
Prototype Foamy Virus Integrase Displays Unique Biochemical Activities among Retroviral Integrases



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression and Purification of PFV IN
2.2. Assembly and Purification of PFV Intasomes
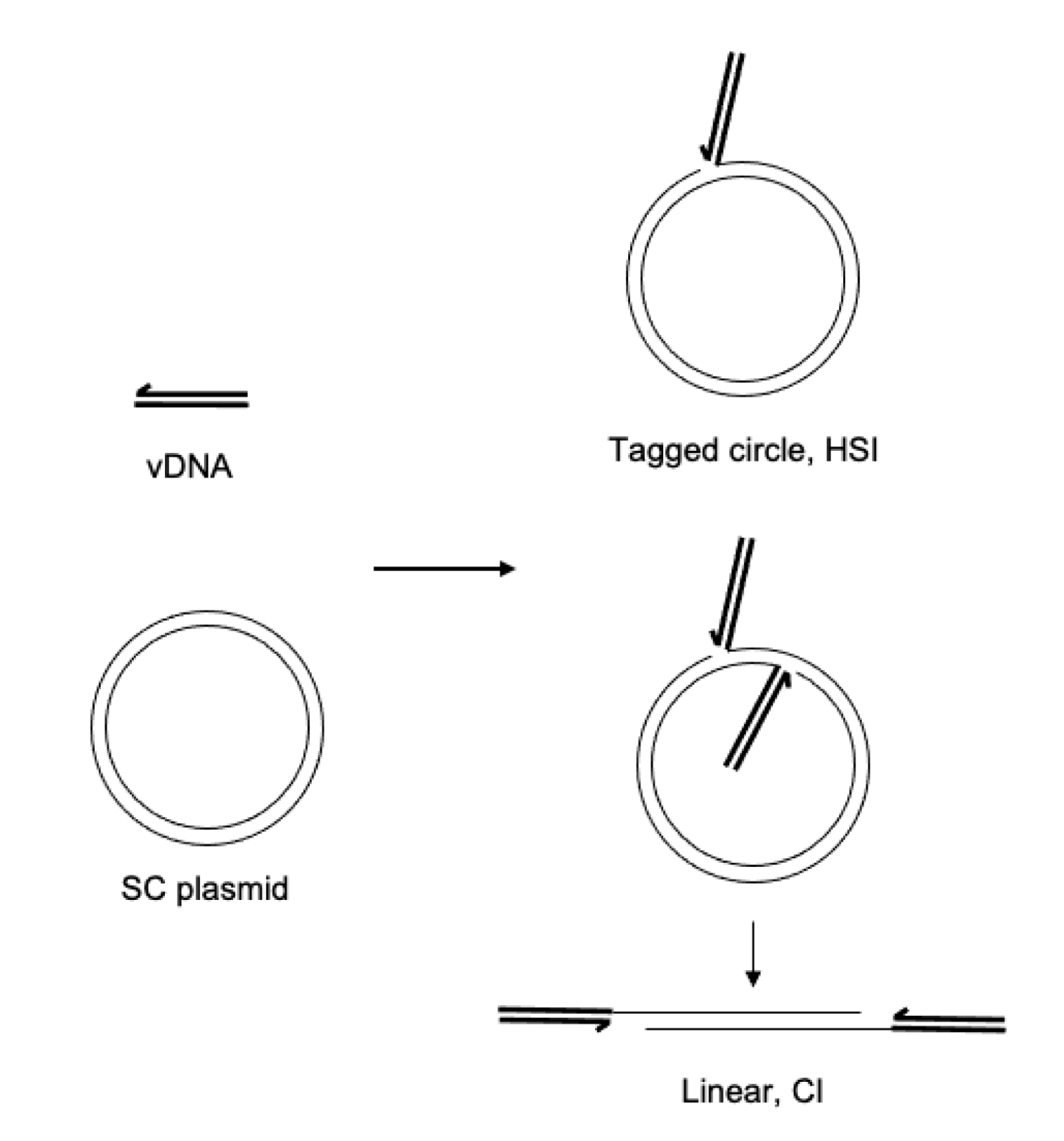
2.3. Integration Assays
3. Results
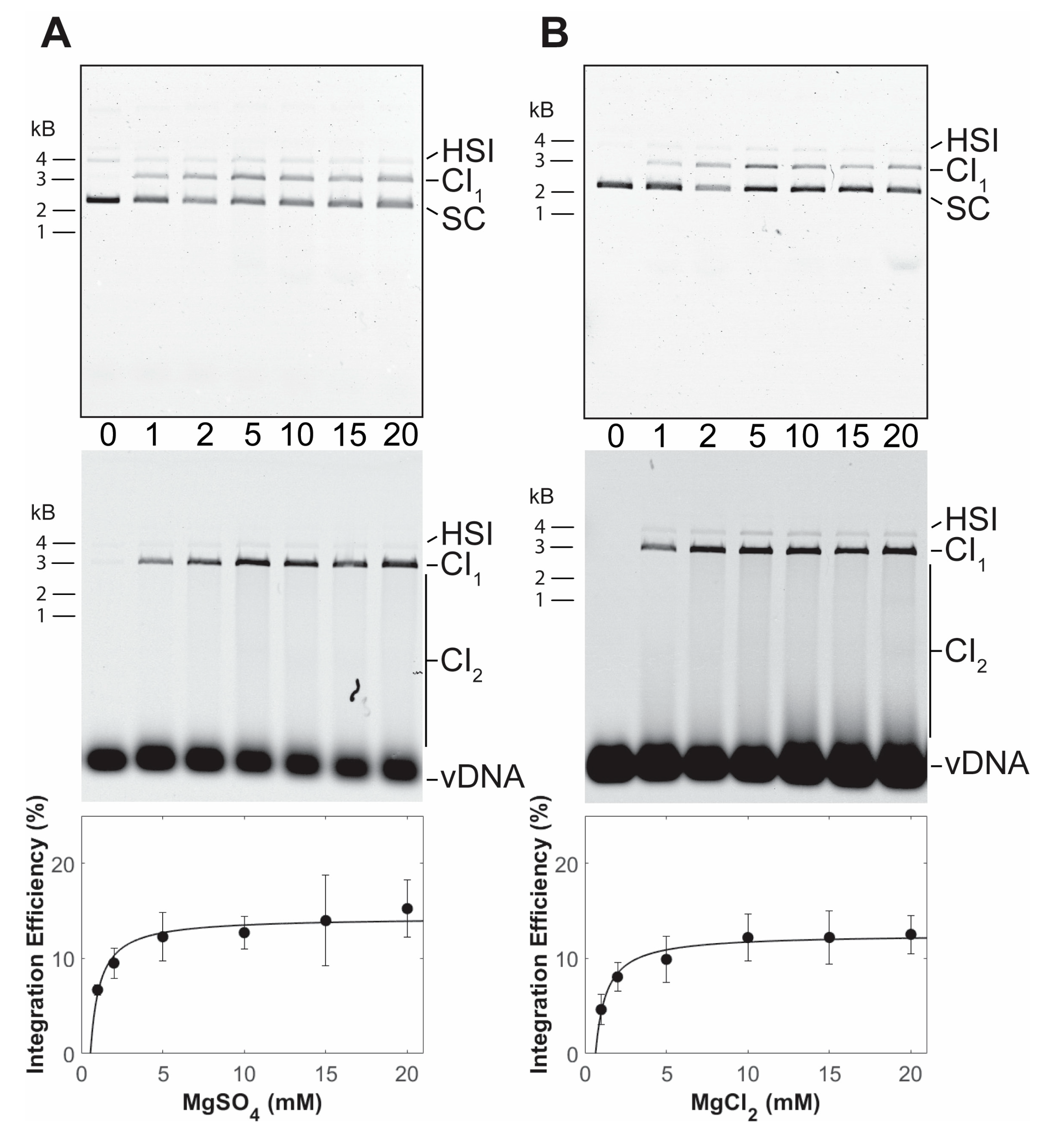
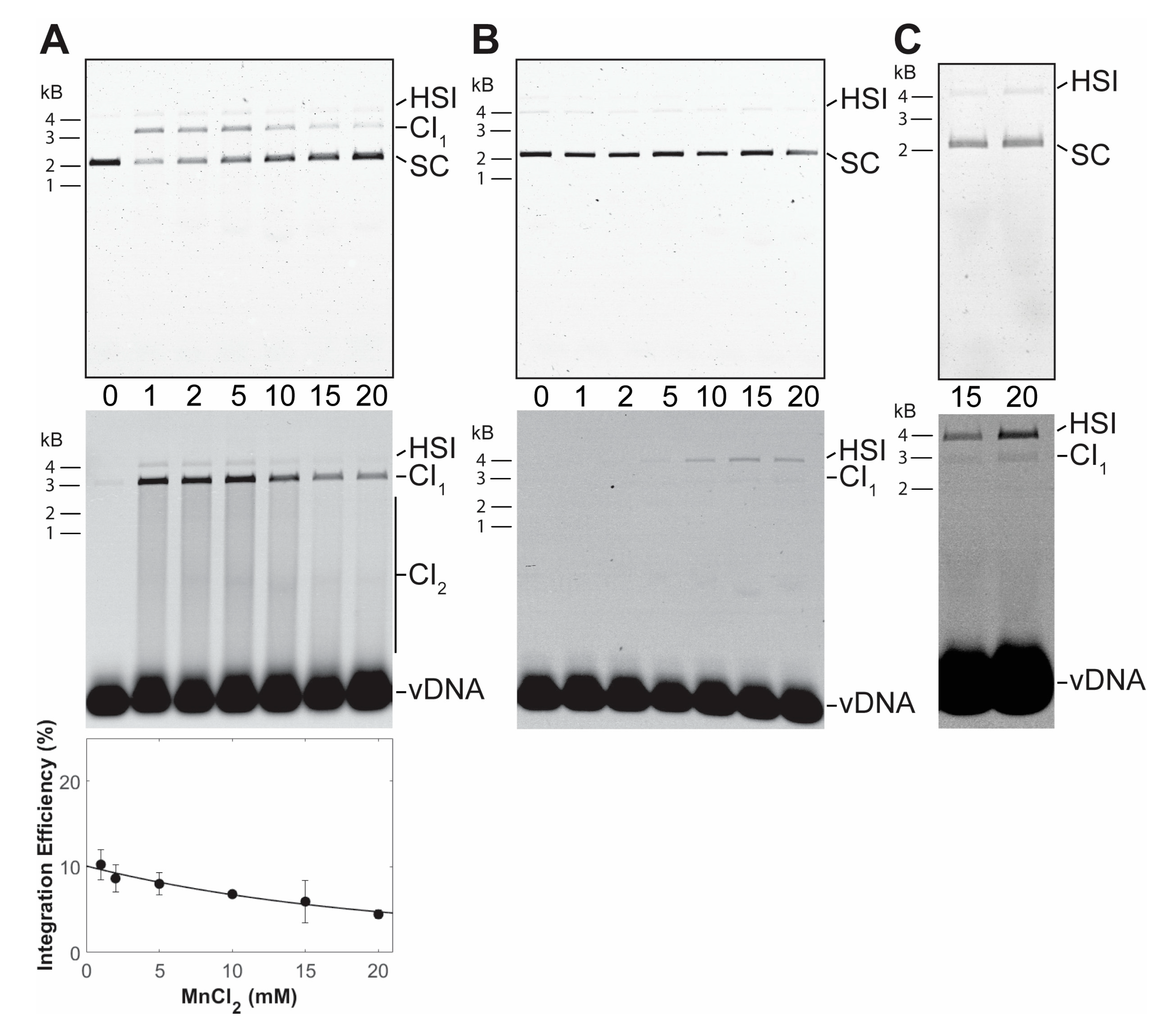
3.1. PFV Intasome Requirements for Divalent Cations
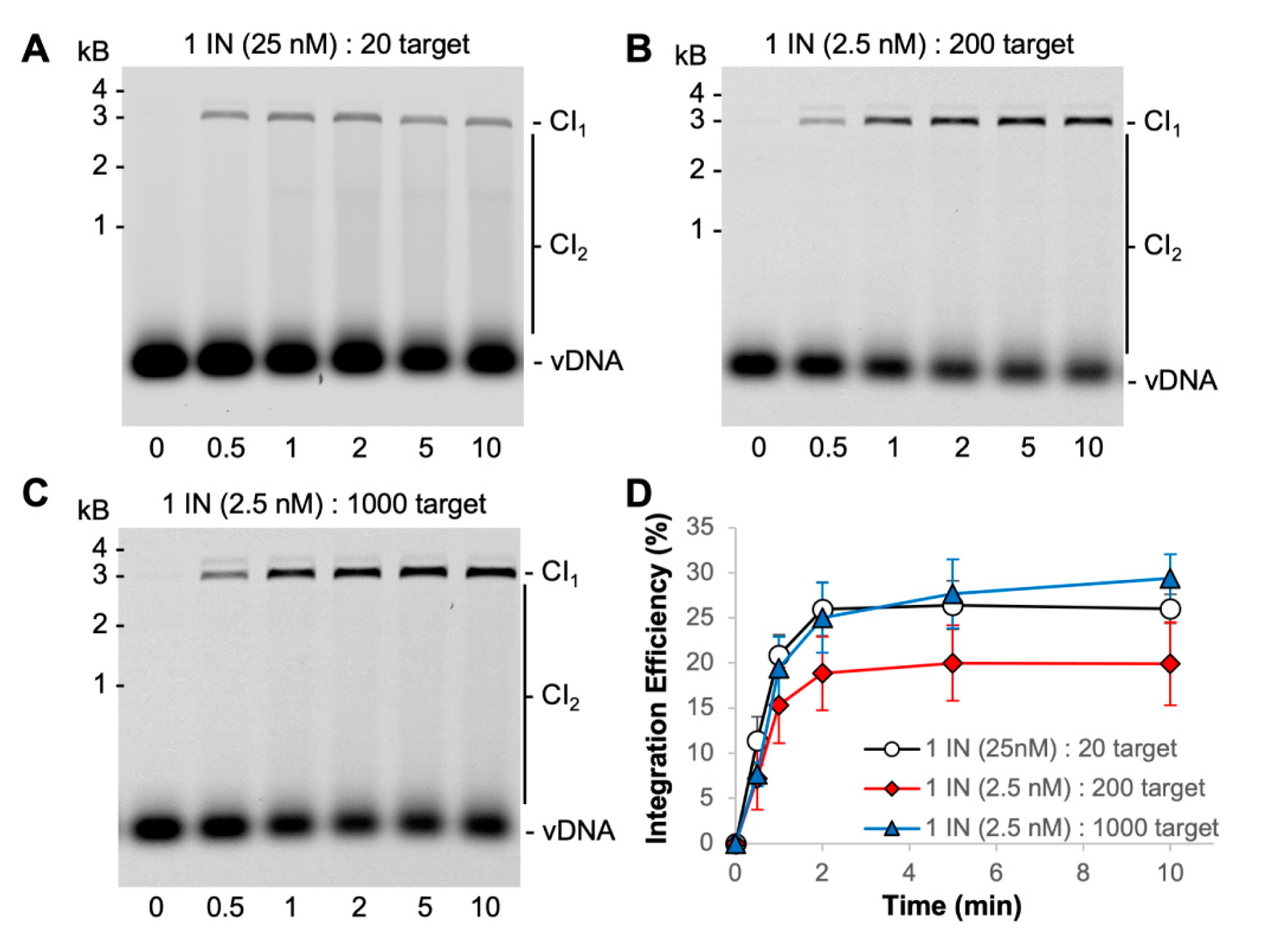
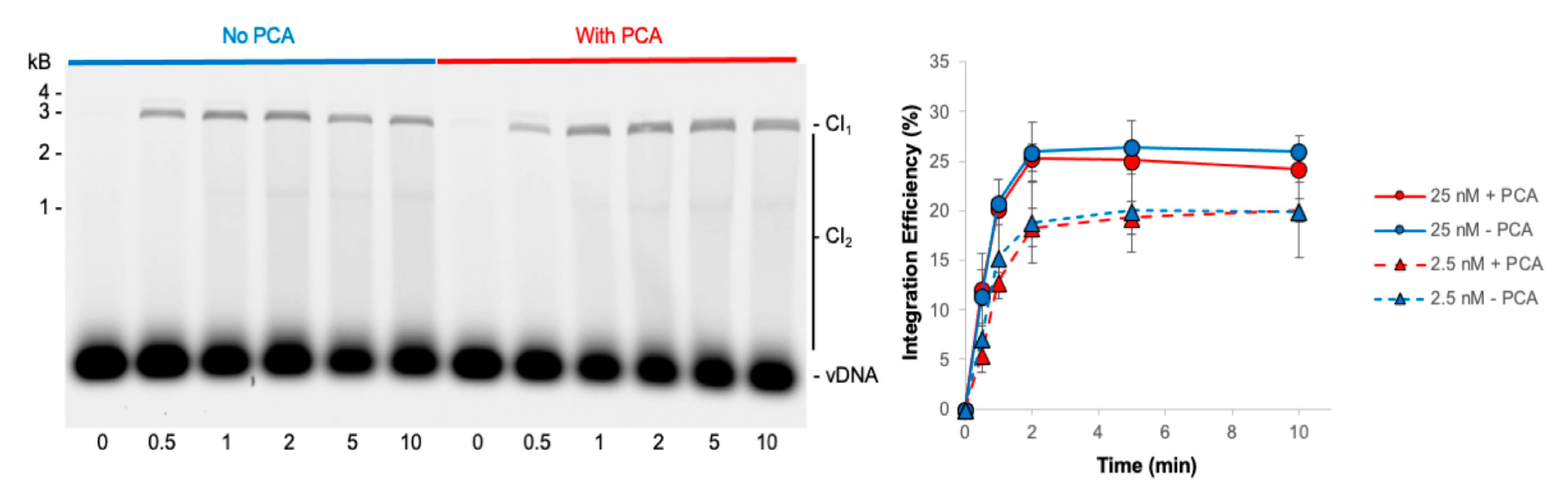
3.2. PFV Intasome Mediated Integration Is Quick
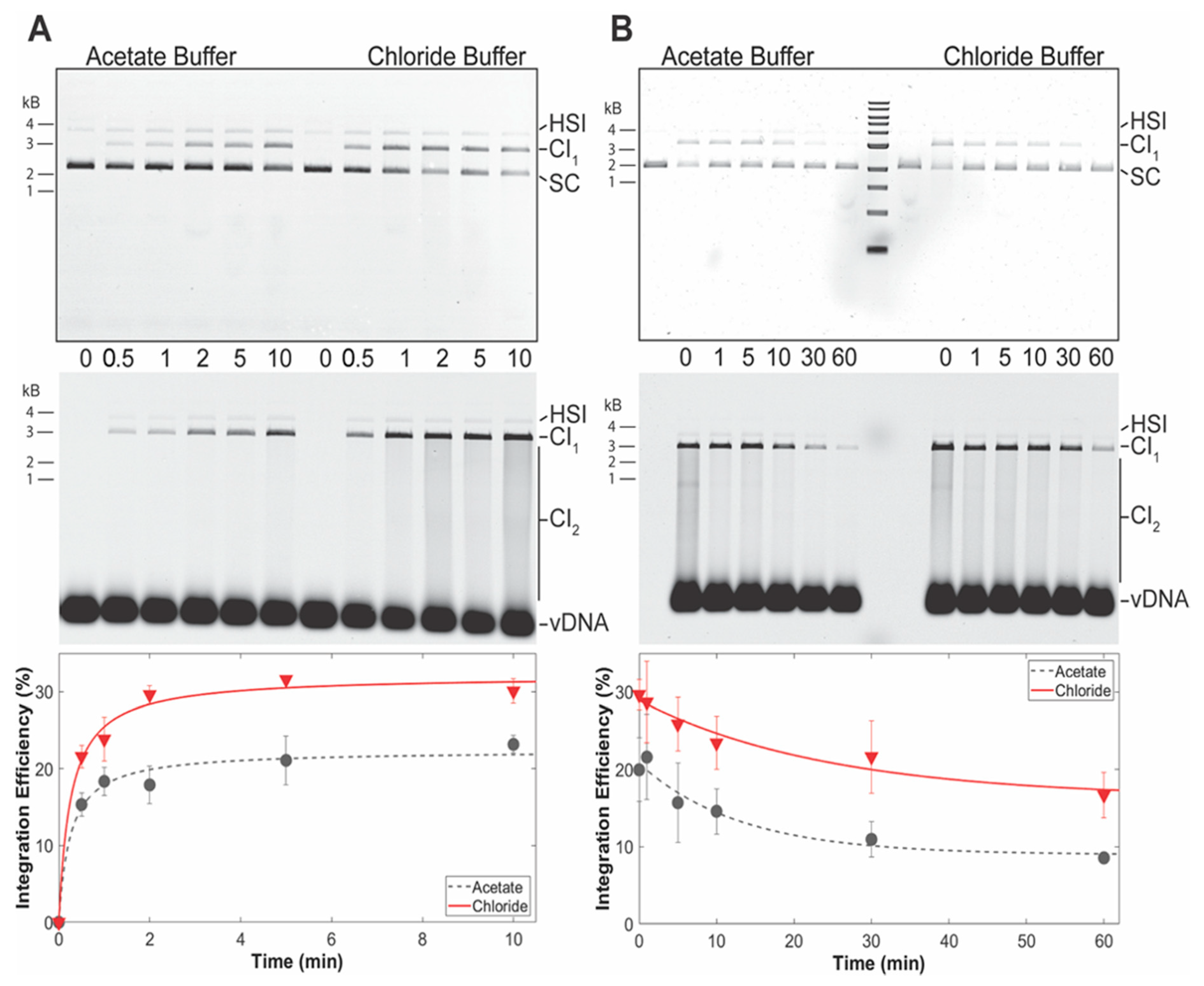
3.3. PFV Intasomes Are Less Active in the Presence of Acetate Buffer
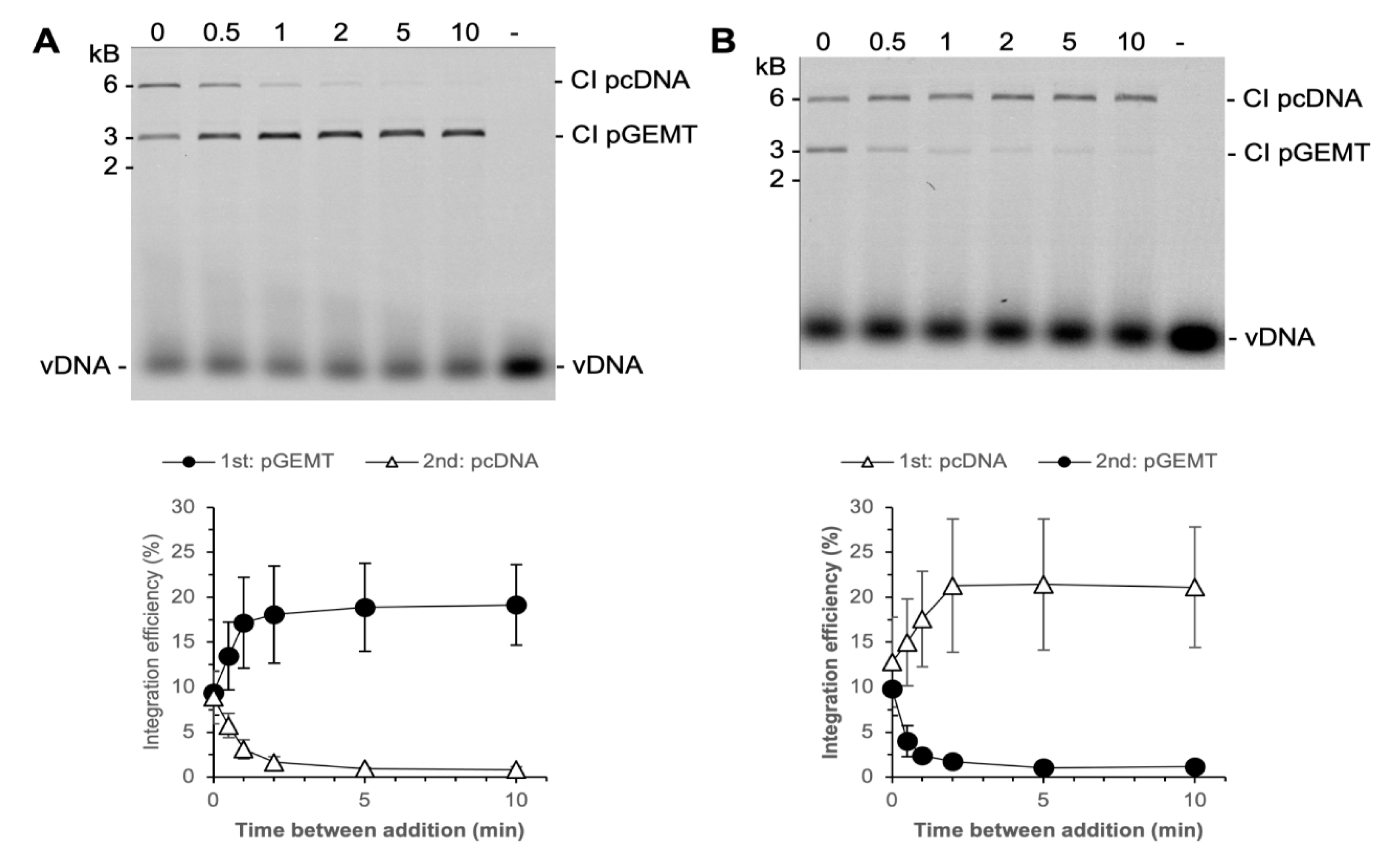
3.4. PFV Intasomes Do Not Commit to Target DNA
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coffin, J.M.; Hughes, S.H.; Varmus, H.E. Retroviruses; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Yoder, K.E.; Bushman, F.D. Repair of gaps in retroviral DNA integration intermediates. J. Virol. 2000, 74, 11191–11200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varadarajan, J.; McWilliams, M.J.; Hughes, S.H. Treatment with suboptimal doses of raltegravir leads to aberrant HIV-1 integrations. Proc. Natl. Acad. Sci. USA 2013, 110, 14747–14752. [Google Scholar] [CrossRef] [Green Version]
- Varadarajan, J.; McWilliams, M.J.; Mott, B.T.; Thomas, C.J.; Smith, S.J.; Hughes, S.H. Drug resistant integrase mutants cause aberrant HIV integrations. Retrovirology 2016, 13, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craigie, R. HIV integrase, a brief overview from chemistry to therapeutics. J. Biol. Chem. 2001, 276, 23213–23216. [Google Scholar] [CrossRef] [Green Version]
