102. [Google Scholar] [CrossRef]
- Bradacs, G.; Heilmann, J.; Weckerle, C.S. Medicinal plant use in Vanuatu: A comparative ethnobotanical study of three islands. J. Ethnopharmacol. 2011, 137, 434–448. [Google Scholar] [CrossRef]
- Bussmann, R.W.; Sharon, D. Traditional medicinal plant use in Northern Peru: Tracking two thousand years of healing culture. J. Ethnobiol. Ethnomed. 2006, 2, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Deep, P.; Srivastava, V.; Verma, S. Current perspectives of medicinal plants having anti dengue potential. Int. J. Pharm. Sci. Rev. Res. 2018, 49, 91–96. [Google Scholar]
- Batoro, J.; Siswanto, D. Ethnomedicinal survey of plants used by local society in Poncokusumo district, Malang, East Java Province, Indonesia. Asian J. Med. Biol. Res. 2017, 3, 158–167. [Google Scholar] [CrossRef]
- Patil, S.; Shetty, S.; Bhide, R.; Narayanan, S. Evaluation of platelet augmentation activity of Carica papaya leaf aqueous extract in rats. J. Pharm. Phytochem. 2013, 1, 57. [Google Scholar]
- Arollado, E.C.; Pena, I.G.; Dahilig, V.R. Platelet augmentation activity of selected Philippine plants. Int. J. Pharm. Phytopharmacol. Res. 2014, 3, 121–123. [Google Scholar]
- Coloma, A.J.L.; Casilla, A.S.B.; Estolero, B.; Ulalan, M.G.E.; Veloso, G.D.; MAN, R. Thrombocytotic efficacy of Tawa-Tawa, papaya and Malunggay among aspirin-induced laboratory rabbits. ICHAMS Health Care J. 2015, 5, 8–16. [Google Scholar]
- George, A.; Zandi, K.; Biggins, J.; Chinnappan, S.; Hassandarvish, P.; Yusof, A. Antiviral activity of a standardized root water extract of Eurycoma longifolia (Physta®) against dengue virus. Trop. Biomed. 2019, 36, 412–421. [Google Scholar]
- Ichsyani, M.; Ridhanya, A.; Risanti, M.; Desti, H.; Ceria, R.; Putri, D.H.; Sudiro, T.M.; Dewi, B.E. Antiviral effects of Curcuma longa L. against dengue virus in vitro and in vivo. Conf. Ser. Earth Environ. Sci. 2017, 101, 012005. [Google Scholar] [CrossRef] [Green Version]
- Parida, M.M.; Upadhyay, C.; Pandya, G.; Jana, A.M. Inhibitory potential of neem (Azadirachta indica Juss) leaves on Dengue virus type-2 replication. J. Ethnopharmacol. 2002, 79, 273–278. [Google Scholar] [CrossRef]
- Norahmad, N.A.; Mohd Abd Razak, M.R.; Mohmad Misnan, N.; Md Jelas, N.H.; Sastu, U.R.; Muhammad, A.; Ho, T.C.D.; Jusoh, B.; Zolkifli, N.A.; Thayan, R.; et al. Effect of freeze-dried Carica papaya leaf juice on inflammatory cytokines production during dengue virus infection in AG129 mice. BMC Complement. Altern. Med. 2019, 19, 44. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumar, V.; Durairajan, S.S.K.; Alagarasu, K.; Li, M.; Dash, A.P. Recent updates on mouse models for human immunodeficiency, influenza, and dengue viral infections. Viruses 2019, 11, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandão, G.C.; Kroon, E.G.; Souza, D.E.R.; Filho, J.D.S.; Oliveira, A.B. Chemistry and antiviral activity of Arrabidaea pulchra (Bignoniaceae). Molecules 2013, 18, 9919–9932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, A.R.A.; Morais, S.M.; Marques, M.M.M.; Lima, D.M.; Santos, S.C.C.; Almeida, R.R.; Vieira, I.G.P.; Guedes, M.I.F. Antiviral activities of extracts and phenolic components of two Spondias species against dengue virus. J. Venom. Anim. Toxins Incl. Trop. Dis. 2011, 17, 406–413. [Google Scholar] [CrossRef]
- Rosmalena, R.; Elya, B.; Dewi, B.E.; Fithriyah, F.; Desti, H.; Angelina, M.; Hanafi, M.; Lotulung, P.D.; Prasasty, V.D.; Seto, D. The antiviral effect of indonesian medicinal plant extracts against dengue virus in vitro and in silico. Pathogens 2019, 8, 85. [Google Scholar] [CrossRef] [Green Version]
- Klawikkan, N.; Nukoolkarn, V.; Jirakanjanakit, N.; Yoksan, S.; Wiwat, C.; Thirapanmethee, K. Effect of Thai medicinal plant extracts against dengue virus in vitro. Mahidol. Univ. J. Pharm. Sci. 2011, 38, 13–18. [Google Scholar]
- Ramalingam, S.; Karupannan, S.; Padmanaban, P.; Vijayan, S.; Sheriff, K.; Palani, G.; Krishnasam, K.K. Anti-dengue activity of Andrographis paniculata extracts and quantification of dengue viral inhibition by SYBR green reverse transcription polymerase chain reaction Senthilraja. Ayu 2018, 39, 87–91. [Google Scholar] [CrossRef]
- Jiang, W.L.; Luo, X.L.; Kuang, S.J. Effects of Alternanthera philoxeroides Griseb against dengue virus in vitro. Di 1 jun yi da xue xue bao= Acad. J. First Med. Coll. PLA 2005, 25, 454–456. [Google Scholar]
- Tayone, W.C.; Tayone, J.C.; Hashimoto, M. Isolation and structure elucidation of potential Anti-Dengue metabolites from Tawa-Tawa (Euphorbia hirta Linn.). Walailak J. Sci. Technol. 2014, 11, 825–832. [Google Scholar] [CrossRef]
- Ahmad, N.; Fazal, H.; Ayaz, M.; Abbasi, B.H.; Mohammad, I.; Fazal, L. Dengue fever treatment with Carica papaya leaves extracts. Asian Pac. J. Trop. Biomed. 2011, 1, 330–333. [Google Scholar] [CrossRef] [Green Version]
- Hettige S Salutary effects of Carica papaya leaf extract in dengue fever patients–a pilot study. Sri Lankan Family Physician 2008, 29, 17–19.
