
Recent Insights into Anthocyanin Pigmentation, Synthesis, Trafficking, and Regulatory Mechanisms in Rice (Oryza sativa L.) Caryopsis


 and
and
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bran Color | Total Anthocyanin (CGE/100 g) | Total Proanthocyanidin (mg CAE/100 g) |
|---|---|---|
| Black | 1884 | 78 |
| Purple | 2874 | 525.4 |
| Red | 8.78 | 716.6 |
| Brown | 3.09 | 4.34 |
| Cultivar’s Name | Type of Anthocyanin | Methods Used | References |
|---|---|---|---|
| Baoji | Nine anthocyanins Minor Cyanidin-3,5-Glucoside, Cyanidin-3-Gentiobioside Cyanidin-3-Rutinoside Cyanidin-3-Sambubioside Major Cyanidin-3-Glucoside Peonidin-3-Glucoside Peonidin Cyanidin Cyanidin-derivative). | -High Performance Liquid Chromatography (HPLC) -Ultra-Performance Liquid Chromatography/time-of-Flight Mass Spectrometry (UPLC/Q-TOF-MS) | [15] |
| Longjin N°1 | Four different anthocyanins Cyanidin-3-Glucoside Peonidin-3-Glucoside Cyanidin-3,5-Diglucoside Cyanidin-3-Rutinoside | -High Performance Liquid Chromatography (HPLC) -Electrospray ionization -Mass Spectrometry with diode array detection. | [1] |
| Okunomurasaki, Chinakuromai and Asamurasaki | Cyanidin-3-Glucoside, Peonidin-3- Glucoside Petunidin-3-Glucoside Malvidin | -High Performance Liquid Chromatography (HPLC) | [3] |
| Rice berry | Cyanidin-3-Glucoside Peonidin-3- Glucoside, Cyanidin, Cyanidin-3,5-Diglucoside | -Mass Spectrometric | [38] |
| 25 rice varieties: Cabaysay, Cheng Chang, Hung Tsan, Longqing No. 3, Mitak, etc. | Cyanidin-3-Galactoside, Peonidin-3-Glucoside, Cyanidin-3-Glucoside, Cyanidin-3-Rutinoside, Cyanidin, and Peonidin | Identification based on retention times four anthocyanins and two anthocyanidins | [40] |
| Yunanheixiannuo | Cyanidin-3-Glucoside Cyanidin-3-Rutinoside Peonidin-3- Glucoside | -Liquid Chromatography- Mass Spectrometry (LC-MS) | [31] |
| Venere | Cyanidin-O-diglucoside, Cyanidin-3-Glucoside Cyanidin-3-Rutinoside Peonidin-3-Glucoside Peonidin-3-Rutinoside | -Liquid Chromatography- Mass Spectrometry (LC-MS) | [34] |
| Dongjin | Cyanidin-3-Glucoside Peonidin-3-Glucoside Cyanidin-3-Gentiobioside Cyanidin-3-Rhamnoside Cyanidin-3,5-Diglucoside Cyanidin-3-Rhamnoglucoside Peonidin-3-Rhamnoglucoside. | High Performance Liquid Chromatography (HPLC) | [41] |
| Cultivar’s Name | Total Anthocyanin Content (TAC) | Major Anthocyanin | References |
|---|---|---|---|
| Venere | 334 µg/g DW | Cyanidin3-O-glucoside (78% of TAC) | [34] |
| Baoji | 416.92 mg CGE/g | Cyanidin 3-O-glucoside (76.5% of TAC) | [15] |
| Canada sweet rice | 3276 µg/g DW | Cyanidin 3-O-glucoside (88% of TAC) | [42] |
| O. sativa japonica var SBR | 630 µg/g DW | Cyanidin 3-O-glucoside (572.47 µg/g; 91.13% of TAC) | [43] |
| Artemide, Nerone, Venere | 1404, 812 and 780 µg/g, respectively | Cyanidin3-O-glucoside (71%, 68%, 65% of TAC respectively) | [44] |
| rice berry | 37 mg/100g DW | Cyanidin 3-O-glucoside Peonidin 3-O-glucoside | [38] |
| Yunanheixiannuo | 7.5 mgCGE/g DM | Cyanidin 3-O-glucoside (76.8% of TAC) 85% of TAC in bran, no anthocyanin in the endosperm | [31] |
| Okunomurasaki, Chinakuromai, and Asamurasaki | 79.5–473.7 mg/100g DM | Cyanidin 3-O-glucoside more than 55% of TAC | [3] |
| 25 rice varieties: Cabaysay, Cheng Chang, Hung Tsan, Longqing No. 3, Mitak, etc. | 79.5–473.7 mg/100 g DM | Cyanidin 3-O-glucoside ( about 82% of TAC) Anthocyanin is mainly found in bran | [40] |
| BIR1-3 and BJR1-3 | 32.4 and 160.1 mg/100g DW, respectively | Cyanidin 3-O-glucoside (65.62% and 89.24% respectively) | [45] |
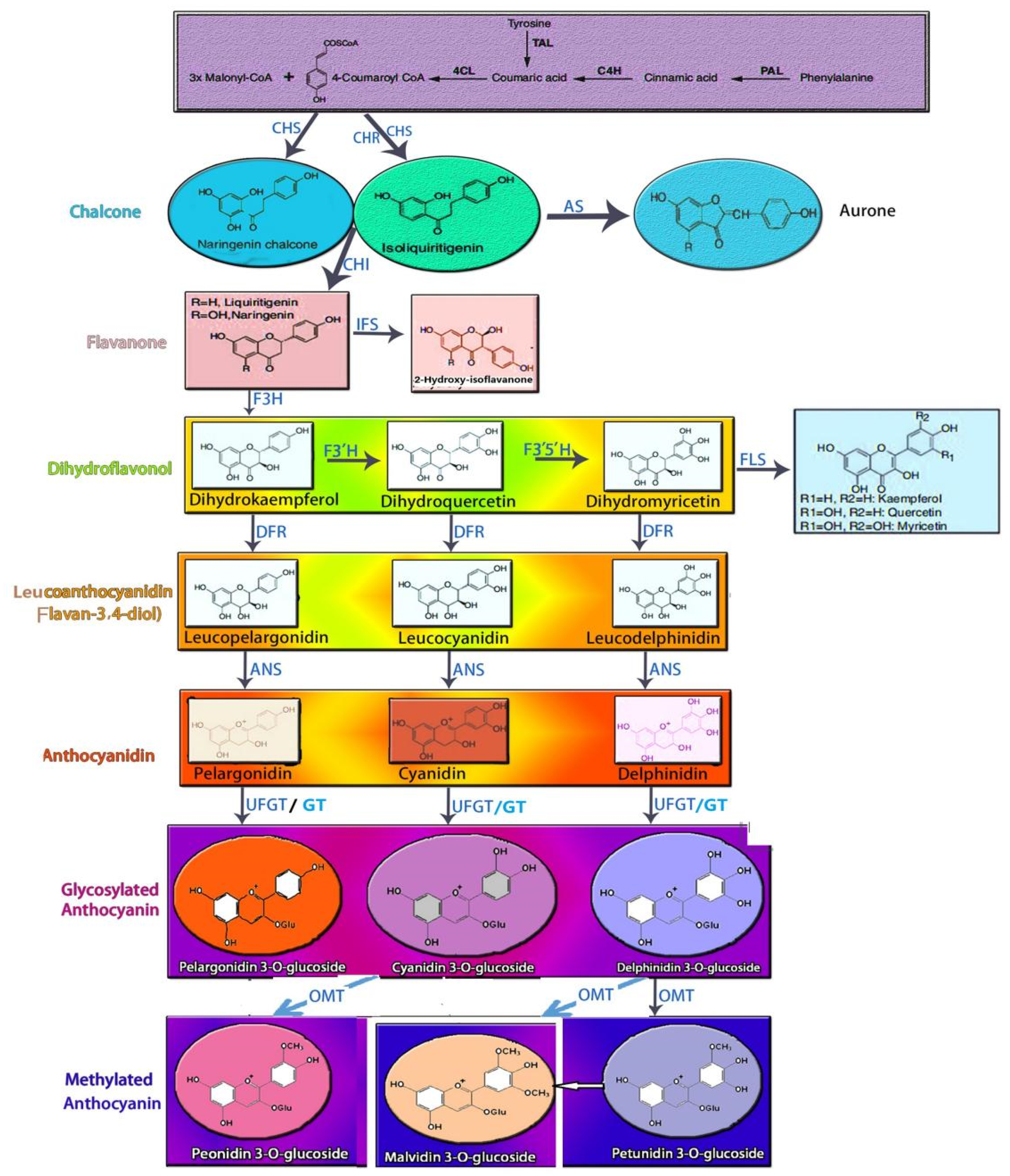
2. Biosynthesis of Anthocyanin in the Rice Caryopsis and Its Relevant Mediators
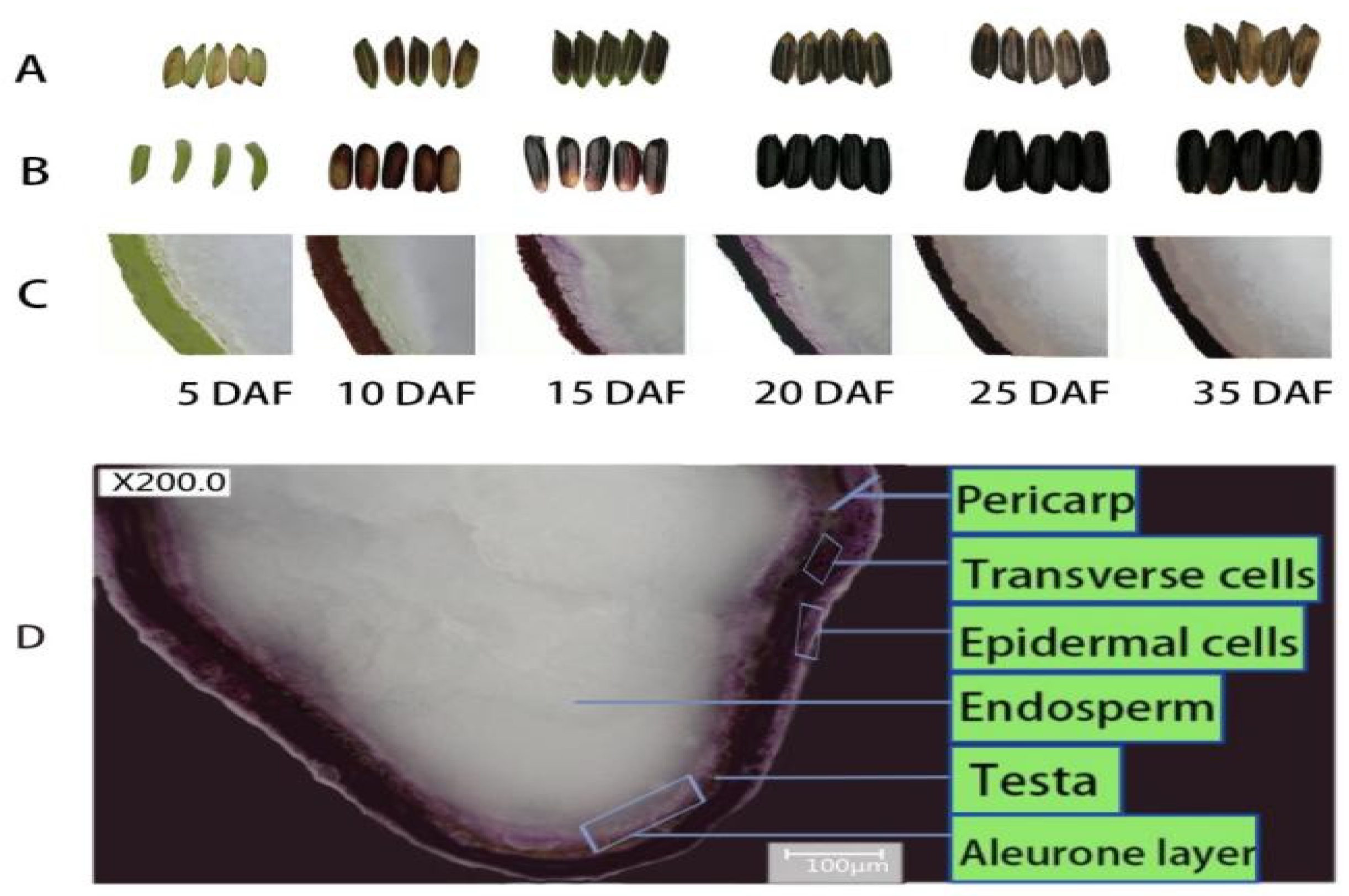
3. Trafficking and Accumulation of Anthocyanin in the Rice Caryopsis
4. Regulation of Anthocyanin in the Rice Caryopsis
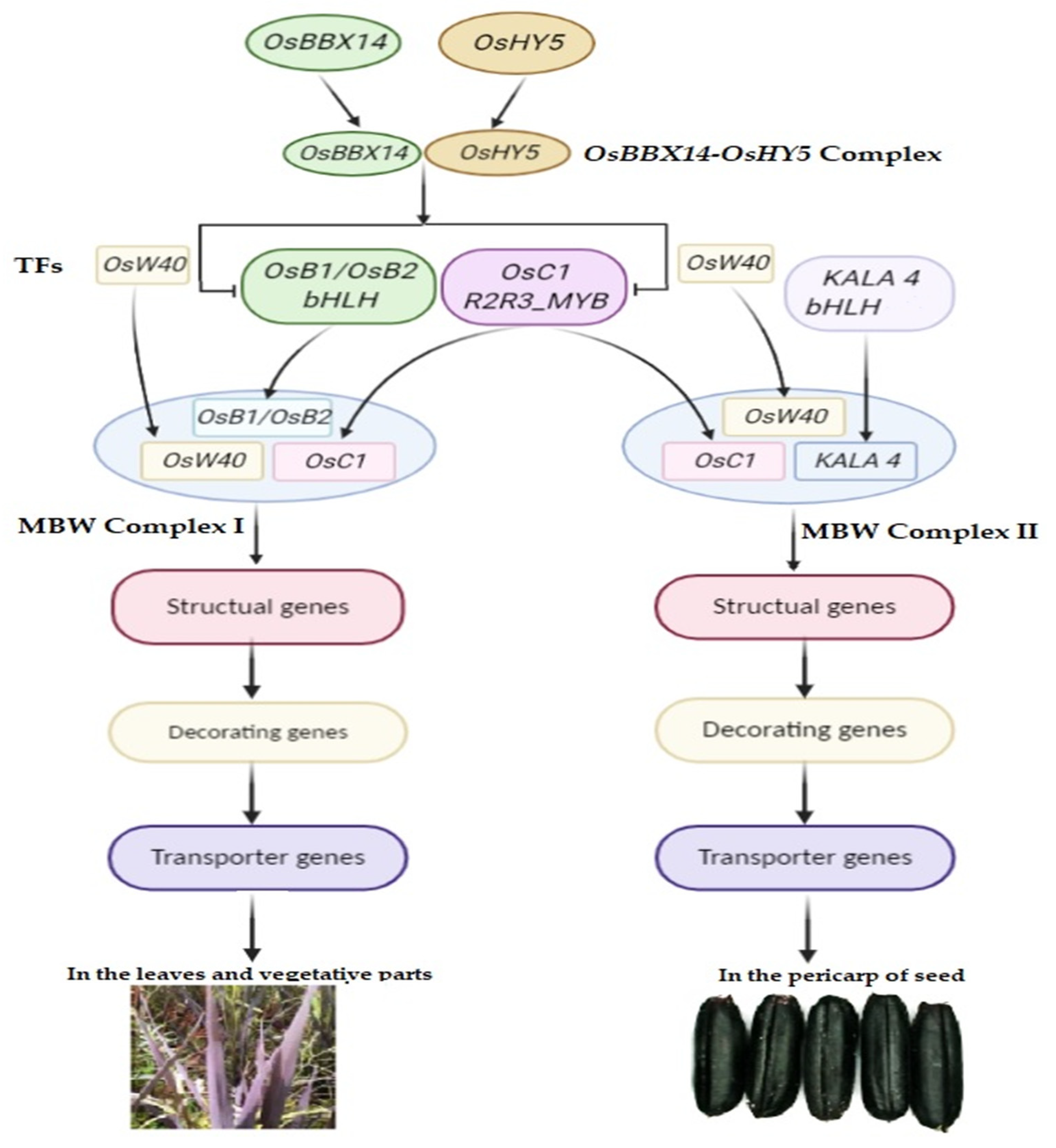
4.1. The MYB-bHLH-WD40 (MBW) Complex and Activation of the Anthocyanin Biosynthesis Pathway in Rice
4.2. Black Rice Caryopsis Pigmentation: From Classical Genetics to Molecular Genetics
4.3. Molecular Regulation of Anthocyanin Production in the Rice Caryopsis