- Engelman, A.; Cherepanov, P. Retroviral Integrase Structure and DNA Recombination Mechanism. Microbiol. Spectr. 2014, 2, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Engelman, A.; Mizuuchi, K.; Craigie, R. HIV-1 DNA integration: Mechanism of viral DNA cleavage and DNA strand transfer. Cell 1991, 67, 1211–1221. [Google Scholar] [CrossRef]
- Ellison, V.; Gerton, J.; Vincent, K.A.; Brown, P.O. An essential interaction between distinct domains of HIV-1 integrase mediates assembly of the active multimer. J. Biol. Chem. 1995, 270, 3320–3326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosta, E.; Yang, W.; Hummer, G. Calcium inhibition of ribonuclease H1 two-metal ion catalysis. J. Am. Chem. Soc. 2014, 136, 3137–3144. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Lin, Z.; Lynn, A.Y.; Varnado, B.; Beutler, J.A.; Murelli, R.P.; Le Grice, S.F.; Tang, L. Two distinct modes of metal ion binding in the nuclease active site of a viral DNA-packaging terminase: Insight into the two-metal-ion catalytic mechanism. Nucleic Acids Res. 2015, 43, 11003–11016. [Google Scholar] [CrossRef] [PubMed]
- Savilahti, H.; Rice, P.A.; Mizuuchi, K. The phage Mu transpososome core: DNA requirements for assembly and function. EMBO J. 1995, 14, 4893–4903. [Google Scholar] [CrossRef] [PubMed]
- Junop, M.S.; Haniford, D.B. Multiple roles for divalent metal ions in DNA transposition: Distinct stages of Tn10 transposition have different Mg2+ requirements. EMBO J. 1996, 15, 2547–2555. [Google Scholar] [CrossRef]
- Deweese, J.E.; Osheroff, N. The use of divalent metal ions by type II topoisomerases. Metallomics 2010, 2, 450–459. [Google Scholar] [CrossRef] [Green Version]
- Mordasini, T.; Curioni, A.; Andreoni, W. Why do divalent metal ions eiTher. promote or inhibit enzymatic reactions? The case of BamHI restriction endonuclease from combined quantum-classical simulations. J. Biol. Chem. 2003, 278, 4381–4384. [Google Scholar] [CrossRef] [Green Version]