- Subenthiran, S.; Choon, T.C.; Cheong, K.C.; Thayan, R.; Teck, M.B.; Muniandy, P.K.; Afzan, A.; Abdullah, N.R.; Ismail, Z. Carica papaya leaves juice significantly accelerates the rate of increase in platelet count among patients with dengue fever and dengue haemorrhagic fever. Evid.-Based Complement. Altern. Med. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadhwal, A.K.; Ankit, B.S.; Chahar, C.; Tantia, P.; Sirohi, P.; Agrawal, R.P. Effect of Carica papaya leaf extract capsule on platelet count in patients of dengue fever with thrombocytopenia. J. Assoc. Physicians India. 2016, 64, 22–26. [Google Scholar] [PubMed]
- Yunita, F.; Hanani, E.; Kristianto, J. The effect of Carica papaya L. leaves extract capsules on platelets count and hematocrit level in dengue fever patient. Int. J. Med. Arom. Plants 2012, 2, 2249–4340. [Google Scholar]
- Sathyapalan, D.T.; Padmanabhan, A.; Moni, M.; P-Prabhu, B.; Prasanna, P.; Balachandran, S.; Trikkur, S.P.; Jose, S.; Edathadathil, F.; Anilkumar, J.O.; et al. Efficacy & safety of Carica papaya leaf extract (CPLE) in severe thrombocytopenia (≤30,000/μl) in adult dengue–Results of a pilot study. PLoS ONE 2020, 15, e0228699. [Google Scholar] [CrossRef] [Green Version]
- Mir, M.; Khurshid, R.; Aftab, R. Management of thrombocytopenia and flu-like symptoms in dengue patients with herbal water of Euphorbia hirta. J. Ayub Med. Coll. Abbottabad. 2012, 24, 6–9. [Google Scholar]
- Ling, A.P.K.; Khoo, B.F.; Seah, C.H.; Foo, K.Y.; Cheah, R.K.; Chye, S.O.; Koh, R.Y. Inhibitory activities of methanol extracts of Andrographis paniculata and Ocimum sanctum against dengue-1virus. In Proceedings of the International Conference on Biological, Environment and Food Engineering (BEFE-2014), Bali, Indonesia, 4–5 August 2014. [Google Scholar] [CrossRef]
- Sathasivam, K.; Ramanathan, S.; Mansor, S.M.; Haris, M.R.M.H.; Wernsdorfer, W.H. Thrombocyte counts in mice after the administration of papaya leaf suspension. Wien. Klin. Wochenschr. 2009, 121, 19–22. [Google Scholar] [CrossRef]
- Sood, R.; Raut, R.; Tyagi, P.; Pareek, P.K.; Barman, T.K.; Singhal, S.; Shirumalla, R.K.; Kanoje, V.; Subbarayan, R.; Rajerethinam, R.; et al. Cissampelos pareira Linn: Natural source of potent antiviral activity against all four dengue virus serotypes. PLoS Negl. Trop. Dis. 2015, 9, e0004255. [Google Scholar] [CrossRef]
- Simões, L.R.; Maciel, G.M.; Brandão, G.C.; Kroon, E.G.; Castilho, R.O.; Oliveira, A.B. Antiviral activity of Distictella elongata (Vahl) Urb. (Bignoniaceae), a potentially useful source of anti-dengue drugs from the state of Minas Gerais, Brazil. Lett. Appl. Microbiol. 2011, 53, 602–607. [Google Scholar] [CrossRef]
- Saptawati, L.; Febrinasari, R.P.; Yudhani, R.D.; Yono, H.; Faza, A.G.; Luthfiani, S.; Ummiyati, H.S.; Sudiro, T.M.; Dewi, B.E. In vitro study of eight Indonesian plants extracts as anti Dengue virus. Health Sci. J. Indones. 2017, 8, 12–18. [Google Scholar] [CrossRef]
- Bourjot, M.; Leyssen, P.; Eydoux, C.; Guillemot, J.C.; Canard, B.; Rasoanaivo, P.; Guéritte, F.; Litaudon, M. Flacourtosides A-F, phenolic glycosides isolated from Flacourtia ramontchi. J. Nat. Prod. 2012, 75, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, A.C.; Valente, L.M.M.; Gomes, M.; Barboza, R.S.; Wolff, T.; Neris, R.L.S.; Figueiredo, C.M.; Assunção-Miranda, I. Antiviral activity of Faramea bahiensis leaves on dengue virus type-2 and characterization of a new antiviral flavanone glycoside. Phytochem. Lett. 2017, 19, 220–225. [Google Scholar] [CrossRef]
- Barboza, R.S.; Valente, L.M.M.; Wolff, T.; Assunção-Miranda, I.; Neris, R.L.S.; Guimarães-Andrade, I.P.; Gomes, M. Antiviral activity of Faramea hyacinthina and Faramea truncata leaves on dengue virus type-2 and their major compounds. Chem. Biodivers 2018, 15, e1700393. [Google Scholar] [CrossRef] [PubMed]
- Wolff, T.; Berrueta, L.A.; Valente, L.M.M.; Barboza, R.S.; Neris, R.L.S.; Guimarães-Andrade, I.P.; Assunção-Miranda, I.; Nascimento, A.C.; Gomes, M.; Gallo, B.; et al. Comprehensive characterisation of polyphenols in leaves and stems of three anti-dengue virus type-2 active Brazilian Faramea species (Rubiaceae) by HPLC-DAD-ESI-MS/MS. Phytochem. Anal. 2019, 30, 62–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, M.; Ganju, L.; Katiyal, A.; Padwad, Y.; Mishra, K.P.; Chanda, S.; Karan, D.; Yogendra, K.M.S.; Sawhney, R.C. Effect of Hippophae rhamnoides leaf extract against Dengue virus infection in human blood-derived macrophages. Phytomedicine 2008, 15, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Chiow, K.H.; Phoon, M.C.; Putti, T.; Tan, B.K.H.; Chow, V.T. Evaluation of antiviral activities of Houttuynia cordata Thunb. extract, quercetin, quercetrin and cinanserin on murine coronavirus and dengue virus infection. Asian Pac. J. Trop. Med. 2016, 9, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, M.L.; Phoon, M.C.; Dong, S.X.; Tan, B.K.H.; Chow, V.T. Houttuynia cordata extracts and constituents inhibit the infectivity of dengue virus type 2 in vitro. Int. J. Integr. Biol. 2013, 14, 78–85. [Google Scholar]
- Wilson, D.K.; Shyamala, G.; Paulpandi, M.; Narayanasamy, A.; Siram, K.; Karuppaiah, A.; Sankar, V. Development and characterization of phytoniosome nano vesicle loaded with aqueous leaf extracts of Justicia adhatoda and Psidium guajoava against dengue virus (DEN-2). J. Clust. Sci. 2020, 6. [Google Scholar] [CrossRef]
- Trujillo-Correa, A.I.; Quintero-Gil, D.C.; Diaz-Castillo, F.; Quiñones, W.; Robledo, S.M.; Martinez-Gutierrez, M. In vitro and in silico anti-dengue activity of compounds obtained from Psidium guajava through bioprospecting. BMC Complement. Altern. Med. 2019, 19, 298. [Google Scholar] [CrossRef]