5. Enhanced Anthocyanin Accumulation in Rice Caryopses through Genetic and Environmental Modulation
5.1. Genetic Engineering Approach
5.2. Environmental Modulation
5.2.1. Chemical Treatment
5.2.2. Temperature
5.2.3. Light
5.2.4. Salinity or Salt Stress
5.2.5. Nutrient Supply
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 4 CL | 4-Coumaroyl CoA Ligase |
| ABC | ATP-Binding Cassette |
| AM | Anthomate |
| ANR | Anthocyanidin reductase |
| ANS | Anthocyanin synthase |
| APV | Anthocyanic pre-vacuolar |
| AVI | Anthocyanic vacuolar intrusion |
| Bhlh | Basic helix-loop-helix |
| BUN | Blood Urea Nitrogen |
| C3G | Cyanidin-3-glucoside |
| C3R | Cyanidin 3-Rutinoside |
| C4H | Cinnamate 4-hydroxylase |
| CAE | Catechin acid equivalent |
| CGE | Cyanidin 3-o-glucoside equivalent, |
| Chr | Chromosome |
| CRISPR/Cas9 | Clustered Regularly Interspaced Short palindromic Repeats/CRISPR-associated protein9 |
| DAD | Diode array detectors |
| DAF | Days after flowering |
| DM | Dry matter |
| DW | Dry weight |
| EBG | Early biosynthesis genes |
| ER | Endoplasmic reticulum |
| F 3′5′H | Flavonoid 3′-5′ hydroxylase |
| F3H | Flavanone 3-hydroxylase |
| FLS | Flavonol synthase |
| GST | Glutathione-S-Transferase |
| KALA | Key Activator Loci for Anthocyanin |
| LAR | Leuco-Anthocyanidin Reductase |
| LC-MS | Liquid Chromatography-Mass Spectrometry |
| LDOX | Leucoanthocyanidin dioxygenase |
| LT | Ligandin transportations |
| MATE | Multidrug and toxic compound extrusion |
| MDA | Malondialdehyde |
| NO | Nitric Oxide |
| ORF | Open reading frame |
| OsBBX14 | Oryza sativa B-box14 |
| OsHY5 | Oryza sativa Hypocotyl 5 |
| P3G | Peonidin-3-glucoside |
| PAL | Phenylalanine ammonia-lyase, |
| PVC | Pre-vacuolar compartment |
| QTL | Quantitative trait loci |
| ROS | Reactive oxygen species |
| TAC | Total anthocyanin content |
| u-HPLC | Ultra-High-performance liquid chromatography |
| UFGT | Uridine flavonoid 3-o-glycosyltransferases |
| UVD | Ultraviolet detection |
| UV-vis | ultraviolet-visible spectroscopy |
| VLB | Vesicle-like bodies |
| VT | Vesicular transport |
References
- Hou, Z.; Qin, P.; Zhang, Y.; Cui, S.; Ren, G. Identification of anthocyanins isolated from black rice (Oryza sativa L.) and their degradation kinetics. Food Res. Int. 2013, 50, 691–697. [Google Scholar] [CrossRef]
- Kocic, B.; Filipovic, S.; Nikolic, M.; Petrovic, B. Effects of anthocyanins and anthocyanin-rich extracts on the risk for cancers of the gastrointestinal tract. Off. J. Balk. Union Oncol. 2011, 16, 602–608. [Google Scholar]
- Chen, X.Q.; Nagao, N.; Itani, T.; Irifune, K. Anti-oxidative analysis, and identification and quantification of anthocyanin pigments in different coloured rice. Food Chem. 2012, 135, 2783–2788. [Google Scholar] [CrossRef]
- Dixon, R.A.; Liu, C.; Jun, J.H. Metabolic engineering of anthocyanins and condensed tannins in plants. Curr. Opin. Biotech. 2013, 24, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple functional roles of anthocyanins in plant-environment interactions. Environ. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Pojer, E.; Mattivi, F.; Johnson, D.; Stockley, C. The case for anthocyanin consumption to promote human health: A review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 483–508. [Google Scholar] [CrossRef] [PubMed]
- Hemamalini, S.; Umamaheswari, D.S.; Lavanya, D.R.; Umamaheswara, R.D.C. Exploring the therapeutic potential and nutritional properties of ‘KaruppuKavuni’ variety rice of Tamil Nadu. Int. J. Pharma Bio Sci. 2018, 9, 88–96. [Google Scholar] [CrossRef]
- Konczak, I.; Zhang, W. Anthocyanins—more than nature’s colours. J. Biomed. Biotechnol. 2004, 239–240. [Google Scholar] [CrossRef] [Green Version]
- Yazhen, S.; Wenju, W.; Panpan, Z.; Yuanyuan, Y.; Panpan, D.; Wusen, Z.; Yanling, W. Anthocyanins: Novel Antioxidants in Diseases Prevention and Human Health. In Flavonoids-A Coloring Model for Cheering Up Life; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Ling, W.; Wang, Q.; Liu, C.; Hu, Y.; Xia, M.; Feng, X.; Xia, X. Effect of anthocyanin-rich extract from black rice (Oryza sativa L. indica) on hyperlipidemia and insulin resistance in fructose-fed rats. Plant Foods Hum. Nutr. 2007, 62, 1–6. [Google Scholar] [CrossRef]