- Nowotny, M.; Yang, W. Stepwise analyses of metal ions in RNase H catalysis from substrate destabilization to product release. EMBO J. 2006, 25, 1924–1933. [Google Scholar] [CrossRef]
- Leschziner, A.E.; Griffin IV, T.J.; Grindley, N.D. Tn552 transposase catalyzes concerted strand transfer in vitro. Proc. Natl. Acad. Sci. USA 1998, 95, 7345–7350. [Google Scholar] [CrossRef] [Green Version]
- Sissi, C.; Palumbo, M. Effects of magnesium and related divalent metal ions in topoisomerase structure and function. Nucleic Acids Res. 2009, 37, 702–711. [Google Scholar] [CrossRef] [Green Version]
- Junop, M.S.; Haniford, D.B. Factors responsible for target site selection in Tn10 transposition: A role for the DDE motif in target DNA capture. EMBO J. 1997, 16, 2646–2655. [Google Scholar] [CrossRef] [Green Version]
- Hazuda, D.J.; Felock, P.J.; Hastings, J.C.; Pramanik, B.; Wolfe, A.L. Differential divalent cation requirements uncouple the assembly and catalytic reactions of human immunodeficiency virus type 1 integrase. J. Virol. 1997, 71, 7005–7011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballandras-Colas, A.; Brown, M.; Cook, N.J.; Dewdney, T.G.; Demeler, B.; Cherepanov, P.; Lyumkis, D.; Engelman, A.N. Cryo-EM reveals a novel octameric integrase structure for betaretroviral intasome function. Nature 2016, 530, 358–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballandras-Colas, A.; Maskell, D.P.; Serrao, E.; Locke, J.; Swuec, P.; Jonsson, S.R.; Kotecha, A.; Cook, N.J.; Pye, V.E.; Taylor, I.A.; et al. A supramolecular assembly mediates lentiviral DNA integration. Science 2017, 355, 93–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hare, S.; Gupta, S.S.; Valkov, E.; Engelman, A.; Cherepanov, P. Retroviral intasome assembly and inhibition of DNA strand transfer. Nature 2010, 464, 232–236. [Google Scholar] [CrossRef]