- Muliawan, S.Y.; Kit, L.S.; Devi, S.; Hashim, O.; Yusof, R. Inhibitory potential of Quercus lusitanica extract on dengue virus type 2 replication. Southeast Asian J. Trop. Med. Public Health 2006, 37, 132–135. [Google Scholar]
- Reis, S.R.I.N.; Valente, L.M.M.; Sampaio, A.L.; Siani, A.C.; Gandini, M.; Azeredo, E.L.; D’Avila, L.A.; Mazzei, J.L.; Maria das Graças, M.H.; Kubelka, C.F. Immunomodulating and antiviral activities of Uncaria tomentosa on human monocytes infected with dengue Virus-2. Int. Immunopharmacol. 2008, 8, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Wahab, N.Z.A.; Ibrahim, N.; Kamarudin, M.K.A.; Lananan, F.; Juahir, H.; Ghazali, A.; Yusra, A.F.I. Cytotoxicity and antiviral activity of Annona muricata aqueous leaves extract against dengue virus type 2. J. Fundam. Appl. Sci. 2018, 10, 580–589. [Google Scholar]
- Zarina, N.; Wahab, A.; Nor, N.; Ibrahim, N. Efficacy of Catharanthus roseus extract against dengue virus type 2 tnfection in vitro. Indian J. Public Health Res. Dev. 2020, 11, 1577–1580. [Google Scholar]
- Abd Wahab, N.Z.; Badya, N.; Ibrahim, N.; Kamarudin, M.K.A.; Juahir, H.; Toriman, M.E. Antiviral activity of Cynometra cauliflora leaves methanolic extract towards dengue virus type 2. Int. J. Eng. Technol. (UAE) 2018, 7, 344–347. [Google Scholar] [CrossRef]
- Wahab, N.Z.A.; Ibrahim, N.; Kamarudin, M.K.A.; Lananan, F.; Juahir, H.; Ghazali, A. In vitro antiviral activity of Orthosiphon stamineus extract against dengue virus type 2. J. Fundam. Appl. Sci. 2018, 10, 541–551. [Google Scholar]
- Panraksa, P.; Ramphan, S.; Khongwichit, S.; Smith, D.R. Activity of andrographolide against dengue virus. Antivir. Res. 2017, 139, 69–78. [Google Scholar] [CrossRef]
- Edwin, E.S.; Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Thanigaivel, A.; Ponsankar, A.; Pradeepa, V.; Selin-Rani, S.; Kalaivani, K.; Hunter, W.B.; Abdel-Megeed, A.; et al. Anti-dengue efficacy of bioactive andrographolide from Andrographis paniculata (Lamiales: Acanthaceae) against the primary dengue vector Aedes aegypti (Diptera: Culicidae). Acta Trop. 2016, 163, 167–178. [Google Scholar] [CrossRef]
- Whitby, K.; Pierson, T.C.; Geiss, B.; Lane, K.; Engle, M.; Zhou, Y.; Doms, R.W.; Diamond, M.S. Castanospermine, a potent inhibitor of dengue virus infection in vitro and in vivo. J. Virol. 2005, 79, 8698–8706. [Google Scholar] [CrossRef] [Green Version]
- Hidari, K.I.P.J.; Takahashi, N.; Arihara, M.; Nagaoka, M.; Morita, K.; Suzuki, T. Structure and anti-dengue virus activity of sulfated polysaccharide from a marine alga. Biochem. Biophys. Res. Commun. 2008, 376, 91–95. [Google Scholar] [CrossRef]
- Talarico, L.B.; Pujol, C.A.; Zibetti, R.G.M.; Faría, P.C.S.; Noseda, M.D.; Duarte, M.E.R.; Damonte, E.B. The antiviral activity of sulfated polysaccharides against dengue virus is dependent on virus serotype and host cell. Antivir. Res. 2005, 66, 103–110. [Google Scholar] [CrossRef]
- Pujol, C.A.; Estevez, J.M.; Carlucci, M.J.; Ciancia, M.; Cerezo, A.S.; Damonte, E.B. Novel DL-galactan hybrids from the red seaweed Gymnogongrus torulosus are potent inhibitors of herpes simplex virus and dengue virus. Antivir. Chem. Chemother. 2002, 13, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talarico, L.B.; Duarte, M.E.R.; Zibetti, R.G.M.; Noseda, M.D.; Damonte, E.B. An algal-derived DL-galactan hybrid is an efficient preventing agent for in vitro dengue virus infection. Planta Med. 2007, 73, 1464–1468. [Google Scholar] [CrossRef] [PubMed]
- De Sf-Tischer, P.C.; Talarico, L.B.; Noseda, M.D.; Silvia, S.M.; Damonte, E.B.; Duarte, M.E.R. Chemical structure and antiviral activity of carrageenans from Meristiella gelidium against herpes simplex and dengue virus. Carbohydr. Polym. 2006, 63, 459–465. [Google Scholar] [CrossRef]
- Jasamai, M.; Boon, Y.W.; Sakulpanich, A.; Jaleel, A. Current prevention and potential treatment options for dengue infection. J. Pharm. Pharm. Sci. 2019, 22, 440–456. [Google Scholar] [CrossRef] [PubMed]
- Allard, P.M.; Dau, E.T.H.; Eydoux, C.; Guillemot, J.C.; Dumontet, V.; Poullain, C.; Canard, B.; Guéritte, F.; Litaudon, M. Alkylated flavanones from the bark of cryptocarya chartacea as dengue virus NS5 polymerase inhibitors. J. Nat. Prod. 2011, 74, 2446–2453. [Google Scholar] [CrossRef] [PubMed]
- Allard, P.M.; Leyssen, P.; Martin, M.T.; Bourjot, M.; Dumontet, V.; Eydoux, C.; Guillemot, J.C.; Canard, B.; Poullain, C.; Guéritte, F.; et al. Antiviral chlorinated daphnane diterpenoid orthoesters from the bark and wood of Trigonostemon cherrieri. Phytochemistry 2012, 84, 160–168. [Google Scholar] [CrossRef]
- Sharma, V.; Sharma, A.; Bharate, S.B. Natural products in mitigation of SARS CoV Infections. Curr. Med. Chem. 2020. [Google Scholar] [CrossRef]
- Tseng, C.K.; Lin, C.K.; Wu, Y.H.; Chen, Y.H.; Chen, W.C.; Young, K.C.; Lee, J.C. Human heme oxygenase 1 is a potential host cell factor against dengue virus replication. Sci. Rep. 2016, 6, 32176. [Google Scholar] [CrossRef]
- Ono, L.; Wollinger, W.; Rocco, I.M.; Coimbra, T.L.M.; Gorin, P.A.J.; Sierakowski, M.R. In vitro and in vivo antiviral properties of sulfated galactomannans against yellow fever virus (BeH111 strain) and dengue 1 virus (Hawaii strain). Antivir. Res. 2003, 60, 201–208. [Google Scholar] [CrossRef]
- Sánchez, I.; Gómez-Garibay, F.; Taboada, J.; Ruiz, B.H. Antiviral effect of flavonoids on the dengue virus. Phytother. Res. 2000, 14, 89–92. [Google Scholar] [CrossRef]
- Rahman, N.A.; Hadinur; Muliawan, S.; Rashid, N.N.; Muhamad, M.; Yusof, R. Studies on Quercus Iusitanica extracts on DENV-2 replication. Dengue Bull. 2006, 30, 260–269. [Google Scholar]