- Chen, P.-N.; Chu, S.-C.; Chiou, H.-L.; Chiang, C.-L.; Yang, S.-F.; Hsieh, Y. Cancer, Cyanidin 3-glucoside and peonidin 3-glucoside inhibit tumor cell growth and induce apoptosis in vitro and suppress tumor growth in vivo. J. Nutr. 2005, 53, 232–243. [Google Scholar]
- Luo, L.-P.; Han, B.; Yu, X.-P.; Chen, X.-Y.; Zhou, J.; Chen, W.; Zhu, Y.-F.; Peng, X.-L.; Zou, Q.; Li, S.-Y. Anti-metastasis activity of black rice anthocyanins against breast cancer: Analyses using an ErbB2 positive breast cancer cell line and tumoral xenograft model. Asian Pac. J. Cancer Prev. 2014, 15, 6219–6225. [Google Scholar] [CrossRef] [Green Version]
- Umar Lule, S.; Xia, W.J.F.R.I. Food phenolics, pros and cons: A review. J. Food Rev. Int. 2005, 21, 367–388. [Google Scholar] [CrossRef]
- Hao, J.; Zhu, H.; Zhang, Z.; Yang, S.; Li, H. Identification of anthocyanins in black rice (Oryza sativa L.) by UPLC/Q-TOF-MS and their in vitro and in vivo antioxidant activities. J. Cereal Sci. 2015, 64, 92–99. [Google Scholar] [CrossRef]
- García-Alonso, M.; Rimbach, G.; Rivas-Gonzalo, J.C.; de Pascual-Teresa, S. Antioxidant and cellular activities of anthocyanins and their corresponding vitisins A studies in platelets, monocytes, and human endothelial cells. J. Agric. Food Chem. 2004, 52, 3378–3384. [Google Scholar] [CrossRef] [PubMed]
- Turturică, M.; Oancea, A.M.; Râpeanu, G.; Bahrim, G. Anthocyanins: Naturally occuring fruit pigments with functional properties. Ann. Univ. Dunarea Jos Galati Fascicle VI-Food Technol. 2015, 39, 9–24. [Google Scholar]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Tahkokorpi, M.; Korteniemi, A.; Taulavuori, E.; Roitto, M.; Laine, K.; Huttunen, S.; Taulavuori, K. Trace amounts of nickel in belowground rhizomes of Vaccinium myrtillus L. decrease anthocyanin concentrations in aerial shoots without water stress. J. Environ. 2010, 69, 338–342. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, A.; Wu, X.; Zhu, Z.; Yang, Z.; Zhu, Y.; Zha, D. Transcriptome analysis revealed expression of genes related to anthocyanin biosynthesis in eggplant (Solanum melongena L.) under high-temperature stress. BMC Plant Biol. 2019, 19, 387. [Google Scholar] [CrossRef] [Green Version]
- Han, S.J.; Ryu, S.N.; Kang, S. A new 2-arylbenzofuran with antioxidant activity from the black colored rice (Oryza sativa L.) bran. Chem. Pharm. Bull. 2004, 52, 1365–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, C.K.; Kimi, L.; Haripriya, S.; Kang, N. Effects of polishing on proximate composition, physico-chemical characteristics, mineral composition and antioxidant properties of pigmented rice. Rice Sci. 2017, 24, 241–252. [Google Scholar] [CrossRef]
- Savitha, Y.; Singh, V. Technology, Status of dietary fiber contents in pigmented and non-pigmented rice varieties before and after parboiling. J. LWT-Food Sci. 2011, 44, 2180–2184. [Google Scholar] [CrossRef]
- Zhang, H.; Shao, Y.; Bao, J.; Beta, T. Phenolic compounds and antioxidant properties of breeding lines between the white and black rice. Food Chem. 2015, 172, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Kou, X.; Fugal, K.; McLaughlin, J. Comparison of HPLC methods for determination of anthocyanins and anthocyanidins in bilberry extracts. J. Agric. Food Chem. 2004, 52, 688–691. [Google Scholar] [CrossRef]
- Goufo, P.; Trindade, H. Rice antioxidants: Phenolic acids, flavonoids, anthocyanins, proanthocyanidins, tocopherols, tocotrienols, γ-oryzanol, and phytic acid. Food Sci. Nutr. 2014, 2, 75–104. [Google Scholar] [CrossRef]
- Xu, T.; Sun, J.; Chang, H.; Zheng, H.; Wang, J.; Liu, H.; Yang, L.; Zhao, H.; Zou, D.T. QTL mapping for anthocyanin and proanthocyanidin content in red rice. Euphytica 2017, 213, 243. [Google Scholar] [CrossRef]
- Jang, S.; Xu, Z. Lipophilic and hydrophilic antioxidants and their antioxidant activities in purple rice bran. J. Agric. Food Chem. 2009, 57, 858–862. [Google Scholar] [CrossRef]