- Mackler, R.M.; Lopez, M.A., Jr.; Yoder, K.E. Assembly and Purification of Prototype Foamy Virus Intasomes. J. Vis. Exp. 2018, 133, e57453. [Google Scholar] [CrossRef]
- Andrake, M.D.; Ramcharan, J.; Merkel, G.; Zhao, X.Z.; Burke, T.R., Jr.; Skalka, A.M. Comparison of metal-dependent catalysis by HIV-1 and ASV integrase proteins using a new and rapid, moderate throughput assay for joining activity in solution. AIDS Res. Ther. 2009, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Mackler, R.M.; Lopez, M.A.; Osterhage, M.J.; Yoder, K.E. Prototype foamy virus integrase is promiscuous for target choice. Biochem. Biophys. Res. Commun. 2018, 503, 1241–1246. [Google Scholar] [CrossRef]
- Miller, M.D.; Bor, Y.C.; Bushman, F. Target DNA capture by HIV-1 integration complexes. Curr. Biol. 1995, 5, 1047–1056. [Google Scholar] [CrossRef] [Green Version]
- Lopez, M.A., Jr.; Mackler, R.M.; Altman, M.P.; Yoder, K.E. Detection and Removal of Nuclease Contamination during Purification of Recombinant Prototype Foamy Virus Integrase. J. Vis. Exp. JoVE 2017, 130, e56605. [Google Scholar] [CrossRef]
- Lopez, M.A., Jr.; Mackler, R.M.; Yoder, K.E. Removal of nuclease contamination during purification of recombinant prototype foamy virus integrase. J. Virol. Methods 2016, 235, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Valkov, E.; Gupta, S.S.; Hare, S.; Helander, A.; Roversi, P.; McClure, M.; Cherepanov, P. Functional and structural characterization of the integrase from the prototype foamy virus. Nucleic Acids Res. 2009, 37, 243–255. [Google Scholar] [CrossRef]
- Wolfe, A.L.; Felock, P.J.; Hastings, J.C.; Blau, C.U.; Hazuda, D.J. The role of manganese in promoting multimerization and assembly of human immunodeficiency virus type 1 integrase as a catalytically active complex on immobilized long terminal repeat substrates. J. Virol. 