- Jia, F.; Zou, G.; Fan, J.; Yuan, Z. Identification of palmatine as an inhibitor of West Nile virus. Arch. Virol. 2010, 155, 1325–1329. [Google Scholar] [CrossRef] [PubMed]
- Zandi, K.; Teoh, B.T.; Sam, S.S.; Wong, P.F.; Mustafa, M.R.; AbuBakar, S. Novel antiviral activity of baicalein against dengue virus. BMC Complement. Altern. Med. 2012, 12, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, C.R.; Costin, J.M.; Fink, R.C.; McMichael, M.; Fontaine, K.A.; Isern, S.; Michael, S.F. In vitro inhibition of dengue virus entry by p-sulfoxy-cinnamic acid and structurally related combinatorial chemistries. Antivir. Res. 2008, 80, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, M.; Zevini, A.; Palermo, E.; Muscolini, M.; Alexandridi, M.; Etna, M.P.; Coccia, E.M.; Fernandez-Sesma, A.; Coyne, C.; Zhang, D.D. Dengue virus targets Nrf2 for NS2B3-mediated degradation leading to enhanced oxidative stress and viral replication. J. Virol. 2020, 94, 1–20. [Google Scholar] [CrossRef]
- Valadão, A.L.C.; Aguiar, R.S.; de Arruda, L.B. Interplay between inflammation and cellular stress triggered by Flaviviridae viruses. Front. Microbiol. 2016, 7, 1233. [Google Scholar] [CrossRef] [Green Version]
- Siti, H.N.; Jalil, J.; Asmadi, A.Y.; Kamisah, Y. Roles of rutin in cardiac remodeling. J. Funct. Foods. 2020, 64, 103606. [Google Scholar] [CrossRef]
- Gui, J.S.; Jalil, J.; Jubri, Z.; Kamisah, Y. Parkia speciosa empty pod extract exerts anti-inflammatory properties by modulating NFκB and MAPK pathways in cardiomyocytes exposed to tumor necrosis factor-α. Cytotechnology 2019, 71, 79–89. [Google Scholar] [CrossRef]
- Wu, T.; Peng, Y.; Yan, S.; Li, N.; Chen, Y.; Lan, T. Andrographolide ameliorates atherosclerosis by suppressing pro-inflammation and ROS generation-mediated foam cell formation. Inflammation 2018, 41, 1681–1689. [Google Scholar] [CrossRef]
- Zhang, A.; Wan, B.; Jiang, D.; Wu, Y.; Ji, P.; Du, Y.; Zhang, G. The Cytoprotective Enzyme Heme Oxygenase-1 Suppresses Pseudorabies Virus Replication in vitro. Front. Microbiol. 2020, 11, 412. [Google Scholar] [CrossRef]
- Kamisah, Y.; Lim, J.J.; Lim, C.L.; Asmadi, A.Y. Inhibitory effects of palm tocotrienol-rich fraction supplementation on bilirubin-metabolizing enzymes in hyperbilirubinemic adult rats. PLoS ONE 2014, 9, e0089248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gretebeck, L.M.; Subbarao, K. Animal models for SARS and MERS coronaviruses. Curr. Opin. Virol. 2015, 13, 123–129. [Google Scholar] [CrossRef]
- Wyganowska-Swiatkowska, M.; Nohawica, M.; Grocholewicz, K.; Nowak, G. Influence of herbal medicines on hmgb1 release, sars-cov-2 viral attachment, acute respiratory failure, and sepsis. A literature review. Int. J. Mol. Sci. 2020, 21, 1–26. [Google Scholar] [CrossRef]
- Chen, Z.; Nakamura, T. Statistical evidence for the usefulness of Chinese medicine in the treatment of SARS. Phytother. Res. 2004, 18, 592–594. [Google Scholar] [CrossRef] [PubMed]
- Shahrajabian, M.H.; Sun, W.; Shen, H.; Cheng, Q. Chinese herbal medicine for SARS and SARS-CoV-2 treatment and prevention, encouraging using herbal medicine for COVID-19 outbreak. Acta Agric. Scand. Sect. B Soil Plant Sci. 2020, 70, 437–443. [Google Scholar] [CrossRef]
- Chen, C.J.; Michaelis, M.; Hsu, H.K.; Tsai, C.C.; Yang, K.D.; Wu, Y.C.; Cinatl, J.; Doerr, H.W. Toona sinensis Roem tender leaf extract inhibits SARS coronavirus replication. J. Ethnopharmacol. 2008, 120, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Chang, F.R.; Yen, C.T.; Ei-Shazly, M.; Lin, W.H.; Yen, M.H.; Lin, K.H.; Wu, Y.C. Anti-human coronavirus (anti-HCoV) triterpenoids from the leaves of Euphorbia neriifolia. Nat. Prod. Commun. 2012, 7, 1415–1417. [Google Scholar] [CrossRef] [Green Version]
- Wen, C.C.; Shyur, L.F.; Jan, J.T.; Liang, P.H.; Kuo, C.J.; Arulselvan, P.; Wu, J.B.; Kuo, S.C.; Yang, N.S. Traditional chinese medicine herbal extracts of Cibotium barometz, Gentiana scabra, Dioscorea batatas, Cassia tora, and Taxillus chinensis inhibit sars-cov replication. J. Tradit. Complement. Med. 2011, 1, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.W.; Tsai, F.J.; Tsai, C.H.; Lai, C.C.; Wan, L.; Ho, T.Y.; Hsieh, C.C.; Chao, P.D.L. Anti-SARS coronavirus 3C-like protease effects of Isatis indigotica root and plant-derived phenolic compounds. Antivir. Res. 2005, 68, 36–42. [Google Scholar] [CrossRef]
- Park, J.Y.; Ko, J.A.; Kim, D.W.; Kim, Y.M.; Kwon, H.J.; Jeong, H.J.; Kim, C.Y.; Park, K.H.; Lee, W.S.; Ryu, Y.B. Chalcones isolated from Angelica keiskei inhibit cysteine proteases of SARS-CoV. J. Enzyme Inhib. Med. Chem. 2016, 31, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Park, J.Y.; Yuk, H.J.; Ryu, H.W.; Lim, S.H.; Kim, K.S.; Park, K.H.; Ryu, Y.B.; Lee, W.S. Evaluation of polyphenols from Broussonetia papyrifera as coronavirus protease inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 504–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, M.; Jiang, H.; Suzuki, Y.; Li, X.; Xiao, P.; Tanaka, T.; Ling, H.; Yang, B.; Saitoh, H.; Zhang, L.; et al. Procyanidins and butanol extract of Cinnamomi cortex inhibit SARS-CoV infection. Antivir. Res. 2009, 82, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, D.; Nandi, R.; Kumar, D. Evaluation of flavonoids as 2019-nCoV cell entry inhibitor through molecular docking and pharmacological analysis. ChemRxiv 2020, 6, 1–27. Available online: https://chemrxiv.org/articles/preprint/Evaluation_of_Flavonoids_as_2019-nCoV_Cell_Entry_Inhibitor_Through_Molecular_Docking_and_Pharmacological_Analysis/12071508 (accessed on 15 December 2020).