- Lee, J.H. Identification and quantification of anthocyanins from the grains of black rice (Oryza sativa L.) varieties. Food Sci. Biotechnol. 2010, 19, 391–397. [Google Scholar] [CrossRef]
- Shao, Y.; Bao, J. Polyphenols in whole rice grain: Genetic diversity and health benefits. Food. Chem. 2015, 180, 86–97. [Google Scholar] [CrossRef]
- Shao, Y.; Xu, F.; Sun, X.; Bao, J.; Beta, T. Identification and quantification of phenolic acids and anthocyanins as antioxidants in bran, embryo and endosperm of white, red and black rice kernels (Oryza sativa L.). J. Cereal Sci. 2014, 59, 211–218. [Google Scholar] [CrossRef]
- Laokuldilok, T.; Shoemaker, C.F.; Jongkaewwattana, S.; Tulyathan, V. Antioxidants and antioxidant activity of several pigmented rice brans. J. Agric. Food Chem. 2011, 59, 193–199. [Google Scholar] [CrossRef]
- Pedro, A.C.; Granato, D.; Rosso, N. Extraction of anthocyanins and polyphenols from black rice (Oryza sativa L.) by modeling and assessing their reversibility and stability. J. Food Chem. 2016, 191, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Zaupa, M.; Calani, L.; Del Rio, D.; Brighenti, F.; Pellegrini, N. Characterization of total antioxidant capacity and (poly) phenolic compounds of differently pigmented rice varieties and their changes during domestic cooking. Food Chem. 2015, 187, 338–347. [Google Scholar] [CrossRef]
- Terungwa, T.I.; Yuguda, M. The impact of rice production, consumption and importation in Nigeria? The political economy perspectives. Int. J. Sustain. Dev. World Policy 2014, 3, 90–99. [Google Scholar]
- Das, A.B.; Goud, V.; Das, C. Extraction of phenolic compounds and anthocyanin from black and purple rice bran (Oryza sativa L.) using ultrasound: A comparative analysis and phytochemical profiling. Ind. Crops Prod. 2017, 95, 332–341. [Google Scholar] [CrossRef]
- Gunaratne, A.; Wu, K.; Li, D.; Bentota, A.; Corke, H.; Cai, Y.-Z. Antioxidant activity and nutritional quality of traditional red-grained rice varieties containing proanthocyanidins. Food Chem. 2013, 138, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Jiamyangyuen, S.; Nuengchamnong, N.; Ngamdee, P. Bioactivity and chemical components of Thai rice in five stages of grain development. J. Cereal Sci. 2017, 74, 136–144. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89. [Google Scholar] [PubMed]
- Chen, M.-H.; McClung, A.M.; Bergman, C.J. Phenolic content, anthocyanins and antiradical capacity of diverse purple bran rice genotypes as compared to other bran colors. J. Cereal Sci. 2017, 77, 110–119. [Google Scholar] [CrossRef]
- Kim, C.; Kikuchi, S.; Kim, Y.; Park, S.; Yoon, U.; Lee, G.; Choi, J.; Kim, Y.; Park, S. Computational identification of seed-specific transcription factors involved in anthocyanin production in black rice. Bio. Chip J. 2010, 4, 247–255. [Google Scholar] [CrossRef]
- Abdel-Aal, E.-S.M.; Young, J.C.; Rabalski, I. Anthocyanin composition in black, blue, pink, purple, and red cereal grains. J. Agric. Food Chem. 2006, 54, 4696–4704. [Google Scholar] [CrossRef] [PubMed]
- Hiemori, M.; Koh, E.; Mitchell, A.E. Influence of cooking on anthocyanins in black rice (Oryza sativa L. japonica var. SBR). J. Agric. Food Chem. 2009, 57, 1908–1914. [Google Scholar] [CrossRef]
- Bordiga, M.; Gomez-Alonso, S.; Locatelli, M.; Travaglia, F.; Coïsson, J.D.; Hermosin-Gutierrez, I.; Arlorio, M. Phenolics characterization and antioxidant activity of six different pigmented Oryza sativa L. cultivars grown in Piedmont (Italy). Food Res. Int. 2014, 65, 282–290. [Google Scholar] [CrossRef]
- Frank, T.; Reichardt, B.; Shu, Q.; Engel, K.-H. Metabolite profiling of colored rice (Oryza sativa L.) grains. J. Cereal Sci. 2012, 55, 112–119. [Google Scholar] [CrossRef]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: A colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. J. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, C.; Terrier, N.; Torregrosa, L.; Vialet, S.; Fournier-Level, A.; Verries, C.; Souquet, J.-M.; Mazauric, J.-P.; Klein, M.; Cheynier, V. Grapevine MATE-type proteins act as vacuolar H+-dependent acylated anthocyanin transporters. J. Plant Physiol. 2009, 150, 402–415. [Google Scholar] [CrossRef] [Green Version]