1996, 70, 1424–1432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, N.D.; Mackler, R.M.; Lopez, M.A., Jr.; Baltierra-Jasso, L.E.; Altman, M.P.; Senavirathne, G.; Yoder, K.E. Prototype foamy virus intasome aggregation is mediated by outer protein domains and prevented by protocatechuic acid. Sci. Rep. 2019, 9, 132. [Google Scholar] [CrossRef] [PubMed]
- Kotlar, R.M.; Jones, N.D.; Senavirathne, G.; Gardner, A.M.; Messer, R.K.; Tan, Y.Y.; Rabe, A.J.; Fishel, R.; Yoder, K.E. Retroviral prototype foamy virus intasome binding to a nucleosome target does not determine integration efficiency. J. Biol. Chem. 2021, 296, 100550. [Google Scholar] [CrossRef] [PubMed]
- Maertens, G.N.; Hare, S.; Cherepanov, P. The mechanism of retroviral integration from X-ray structures of its key intermediates. Nature 2010, 468, 326–329. [Google Scholar] [CrossRef] [Green Version]
- Jones, N.D.; Lopez, M.A., Jr.; Hanne, J.; Peake, M.B.; Lee, J.B.; Fishel, R.; Yoder, K.E. Retroviral intasomes search for a target DNA by 1D diffusion which rarely results in integration. Nat. Commun. 2016, 7, 11409. [Google Scholar] [CrossRef]
- Maskell, D.P.; Renault, L.; Serrao, E.; Lesbats, P.; Matadeen, R.; Hare, S.; Lindemann, D.; Engelman, A.N.; Costa, A.; Cherepanov, P. Structural basis for retroviral integration into nucleosomes. Nature 2015, 523, 366–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benleulmi, M.S.; Matysiak, J.; Henriquez, D.R.; Vaillant, C.; Lesbats, P.; Calmels, C.; Naughtin, M.; Leon, O.; Skalka, A.M.; Ruff, M.; et al. Intasome architecture and chromatin density modulate retroviral integration into nucleosome. Retrovirology 2015, 12, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Lin, S.; Craigie, R. Outer domains of integrase within retroviral intasomes are dispensible for catalysis of DNA integration. Protein Sci. 2016, 25, 472–478. [Google Scholar] [CrossRef] [Green Version]