- Yi, L.; Li, Z.; Yuan, K.; Qu, X.; Chen, J.; Wang, G.; Zhang, H.; Luo, H.; Zhu, L.; Jiang, P.; et al. Small molecules blocking the entry of severe acute respiratory syndrome coronavirus into host cells. J. Virol. 2004, 78, 11334–11339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.S.; Lee, J.; Lee, J.M.; Kim, Y.; Chin, Y.W.; Jee, J.G.; Keum, Y.S.; Jeong, Y.J. Identification of myricetin and scutellarein as novel chemical inhibitors of the SARS coronavirus helicase, nsP13. Bioorganic Med. Chem. Lett. 2012, 22, 4049–4054. [Google Scholar] [CrossRef] [PubMed]
- Tanner, J.A.; Zheng, B.; Zhou, J.; Watt, R.M.; Jiang, J.; Wong, K.; Lin, Y.; Lu, L.; He, M.; Kung, H.; et al. The adamantane-derived bananins are potent inhibitors of the helicase activities and replication of SARS coronavirus. Chem. Biol. 2005, 12, 303–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.C.; Ho, C.T.; Chuo, W.H.; Li, S.; Wang, T.T.; Lin, C.C. Effective inhibition of MERS-CoV infection by resveratrol. BMC Infect. Dis. 2017, 17, 144. [Google Scholar] [CrossRef] [Green Version]
- Li, S.Y.; Chen, C.; Zhang, H.Q.; Guo, H.Y.; Wang, H.; Wang, L.; Zhang, X.; Hua, S.N.; Yu, J.; Xiao, P.G.; et al. Identification of natural compounds with antiviral activities against SARS-associated coronavirus. Antivir. Res. 2005, 67, 18–23. [Google Scholar] [CrossRef]
- Cinatl, J.; Morgenstern, B.; Bauer, G.; Chandra, P.; Rabenau, H.; Doerr, H.W. Glycyrrhizin, an active component of Liquorice roots, and replication of SARS-associated coronavirus. Lancet 2003, 361, 2045–2046. [Google Scholar] [CrossRef] [Green Version]
- Ho, T.Y.; Wu, S.L.; Chen, J.C.; Li, C.C.; Hsiang, C.Y. Emodin blocks the SARS coronavirus spike protein and angiotensin-converting enzyme 2 interaction. Antivir. Res. 2007, 74, 92–101. [Google Scholar] [CrossRef]
- Schwarz, S.; Wang, K.; Yu, W.; Sun, B.; Schwarz, W. Emodin inhibits current through SARS-associated coronavirus 3a protein. Antivir. Res. 2011, 90, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Niu, C.; Cherney, M.M.; Zhang, J.; Huitema, C.; Eltis, L.D.; Vederas, J.C.; James, M.N.G. A Mechanistic View of enzyme inhibition and peptide hydrolysis in the active site of the sars-cov 3c-like peptidase. J. Mol. Biol. 2007, 371, 1060–1074. [Google Scholar] [CrossRef] [PubMed]
- Ryu, Y.B.; Jeong, H.J.; Kim, J.H.; Kim, Y.M.; Park, J.Y.; Kim, D.; Naguyen, T.T.H.; Park, S.J.; Chang, J.S.; Park, K.H.; et al. Biflavonoids from Torreya nucifera displaying SARS-CoV 3CLpro inhibition. Bioorg. Med. Chem. 2010, 18, 7940–7947. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Jeong, H.J.; Kim, J.H.; Kim, Y.M.; Park, S.J.; Kim, D.; Park, K.H.; Lee, W.S.; Ryu, Y.B. Diarylheptanoids from Alnus japonica inhibit papain-Like protease of severe acute respiratory syndrome coronavirus. Biol. Pharm. Bull. 2012, b12-00623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyaerts, E.; Vijgen, L.; Pannecouque, C.; Van Damme, E.; Peumans, W.; Egberink, H.; Balzarini, J.; Van Ranst, M. Plant lectins are potent inhibitors of coronaviruses by interfering with two targets in the viral replication cycle. Antivir. Res. 2007, 75, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, L.E.; Lin, C.N.; Su, B.L.; Jan, T.R.; Chen, C.M.; Wang, C.H.; Lin, D.S.; Lin, C.T.; Chueh, L.L. Synergistic antiviral effect of Galanthus nivalis agglutinin and nelfinavir against feline coronavirus. Antivir. Res. 2010, 88, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Heurich, A.; Hofmann-Winkler, H.; Gierer, S.; Liepold, T.; Jahn, O.; Pohlmann, S. TMPRSS2 and ADAM17 Cleave ACE2 Differentially and Only Proteolysis by TMPRSS2 Augments Entry Driven by the Severe Acute Respiratory Syndrome Coronavirus Spike Protein. J. Virol. 2014, 88, 1293–1307. [Google Scholar] [CrossRef] [Green Version]
- Beltrán-García, J.; Osca-Verdegal, R.; Pallardó, F.V.; Ferreres, J.; Rodríguez, M.; Mulet, S.; Sanchis-Gomar, F.; Carbonell, N.; García-Giménez, J.L. Oxidative Stress and Inflammation in COVID-19-Associated Sepsis: The Potential Role of Anti-Oxidant Therapy in Avoiding Disease Progression. Antioxidants 2020, 9, 936. [Google Scholar] [CrossRef]
- Imai, M.; Iwatsuki-Horimoto, K.; Hatta, M.; Loeber, S.; Halfmann, P.J.; Nakajima, N.; Watanabe, T.; Ujie, M.; Takahashi, K.; Ito, M.; et al. Syrian hamsters as a small animal model for SARS-CoV-2 infection and countermeasure development. Proc. Natl. Acad. Sci. USA 2020, 117, 16587–16595. [Google Scholar] [CrossRef]






| Name | Family | Part Used | Preparation | Reference |
|---|---|---|---|---|
| Annona reticulata | Annonaceae | Barks and bulbs | Decoction | [60] |
| Andrapogon citratum | Poaceae | Oil | Oil put on candles | [66] |
| Azadirachta indica | Meliaceae | Leaves | Extracts | [64] |
| Brassica campestris | Brassicaceae | Oil | Mixture with camphor | [67] |
| Carica papaya | Caricaceae | Leaves | Juice | [56] |
| Fruits | Oral decoction of unripe fruit | [59] | ||
| Leaves and fruits | Not mentioned | [58] | ||
| Female leaves | Decoction, (boiling the leaf juice) | [57] | ||
| Leaves | Pounding and extraction | [55] | ||
| Catharanthus roseus | Apocynaceae | Whole plant | Oral decoction | [68] |
| Citrus limon | Rutaceae | Pulps | Juice mixed with Protium spruceanum | [63] |
| Clerodendrum viscosum | Lamiaceae | Leaves | Not mentioned | [66] |
| Coleus scutellarioides | Lamiaceae | Leaves | Juice | [60] |
| Enicostemma hyssopifolium | Gentianaceae | Not stated | Decoction | [69] |
| Euodia species | Rutaceae | Leaves | Water extract | [65] |