- Ling-Peng, D.; Xin-Jiao, D.; Hai-Hu, M. Molecular mechanism for cadmium-induced anthocyanin accumulation in Azolla imbricate. Chemosphere 2012, 87, 319–325. [Google Scholar] [CrossRef]
- Abrahams, S.; Lee, E.; Walker, A.R.; Tanner, G.J.; Larkin, P.J.; Ashton, A.R. The Arabidopsis TDS4 gene encodes leucoanthocyanidin dioxygenase (LDOX) and is essential for proanthocyanidin synthesis and vacuole development. Plant J. 2003, 35, 624–636. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Yamaji, N.; Mitani-Ueno, N.; Kashino, M.; Ma, J. A nodelocalized transporter Os ZIP 3 is responsible for the preferential distribution of Zn to developing tissues in rice. Plant J. 2015, 84, 374–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier-Level, A.; Hugueney, P.; Verriès, C.; This, P.; Ageorges, A. Genetic mechanisms underlying the methylation level of anthocyanins in grape (Vitis vinifera L.). BMC Plant Biol. 2011, 11, 179. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Wu, J.; Ji, K.-X.; Zeng, Q.-Y.; Bhuiya, M.-W.; Su, S.; Shu, Q.-Y.; Ren, H.-X.; Liu, Z.-A.; Wang, L.-S. Methylation mediated by an anthocyanin, O-methyltransferase, is involved in purple flower coloration in Paeonia. J. Exp. Bot. 2015, 66, 6563–6577. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Butelli, E.; Martin, C. Engineering anthocyanin biosynthesis in plants. Curr. Opin. Plant Biol. 2014, 19, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Holton, T.A.; Cornish, E.C. Genetics and biochemistry of anthocyanin biosynthesis. J. Plant Cell. 1995, 7, 1071. [Google Scholar] [CrossRef] [PubMed]
- Tanner, G.J.; Francki, K.T.; Abrahams, S.; Watson, J.M.; Larkin, P.J.; Ashton, A.R. Proanthocyanidin biosynthesis in plants purification of legume leucoanthocyanidin reductase and molecular cloning of its cDNA. J. Biol. Chem. 2003, 278, 31647–31656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devic, M.; Guilleminot, J.; Debeaujon, I.; Bechtold, N.; Bensaude, E.; Koornneef, M.; Pelletier, G.; Delseny, M. The BANYULS gene encodes a DFR-like protein and is a marker of early seed coat development. Plant J. 1999, 19, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S. Transport of flavonoids: From cytosolic synthesis to vacuolar accumulation. In The Science of Flavonoids; Springer: Berlin/Heidelberg, Germany, 2006; pp. 123–146. [Google Scholar] [CrossRef]
- Reimold, U.; Kröger, M.; Kreuzaler, F.; Hahlbrock, K. Coding and 3′ non-coding nucleotide sequence of chalcone synthase mRNA and assignment of amino acid sequence of the enzyme. EMBO J. 1983, 2, 1801–1805. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Yu, S.; Zeng, D.; Liu, H.; Wang, H.; Yang, Z.; Xie, X.; Shen, R.; Tan, J.; Li, H. Development of “purple endosperm rice” by engineering anthocyanin biosynthesis in the endosperm with a high-efficiency transgene stacking system. Mol. Plant 2017, 10, 918–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.; Dabi, T.; Beeche, A.; Yamamoto, R.; Lawton, M.A.; Lamb, C. Cloning and properties of a rice gene encoding phenylalanine ammonia-lyase. J. Plant Mol. Biol. 1995, 29, 535–550. [Google Scholar] [CrossRef]
- Yang, G.-H.; Li, B.; Gao, J.-W.; Liu, J.-Z.; Zhao, X.-Q.; Zheng, Q.; Tong, Y.-P.; Li, Z.-S. Cloning and expression of two chalcone synthase and a flavonoid 3′5′-Hydroxylase 3′-end cDNAs from developing seeds of blue-grained wheat involved in anthocyanin biosynthetic pathway. J. Acta. Bot. Sin. 2004, 46, 588–594. [Google Scholar]
- Gui, J.; Shen, J.; Li, L. Functional characterization of evolutionarily divergent 4-coumarate: Coenzyme A ligases in rice. J. Plant Physiol. 2011, 157, 574–586. [Google Scholar] [CrossRef] [Green Version]