- Pasi, M.; Mornico, D.; Volant, S.; Juchet, A.; Batisse, J.; Bouchier, C.; Parissi, V.; Ruff, M.; Lavery, R.; Lavigne, M. DNA minicircles clarify the specific role of DNA structure on retroviral integration. Nucleic Acids Res. 2016, 44, 7830–7847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Vargas, E.; Larsen, L.; Knapp, W.; Hatfield, G.W.; Lathrop, R.; Sandmeyer, S. Directed DNA shuffling of retrovirus and retrotransposon integrase protein domains. PLoS ONE 2013, 8, e63957. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.D.; Renault, L.; Maskell, D.P.; Ghoneim, M.; Pye, V.E.; Nans, A.; Rueda, D.S.; Cherepanov, P.; Costa, A. Retroviral integration into nucleosomes through DNA looping and sliding along the histone octamer. Nat. Commun. 2019, 10, 4189. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Lapkouski, M.; Yang, W.; Craigie, R. Assembly of prototype foamy virus strand transfer complexes on product DNA bypassing catalysis of integration. Protein Sci. 2012, 21, 1849–1857. [Google Scholar] [CrossRef] [Green Version]
- Vanderlinden, W.; Brouns, T.; Walker, P.U.; Kolbeck, P.J.; Milles, L.F.; Ott, W.; Nickels, P.C.; Debyser, Z.; Lipfert, J. The free energy landscape of retroviral integration. Nat. Commun. 2019, 10, 4738. [Google Scholar] [CrossRef] [PubMed]
- Engelman, A.N.; Cherepanov, P. Retroviral intasomes arising. Curr. Opin. Struct. Biol. 2017, 47, 23–29. [Google Scholar] [CrossRef]
- Hare, S.; Maertens, G.N.; Cherepanov, P. 3’-processing and strand transfer catalysed by retroviral integrase in crystallo. EMBO J. 2012, 31, 3020–3028. [Google Scholar] [CrossRef] [Green Version]
- Engelman, A.; Craigie, R. Efficient magnesium-dependent human immunodeficiency virus type 1 integrase activity. J. Virol. 1995, 69, 5908–5911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellison, V.; Abrams, H.; Roe, T.; Lifson, J.; Brown, P. Human immunodeficiency virus integration in a cell-free system. J. Virol. 