| Euphorbia hirta | Euphorbiaceae | Leaves | Decoction | [56,60] |
| Leaves and stems | Decoction | [70] | ||
| Roots | Oral decoction | [71] | ||
| Leaves | Decoction and infusion | [61] | ||
| Leaves, stems and roots | Decoction | [55] | ||
| Mentha arvensis | Lamiaceae | Leaves | Decoction | [60] |
| Mikania micrantha | Asteraceae | Leaves | Cold maceration | [72] |
| Mikania cordata | Asteraceae | Leaves | Leaves juice | [62] |
| Musa paradisica | Musaceae | Latex of the stems | Decoction | |
| Ocinum sanctum | Lamiaceae | Leaves | Juice | [57] |
| Leaves | Juice | [67] | ||
| Physalis angulata | Solanaceae | Leaves | Oral infusion | [72] |
| Phytolacca bogotensis | Phytolaccaceae | Fresh leaves | Infusion of fresh leaves | [73] |
| Protium spruceanum | Burseraceae | Stem barks | Decoction with Citrus limon | [63] |
| Psidium guajava | Myrtaceae | Leaves | Decoction | [74] |
| Leaves and young fruits | Not mentioned | [75] | ||
| Tinospora cordifolia | Menispermaceae | Leaves and stems | Decoction | [74] |
| Trigonella foenam | Fabaceae | Leaves | Water infusion | [74] |
| Synespalum dulcificum | Sapotaceae | Leaves | Juice | [60] |
| Vitex negundo | Lamiaceae | Leaves | Decoction | [60] |
| Zingiber purpureum | Zingiberaceae | Rhizomes and leaves | Oral decoction mixed with turmeric and onion | [65] |
| Plant | Family | Part Used | Type of Study | Mode of Action | Reference |
|---|---|---|---|---|---|
| Alternanthera photoperiods | Amaranthaceae | Whole plant | In vitro | Inhibited DENV-2 replication (IC50 = 47.43 µg/mL) | [89] |
| Alternanthera sessillis | Amaranthaceae | Leaves | In vivo | Significant platelet increasing activity | [77] |
| Acorus calamus | Acoracea | Leaves | In vitro | Inhibited DENV-2 replication (96.5% at a dose of 20 µg/mL) | [86] |
| Andrographis paniculata | Flaviviridae | Leaves | In vitro | Anti-DENV-1 activity in HEPG2 (78.3 ± 2.9 PFU/mL) | [88] |
| Ocimum sanctum | Lamiaceae | Leaves | In vitro | Anti-DENV-1 activity in HEPG2 (1020.0 ± 271.0 PFU/mL) | [98] |
| Arrabidaea pulchra | Bignoniaceae | Leaves | In vitro | Inhibited DENV-2 (EC50 = 46.8 ± 1.6 µg mL−1) | [84] |
| Azadirachta indica | Meliaceae. | Leaf extract | In vitro and in vivo | Inhibited DENV-2 replication in both in vitro and in vivo | [81] |
| Carica papaya | Caricaceae | Leaves | In vivo | Increased the platelet counts | [99] |
| Leaf extract | Human | Increased the platelet counts and the total white cell counts had increased | [92] | ||
| Leaf extract | Human | Increased the platelet counts | [91] | ||
| Leaf extract | In vivo | Increased the platelet counts | [77] | ||
| Leaf juice | Human | Increased the platelet counts | [93] | ||
| Leaves | In vivo | Increased the platelet count and decreased the clotting time in rats | [76] | ||
| Leaf juice | Human | Increased the platelet counts | [95] | ||
| Leaf juice | In vivo | Increased the platelet counts | [78] | ||
| Leaf juice | Human | Increased the platelet counts | [94] | ||
| Leaves | In vivo | Increased the plasma CCL2/MCP-1 level | [82] | ||
| Leaves | Human | Increased the platelet counts | [96] | ||
| Cissampelos pareira | Menispermacea | Aerial part | Human | Increased the platelet counts | [100] |
| Cladogynos orientalis | Euphorbiaceae | Whole plant | In vitro | Inhibited DENV-1–4 replication | [87] |
| Curcuma longa | Zingiberacea | Not stated | In vitro | Anti-DENV activity in Huh7it-1 cells (IC50 17.91 = μg/mL) | [80] |
| in vivo | Anti- DENV-2 and reduce viremia period | ||||
| Cymbopogon citratus | Poaceae | Root | In vitro | Inhibited DENV-2 replication (98.9% at a dose of 20 µg/mL) | [86] |
| Distictella elongata | Bignoniaceae | Leaves, stems and fruits | In vitro | Anti-DENV-2 activity (EC50 = 9.8 µg/mL) | [101] |
| Euphorbia hirta | Euphorbiaceae | Leaves | In vivo | DENV-2 inhibition by 34.7% | [15] |
| Whole plant | In vivo | Increased the platelet counts | [77] | ||
| Whole plant | In vitro | Significantly reduced the plaque forming capacity of the DENV-1-2 (85% and 90% respectively) | [90] | ||
| Leaves | In vitro | Inhibited DENV-2 replication (34.7% at 20 µg/mL dose) | [102] | ||
| Whole weed | Human | Increased the platelet counts | [97] | ||
| Eurycoma longifolia | Simaroubaceae | Root | In vitro | Inhibited DENV-1, DENV-2, DENV-3 and DENV-4 (IC50 = 33.84, 33.55, 58.35 and 119 µg/mL, respectively) | [79] |
| In vivo | 30% lower viral load and 12% higher platelet count compared to the control group | ||||
| Flacourtia ramontchi | Salicaceae | Stem barks | In vitro | Inhibited DENV-2 NS5 polymerase activity (89% inhibition at 10 μg/mL) | [103] |
| Flagellaria indica | Flagellariaceae | Whole plant | In vitro | Inhibited DENV-2 (45.52% at a dose of 12.5 µg/mL) | [87] |
| Faramea bahinensis | Rubiaceae | Leaves | In vitro | Anti-DENV activity in HEPG2 (100% reduction in viral load) | [104] |
| Faramea hyacinthina and Faramea truncata | Rubiaceae | Leaves | In vitro | Anti-DENV activity in HEPG2 (90 to 100% at a dose of 50 μg/mL) | [105] |
| Faramea bahinensis, Faramea hyacinthina and Faramea truncata | Rubiaceae | Leaves and stem | In vitro | Anti-DENV activity in HEPG2 (70 to 93% at a dose of 50 μg/mL) | [106] |
| Hippophae rhamnoides | Elaeagnaceae | Leaves | Human | Anti-DENV activity in BHK-21 cells (1 PFU/mL at a dose of 50 mg/mL), decreased TNF-α and increased IFN-Υ | [107] |
| Houttuynia cordata | Saururaceae | Whole plant | In vitro | Inhibited DENV-2 replication (35.99% at a dose of 1.56 µg/mL) | [87] |