- Shih, C.H.; Chu, H.; Tang, L.K.; Sakamoto, W.; Maekawa, M.; Chu, I.K.; Wang, M.; Lo, C. Functional characterization of key structural genes in rice flavonoid biosynthesis. J. Planta 2008, 228, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.-Y.; Foley, M.E.; Horvath, D.P.; Anderson, J.V.; Feng, J.; Zhang, L.; Mowry, C.R.; Ye, H.; Suttle, J.C.; Kadowaki, K.; et al. Association between seed dormancy and pericarp color is controlled by a pleiotropic gene that regulates abscisic acid and flavonoid synthesis in weedy red rice. J. Genet. 2011, 189, 1515–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.H.; Kim, B.G.; Kim, J.H.; Kim, H.; Lim, C.E.; Lim, J.; Lee, C.; Lim, Y.; Ahn, J.-H. Four glucosyltransferases from rice: cDNA cloning, expression, and characterization. J. Plant Physiol. 2008, 165, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-G.; Lee, Y.; Hur, H.-G.; Lim, Y.; Ahn, J.-H. Flavonoid 3′-O-methyltransferase from rice: cDNA cloning, characterization and functional expression. J. Phytochem. 2006, 67, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.G.; Kim, J.H.; Min, S.Y.; Shin, K.-H.; Kim, J.H.; Kim, H.Y.; Ryu, S.N. Anthocyanin content in rice is related to expression levels of anthocyanin biosynthetic genes. J. Plant Biol. 2007, 50, 156–160. [Google Scholar] [CrossRef]
- Lamberts, L.; De Bie, E.; Vandeputte, G.E.; Veraverbeke, W.S.; Derycke, V.; De Man, W.; Delcour, J.A. Effect of milling on colour and nutritional properties of rice. Food Chem. 2007, 100, 1496–1503. [Google Scholar] [CrossRef]
- Blank, F. The anthocyanin pigments of plants. Bot. Rev. 1947, 13, 241–317. [Google Scholar] [CrossRef]
- Finocchiaro, F.; Ferrari, B.; Gianinetti, A. A study of biodiversity of flavonoid content in the rice caryopsis evidencing simultaneous accumulation of anthocyanins and proanthocyanidins in a black-grained genotype. J. Cereal Sci. 2010, 51, 28–34. [Google Scholar] [CrossRef]
- Grotewold, E.; Davies, K. Trafficking and sequestration of anthocyanins. Nat. Prod. Commun. 2008, 3. [Google Scholar] [CrossRef] [Green Version]
- Gomez, C.; Conejero, G.; Torregrosa, L.; Cheynier, V.; Terrier, N.; Ageorges, A. In vivo grapevine anthocyanin transport involves vesicle-mediated trafficking and the contribution of anthoMATE transporters and GST. Plant J. 2011, 67, 960–970. [Google Scholar] [CrossRef]
- Poustka, F.; Irani, N.G.; Feller, A.; Lu, Y.; Pourcel, L.; Frame, K.; Grotewold, E. A trafficking pathway for anthocyanins overlaps with the endoplasmic reticulum-to-vacuole protein-sorting route in Arabidopsis and contributes to the formation of vacuolar inclusions. Plant. Physiol. 2007, 145, 1323–1335. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; An, W.; Wang, K.; Jiang, T.; Ren, Q.; Liang, W.; Wang, H. Expression profile analysis of MATE gene family in rice. Biol. Plant. 2019, 63, 556–564. [Google Scholar] [CrossRef] [Green Version]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef] [PubMed]
- Mueller, L.A.; Goodman, C.D.; Silady, R.A.; Walbot, V. AN9, a petunia glutathione S-transferase required for anthocyanin sequestration, is a flavonoid-binding protein. J. Plant. Physiol. 2000, 123, 1561–1570. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Li, H.; Huang, J.-R. Arabidopsis TT19 functions as a carrier to transport anthocyanin from the cytosol to tonoplasts. Mol. Plant 2012, 5, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Jasinski, M.; Ducos, E.; Martinoia, E.; Boutry, M. The ATP-Binding Cassette Transporters: Structure, Function, and Gene Family Comparison between Rice and Arabidopsis. J. Plant Physiol. 2003, 131, 1169–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.-L.; Xie, X.-R.; Zhang, J.; Xiang, G.; Li, Y.; Wu, H.-B. In Silico Analysis of a MRP Transporter Gene Reveals Its Possible Role in Anthocyanins or Flavonoids Transport in Oryza sativa. Am. J. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Goodman, C.D.; Casati, P.; Walbot, V. A multidrug resistance–associated protein involved in anthocyanin transport in Zea mays. Plant Cell. 2004, 16, 1812–1826. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Zhang, Q.; Zhu, Q.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.; Li, H.; Lin, Y. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. J. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef]