1990, 64, 2711–2715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farnet, C.M.; Haseltine, W.A. Integration of human immunodeficiency virus type 1 DNA in vitro. Proc. Natl. Acad. Sci. USA 1990, 87, 4164–4168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltierra-Jasso, L.E.; Jones, N.D.; Ballandras-Colas, A.; Engelman, A.N.; Fishel, R.; Yoder, K.E. The mouse mammary tumor virus intasome exhibits distinct dynamics on target DNA. bioRxiv 2021, 468995. [Google Scholar] [CrossRef]
- Bonnet, I.; Biebricher, A.; Porte, P.L.; Loverdo, C.; Benichou, O.; Voituriez, R.; Escude, C.; Wende, W.; Pingoud, A.; Desbiolles, P. Sliding and jumping of single EcoRV restriction enzymes on non-cognate DNA. Nucleic Acids Res. 2008, 36, 4118–4127. [Google Scholar] [CrossRef] [Green Version]
- Serrao, E.; Ballandras-Colas, A.; Cherepanov, P.; Maertens, G.N.; Engelman, A.N. Key determinants of target DNA recognition by retroviral intasomes. Retrovirology 2015, 12, 39. [Google Scholar] [CrossRef] [Green Version]
- Lesbats, P.; Serrao, E.; Maskell, D.P.; Pye, V.E.; O’Reilly, N.; Lindemann, D.; Engelman, A.N.; Cherepanov, P. Structural basis for spumavirus GAG tethering to chromatin. Proc. Natl. Acad. Sci. USA 2017, 114, 5509–5514. [Google Scholar] [CrossRef] [Green Version]
- Cherepanov, P.; Maertens, G.; Proost, P.; Devreese, B.; Van Beeumen, J.; Engelborghs, Y.; De Clercq, E.; Debyser, Z. HIV-1 integrase forms stable tetramers and associates with LEDGF/p75 protein in human cells. J. Biol. Chem. 2003, 278, 372–381. [Google Scholar] [CrossRef] [Green Version]
- Shun, M.C.; Raghavendra, N.K.; Vandegraaff, N.; Daigle, J.E.; Hughes, S.; Kellam, P.; Cherepanov, P.; Engelman, A. LEDGF/p75 functions downstream from preintegration complex formation to effect gene-specific HIV-1 integration. Genes Dev. 2007, 21, 1767–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linial, M. Why aren’t foamy viruses pathogenic? Trends Microbiol. 2000, 8, 284–289. [Google Scholar] [CrossRef]