| Aerial stem and leaves | In vitro | Inhibited the intracellular viral RNA replication (EC50 = 0.8 µg/mL) | [36] | ||
| Aerial leaves | In vitro | Inhibited DENV-2 replication (IC50 = 0.98 mg/µL) | [108,109] | ||
| Ipomea batata | Convolvulaceae | Leaves | In vivo | Significant platelet increasing activity | [77] |
| Justicia adhatoda | Acanthaceae | In vitro | Inhibited the growth of Vero cells infected with DENV-2 at a dose of 60 µg/m | [110] | |
| Piper retrofractum | Piperaceae | Whole plant | In vitro | Inhibited DENV-2 replication (84.93%at a dose of 100 µg/mL) | [87] |
| Psidium guajava | Myrtaceae | Bark | In vitro | Inhibited DENV-2 replication | [111] |
| Leaves | In vitro | Inhibited DENV-2 replication at a dose of 60 µg/µL | [110] | ||
| Phyllanthus sp | Phyllanthaceae | Whole plant | In vitro | Inhibited DENV-2 replication (91.48% at a dose of 250 µg/mL) | [49] |
| Myristica fragrans | Myristicaceae | Leaves | In vitro | Inhibited DENV-2 replication (122.7% at a dose of 20 µg/mL) | [86] |
| Quersus lucitanica | Fagaceae | Seed | In vitro | Inhibited DENV-2 (100% at a dose of 0.032 mg/mL) | [112] |
| Rhizophora apiculata | Rhizophoraceae | Whole plant | In vitro | Inhibited DENV-2 replication (56.14% at a dose of 12.5 µg/mL) | [87] |
| Spondias mombin & | Anacardiaceae | Leaves juice | In vitro | Inhibited DENV-2 replication (3.31% at a dose of 500 µg/µL) | [85] |
| Spondias tuberosa | Anacardiaceae | Leaves juice | In vitro | Inhibited DENV-2 replication (99% at a dose of 500 17.98 µg/mL) | [85] |
| Uncaria tomentosa | Rubiaceae | Stem barks of | In vitro | Reducing DENV-Ag+ cell rates | [113] |
| Annona muricata | Annonaceae | Fruit | In vitro | Inhibited DENV-2 replication (99% at a dose of 1.25 mg/mL) | [114] |
| Catharanthus roseus | Apocynaceae | Leaves | In vitro | Inhibited DENV-2 replication (60% at a dose of 0.078 mg/mL) | [115] |
| Cynometra cauliflora | Fabaceae | Leaves | In vitro | Inhibited DENV-2 replication (78% at a dose of 12.5 mg/mL) | [116] |
| Orthosiphon stamineus | Lamiaceae | Leaves | In vitro | Inhibited DENV-2 replication (88% at a dose of 0.31 mg/mL) | [117] |
| Compound | Class of Compound | Plant Source | Antiviral Effects | Reference |
|---|---|---|---|---|
| Andrographolide | Diterpenoid | Andrographis paniculata | Anti-DENV activity in HepG2 (EC50 = 21.304 µM) and HeLa cell lines (EC50 = 22.739 µM) | [118] |
| Good anti-DENV effects in both visual (EC50 = 0.56 µg/mL) and neutral red cytopathic effects (EC50 = 0.58 µg/mL) | [129] | |||
| Reduced mean number of A. aegypti eggs and increased larvae mortality concentration-dependently | [119] | |||
| Delayed disease onset, reduced mortality and DENV-1, DENV-2, DENV-3 and DENV-4 loads in infected mouse brains | [130] | |||
| Apiofuranoside | Flavanone glycosides | Faramea bahiensis | Controlled viral replication and reduced numbers of infected cells (12%) and RNA copies of DENV-2 (67%) in HepG2 cells | [104] |
| Betulinic acid 3ß-caffeate | Phenyl terpenoid | Flacourtia ramontchi | Inhibited DENV replication (IC50 = 0.85 µM) | [103] |
| Caffeoylcalleryanin | Phenolic glycoside | Arrabidaea pulchra | Anti-DENV-2 activity (IC50 = 2.8 µg/mL) | [84] |
| Cardamonin | Phenolic | Boesenbergia rotunda | Non-competitive anti-DENV activity | [37] |
| Castanospermine | Alkaloid | Castanospermine australe | Inhibited secretion and infectivity in all DENV serotypes in vitro. Only inhibited DENV-2 in vivo and prevented mouse mortality. | [120] |
| Chartaceones 1 | Dialkylated flavanone | Crypotocarya chartacea | Inhibited NS5 RdRp (IC50 = 14.8 µM) | [127] |
| Chartaceones 2 | Inhibited NS5 RdRp (IC50 = 72.7 µM) | [127] | ||
| Chartaceones 3 | Inhibited NS5 RdRp (IC50 = 4.2 µM) | [127] | ||
| Chartaceones 4 | Inhibited NS5 RdRp (IC50 = 1.8 µM) | [127] | ||
| Chartaceones 5 | Inhibited NS5 RdRp (IC50 = 2.9 µM) | [127] | ||
| Chartaceones 6 | Inhibited NS5 RdRp (IC50 = 2.4 µM) | [127] | ||
| Chlorogenic acid | Phenolic acid | Houttuynia cordata | Synergistic antiviral effect of combined hyperoside and chlorogenic acid | [109] |
| Ellagic acid | Phenolic acid | Spondias mombin and Spondias tuberosa | Anti-DENV-2 (25%) at 500 µg/mL | [85] |
| Flacourtoside A | Phenolic glycoside | Flacourtia ramontchi | Inhibited DENV replication (IC50 = 9.3 µM) | [103] |
| Flacourtoside B | Inhibited DENV replication (IC50 = 71.1 µM) | |||
| Flacourtoside C | Inhibited DENV replication (IC50 = 23.8 µM) | |||
| Flacourtoside D | Inhibited DENV replication (IC50 = 35.5 µM) | |||
| Flacourtoside E | Inhibited DENV replication (IC50 = 13.4 µM) | |||
| Flacourtoside F | Inhibited DENV replication (IC50 = 39.8 µM) | |||
| Fucoidan | Sulfated polysaccharide | Cladosiphon okamuranus | Inhibited DENV-2 Reduced infectivity by 20% at 10 µg/mL in BHK-21 cells | [121] |
| Galactan | Sulfated polysaccharide | Cryptomenia crenulate | Inhibited DENV-2 and DENV-3 replication in Vero cells (IC50 = 1.0 µg/mL), slight inhibitory effect against DENV-4, inactive against DENV-1 | [122] |
| Gymnogongrus tolulosus | Inhibited DENV-2 and DENV-3 replication in Vero cells (IC50 = 0.19–1.7 µg/mL) | [123] | ||
| Galactomannan | Polysaccharride | Mimosa scabrella | Reduced DENV-1 titer 100-fold in C6/36 cells at 347 mg/L | [131] |
| Leucaena leucocephala | Reduced DENV-1 titer 100-fold in C6/36 cells at 37 mg/L | [131] | ||
| Glabranine | Flavanone | Tephrosia species | Anti-DENV (70% inhibition) at 25 µM | [132] |