- Marrs, K.A.; Alfenito, M.R.; Lloyd, A.M.; Walbot, V. A glutathione S-transferase involved in vacuolar transfer encoded by the maize gene Bronze-2. Nature 1995, 375, 397–400. [Google Scholar] [CrossRef]
- Das, N.; Bhattacharya, S.; Bhattacharyya, S.; Maiti, M.K. Expression of rice MATE family transporter OsMATE2 modulates arsenic accumulation in tobacco and rice. Plant Mol. Biol. 2018, 98, 101–120. [Google Scholar] [CrossRef]
- Heim, M.A.; Jakoby, M.; Werber, M.; Martin, C.; Weisshaar, B.; Bailey, P.C. The basic helix–loop–helix transcription factor family in plants: A genome-wide study of protein structure and functional diversity. Mol. Biol. Evol. 2003, 20, 735–747. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, T.; Maeda, H.; Oguchi, T.; Yamaguchi, T.; Tanabe, N.; Ebana, K.; Yano, M.; Ebitani, T.; Izawa, T. The birth of a black rice gene and its local spread by introgression. Plant Cell. 2015, 27, 2401–2414. [Google Scholar] [CrossRef] [Green Version]
- Petroni, K.; Tonelli, C. Recent advances on the regulation of anthocyanin synthesis in reproductive organs. Plant Sci. 2011, 181, 219–229. [Google Scholar] [CrossRef]
- Schijlen, E.G.; De Vos, C.R.; van Tunen, A.J.; Bovy, A.G. Modification of flavonoid biosynthesis in crop plants. Phytochemistry 2004, 65, 2631–2648. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wu, H.; Zhu, H.; Huang, C.; Liu, C.; Chang, Y.; Kong, Z.; Zhou, Z.; Wang, G.; Lin, Y.; et al. Determining factors, regulation system, and domestication of anthocyanin biosynthesis in rice leaves. New Phytol. 2019, 223, 705–721. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, W.; Ohmori, T.; Kageyama, K.; Miyazaki, C.; Saito, A.; Murata, M.; Noda, K.; Maekawa, M. The Purple leaf (Pl) Locus of Rice: The Plw Allele has a Complex Organization and Includes Two Genes Encoding Basic Helix-Loop-Helix Proteins Involved in Anthocyanin Biosynthesis. Plant. Cell Physiol. 2001, 42, 982–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, M.T.; Thomson, M.J.; Pfeil, B.E.; McCouch, S. Caught red-handed: Rc encodes a basic helix-loop-helix protein conditioning red pericarp in rice. Plant Cell. 2006, 18, 283–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitoh, K.; Onishi, K.; Mikami, I.; Thidar, K.; Sano, Y. Allelic diversification at the C (OsC1) locus of wild and cultivated rice: Nucleotide changes associated with phenotypes. Genetics 2004, 168, 997–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, H.; Yamaguchi, T.; Omoteno, M.; Takarada, T.; Fujita, K.; Murata, K.; Iyama, Y.; Kojima, Y.; Morikawa, M.; Ozaki, H.J. Genetic dissection of black grain rice by the development of a near isogenic line. Breed. Sci. 2014, 64, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.-H.; Lee, Y.-J.; Byeon, E.-J.; Kang, B.-C.; Kyeoung, D.-S.; Kim, C.-K. Whole-genome resequencing and transcriptomic analysis of genes regulating anthocyanin biosynthesis in black rice plants. 3 Biotech 2018, 8, 115. [Google Scholar] [CrossRef] [Green Version]
- Bai, B.; Lu, N.; Li, Y.; Guo, S.; Yin, H.; He, Y.; Sun, W.; Li, W.; Xie, X. OsBBX14 promotes photomorphogenesis in rice by activating OsHY5L1 expression under blue light conditions. Plant Sci. 2019, 284, 192–202. [Google Scholar] [CrossRef]
- Ham, T.-H.; Kwon, S.W.; Ryu, S.-N.; Koh, H.-J. Biotechnology, Correlation analysis between grain color and cyanidin-3-glucoside content of rice grain in segregate population. Plant Breed. 2015, 3, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; Lee, K.E.; Lee, E.S.; Matin, M.N.; Lee, D.S.; Yun, J.S.; Kim, J.B.; Kang, S.G. The genetic constitutions of complementary genes Pp and Pb determine the purple color variation in pericarps with cyanidin-3-O-glucoside depositions in black rice. J. Plant Biol. 2013, 56, 24–31. [Google Scholar] [CrossRef]
- Hu, J.; Anderson, B.; Wessler, S.R. Isolation and characterization of rice R genes: Evidence for distinct evolutionary paths in rice and maize. Genetics 1996, 142, 1021–1031. [Google Scholar] [CrossRef]
- Wang, C.; Shu, Q. Fine mapping and candidate gene analysis of purple pericarp gene Pb in rice (Oryza sativa L.). Chin. Sci. Bull. 2007, 52, 3097–3104. [Google Scholar] [CrossRef]