- Bhatt, V.; Shi, K.; Salamango, D.J.; Moeller, N.H.; Pandey, K.K.; Bera, S.; Bohl, H.O.; Kurniawan, F.; Orellana, K.; Zhang, W.; et al. Structural basis of host protein hijacking in human T-cell leukemia virus integration. Nat. Commun. 2020, 11, 3121. [Google Scholar] [CrossRef]
- Passos, D.O.; Li, M.; Yang, R.; Rebensburg, S.V.; Ghirlando, R.; Jeon, Y.; Shkriabai, N.; Kvaratskhelia, M.; Craigie, R.; Lyumkis, D. Cryo-EM structures and atomic model of the HIV-1 strand transfer complex intasome. Science 2017, 355, 89–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Z.; Shi, K.; Banerjee, S.; Pandey, K.K.; Bera, S.; Grandgenett, D.P.; Aihara, H. Crystal structure of the Rous sarcoma virus intasome. Nature 2016, 530, 362–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabe, A.J.; Tan, Y.Y.; Larue, R.C.; Yoder, K.E. Prototype Foamy Virus Integrase Displays Unique Biochemical Activities among Retroviral Integrases. Biomolecules 2021, 11, 1910. https://doi.org/10.3390/biom11121910
Rabe AJ, Tan YY, Larue RC, Yoder KE. Prototype Foamy Virus Integrase Displays Unique Biochemical Activities among Retroviral Integrases. Biomolecules. 2021; 11(12):1910. https://doi.org/10.3390/biom11121910
Chicago/Turabian StyleRabe, Anthony J., Yow Yong Tan, Ross C. Larue, and Kristine E. Yoder. 2021. "Prototype Foamy Virus Integrase Displays Unique Biochemical Activities among Retroviral Integrases" Biomolecules 11, no. 12: 1910. https://doi.org/10.3390/biom11121910
APA StyleRabe, A. J., Tan, Y. Y., Larue, R. C., & Yoder, K. E. (2021). Prototype Foamy Virus Integrase Displays Unique Biochemical Activities among Retroviral Integrases. Biomolecules, 11(12), 1910. https://doi.org/10.3390/biom11121910