| Hyperoside | Flavonol glycoside | Houttuynia cordata | Synergistic antiviral effect of combination of hyperoside and chlorogenic acid | [109] |
| 4-Hydroxy-panduratin A | Phenolic derivative | Boesenbergia rotunda | Competitive inhibitory activity against DENV-2 NS3 protease (K1 = 21 µM) | [37] |
| Kappa carrageenan | Sulfated polysaccharide | Gymnogongrus griffithsiae | Selectivity inhibited DENV-2 (IC50 = 0.9 µg/mL), DENV-3 and DENV-4 replication. Inactive against DENV-1 | [122] |
| Meristiella gelidium | Anti-DENV-2 activity (IC50 between 0.14–1.6 µg/mL) | [125] | ||
| 7-O-methyl-glabranine | Flavanone | Tephrosia species | Exhibited 70% inhibition on DENV in a plaque-forming assay at 25 µM | [132] |
| Methyl gallate | Phenolic acid | Quercus lusitanica | Inhibited DENV-2 protease (98%) at 0.3 mg/mL | [133] |
| Palmatine | Alkaloid | Coptis chinensis | Inhibited DENV replication (EC50 = 26.4 µmol/L) | [134] |
| Panduratin A | Phenolic derivative | Boesenbergia rotunda | Competitive inhibitory activity against DENV-2 NS3 protease (K1 = 25 µM) | [37] |
| Pectolinarin | Flavone | Distictella elongate | Good anti-DENV-2 effect (EC50 = 86.4 µg/mL) | [101] |
| Pinostrobin | Flavanone | Boesenbergia rotunda | Non-competitive DENV replication inhibitory activity | [37] |
| Quercetin | Flavonoid | Houttuynia cordata | Anti-DENV-2 effect at 500 µg/mL (IC50 = 176.76 µg/mL) | [108] |
| Quercetin | Flavonol | Houttuynia cordata | Synergistic antiviral effect of quercetirin and quercetin combination | [109] |
| Spondias mombin and Spondias tuberosa | Inhibited viral replication (50%) of DENV-2 at 500 µg/mL | [85] | ||
| Quercetirin | Flavonoid glycoside | Houttuynia cordata | Exhibited less activity in an uncombined form compared to its mixture with quercetin (IC50 = 467.27 µg/mL) | [108] |
| Houttuynia cordata | Synergistic antiviral effect of quercetirin and quercetin combination. greater antiviral effects | [109] | ||
| Rutin | Flavonol glycoside | Spondias mombin and Spondias tuberosa | Inhibited viral replication of DENV-2 at 500 µg/mL by 68.42% | [85] |
| Scolochinenoside D | Phenolic glycoside | Flacourtia ramontchi | Inhibited DENV replication (IC50 = 9.5 µM) | [103] |
| Trigocherrin A | Diterpenoid | Trigonostemon cherrieri | Inhibited DENV replication (IC50 = 12.7 µM) | [128] |
| Trigocherriolide B | Diterpenoid | Trigonostemon cherrieri | Inhibited DENV replication (IC50 = 3.1 µM) | [128] |
| Trigocherriolide C | Diterpenoid | Trigonostemon cherrieri | Inhibited DENV replication (IC50 = 16.0 µM) | [128] |
| Ursolic acid | Triterpenoid | Arrabidaea pulchra | Displayed anti-DENV-2 (IC50 = 3.2 µg/mL) | [84] |
| Verbascoside | Phenyl glycoside | Arrabidaea pulchra | Displayed anti-DENV-2 (IC50 = 3.4 µg/mL) | [84] |
| Baicalein | Bioflavonoid | Scutellaria baicalensis | Displayed anti-DENV-2 (IC50 = 1.55 μg/mL) | [135] |
| Zosteric acid | Phenolic acid | Zostera marina | Displayed anti-DENV-2 (IC50 = 2.3 mM) | [136] |
| Species | Family | Part Used | Type of Study | Mode of Action | Reference |
|---|---|---|---|---|---|
| Boesenbergia rotunda | Zingiberaceae | Rhizome ethanol extract | In vitro | Suppressed SARS-CoV-2 infectivity in Vero E6 cells | [38] |
| Broussonetia papyrifera | Moraceae | Roots | In vitro | Chloroform fraction inhibited viral replication | [153] |
| Euphoria nerifolia | Euphorbiaceae | Leaves | In vitro | Exhibited antiviral activity against SARS-CoV | [149] |
| Toona sinensis | Meliaceae | Tender leaf extract | In vitro | Inhibited SARS-CoV replication | [148] |
| Gentiana scabra | Gentianaceae | Rhizomes | Inhibited viral replication of SARS-CoV in Vero E6 cells | [150] | |
| Dioscorea batatas | Dioscoreaceae | Tubers | In vitro | Inhibited viral replication of SARS-CoV in Vero E6 cells | [150] |
| Cassia tora | Caesalpinioideae | Dried seeds | In vitro | Inhibited viral replication of SARS-CoV in Vero E6 cells | [150] |
| Taxillus chinensis | Loranthaceae | Dried stems and leaves | In vitro | Inhibited viral replication of SARS-CoV in Vero E6 cells | [150] |
| Cibotium barometz | Cibotiaceae | Rhizomes | In vitro | Inhibited viral replication of SARS-CoV in Vero E6 cells | [150] |
| Cinnamomi verum | Lauraceae | Butanol fraction | In vitro | Inhibited wild type SARS-CoV | [154] |
| Houttuynia cordata | Saururaceae | Dried whole plant | Had antiviral activity against SARS-CoV via inhibition on 3CLpro and RNA-dependent RNA polymerase of the virus | [39] | |
| IsatisIndigotica | Cruciferae | Root water extract | In vitro | Inhibited the cleavage activity of SARS 3CLpro enzyme | [151] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleh, M.S.M.; Kamisah, Y. Potential Medicinal Plants for the Treatment of Dengue Fever and Severe Acute Respiratory Syndrome-Coronavirus. Biomolecules 2021, 11, 42. https://doi.org/10.3390/biom11010042
Saleh MSM, Kamisah Y. Potential Medicinal Plants for the Treatment of Dengue Fever and Severe Acute Respiratory Syndrome-Coronavirus. Biomolecules. 2021; 11(1):42. https://doi.org/10.3390/biom11010042
Chicago/Turabian StyleSaleh, Mohammed S. M., and Yusof Kamisah. 2021. "Potential Medicinal Plants for the Treatment of Dengue Fever and Severe Acute Respiratory Syndrome-Coronavirus" Biomolecules 11, no. 1: 42. https://doi.org/10.3390/biom11010042
APA StyleSaleh, M. S. M., & Kamisah, Y. (2021). Potential Medicinal Plants for the Treatment of Dengue Fever and Severe Acute Respiratory Syndrome-Coronavirus. Biomolecules, 11(1), 42. https://doi.org/10.3390/biom11010042