- Kim, C.K.; Seol, Y.J.; Shin, Y.; Lim, H.M.; Lee, G.S.; Kim, A.R.; Lee, T.H.; Lee, J.H.; Park, D.S.; Yoo, S.; et al. Whole-genome resequencing and transcriptomic analysis to identify genes involved in leaf-color diversity in ornamental rice plants. PLoS ONE 2015, 10, e0124071. [Google Scholar] [CrossRef] [Green Version]
- Chin, H.-S.; Wu, Y.-P.; Hour, A.-L.; Hong, C.-Y.; Lin, Y.-R. Genetic and Evolutionary Analysis of Purple Leaf Sheath in Rice. Rice 2016, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Sakulsingharoj, C.; Inta, P.; Sukkasem, R.; Pongjaroenkit, S.; Chowpongpang, S.; Sangtong, V. Cloning and characterization of OSB1 gene controlling anthocyanin biosynthesis from Thai black rice. Genom. Genet. 2016, 9, 7–18. [Google Scholar] [CrossRef]
- Mbanjo, E.G.N.; Kretzschmar, T.; Jones, H.; Ereful, N.; Blanchard, C.; Boyd, L.A.; Sreenivasulu, N. The Genetic Basis and Nutritional Benefits of Pigmented Rice Grain. Front. Genet. 2020, 11, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rachasima, L.N.; Sukkasem, R.; Pongjaroenkit, S.; Sangtong, V.; Chowpongpang, S.; Sakulsingharoj, C. Expression analysis and nucleotide variation of OsC1 gene associated with anthocyanin pigmentation in Thai rice cultivars. Genom. Genet. 2017, 10, 46–53. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, Z.; Chen, C.; Wu, W.; Ren, N.; Jiang, C.; Yu, J.; Zhao, Y.; Zheng, X.; Yang, Q. The C–S–A gene system regulates hull pigmentation and reveals evolution of anthocyanin biosynthesis pathway in rice. J. Exp. Bot. 2018, 69, 1485–1498. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.; Visser, R.G.; Bovy, A. Anthocyanin biosynthesis and degradation mechanisms in Solanaceous vegetables: A review. J. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L.J. New insights into the regulation of anthocyanin biosynthesis in fruits. J. Trends. Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Wang, X.; Gao, J.; Guo, Y.; Huang, Z.; Du, Y. The tomato Hoffman’s anthocyaninless gene encodes a bHLH transcription factor involved in anthocyanin biosynthesis that is developmentally regulated and induced by low temperatures. PLoS ONE 2016, 11, e0151067. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Sun, X.; Wang, W.; Ding, H.; Liu, W.; Li, G.; Jiang, M.; Zhu, C.; Yao, F. Fine mapping of Pa-6 gene for purple apiculus in rice. J. Plant. Biol. 2012, 55, 218–225. [Google Scholar] [CrossRef]
- Fan, F.-J.; Fan, Y.-Y.; Du, J.-h.; Zhuang, J.-Y. Fine mapping of C (chromogen for anthocyanin) gene in rice. Rice Sci. 2008, 15, 1–6. [Google Scholar] [CrossRef]
- Zhao, S.; Wang, C.; Ma, J.; Wang, S.; Tian, P.; Wang, J.; Cheng, Z.; Zhang, X.; Guo, X.; Lei, C. Map-based cloning and functional analysis of the chromogen gene C in rice (Oryza sativa L.). J. Plant Biol. 2016, 59, 496–505. [Google Scholar] [CrossRef]
- Chen, C.; Wu, W.; Sun, X.; Li, B.; Hu, G.; Zhang, Q.; Li, J.; Zhang, H.; Li, Z. Fine-mapping and candidate gene analysis of BLACK HULL1 in rice (Oryza sativa L.). J. Plant Omics. 2014, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Dai, G.; Zhou, W.; Liang, H.; Huang, J.; Qing, D.; Chen, W.; Wu, H.; Yang, X.; Li, D.J.; et al. Mapping and identifying a candidate gene Plr4, a recessive gene regulating purple leaf in rice, by using bulked segregant and transcriptome analysis with next-generation sequencing. Int. J. Mol. Sci. 2019, 20, 4335. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Deng, W.; Li, F.; Zhou, J.; Li, J.; Xu, P.; Deng, X.; Hu, F.; Wang, L.; Chen, S.; et al. A genetic study on the purple stigma genes and their locations in Oryza longistaminata. J. Yunnan Univ.-Nat. Sci. Ed. 2010, 32, 103–107. [Google Scholar]
- Gao, D.; He, B.; Zhou, Y.; Sun, L. Genetic and molecular analysis of a purple sheath somaclonal mutant in japonica rice. Plant Cell. Rep. 2011, 30, 901–911. [Google Scholar] [CrossRef]
- Schell, J.S. Transgenic plants as tools to study the molecular organization of plant genes. J. Sci. 1987, 237, 1176–1183. [Google Scholar] [CrossRef]
- Shin, Y.M.; Park, H.J.; Yim, S.D.; Baek, N.I.; Lee, C.H.; An, G.; Woo, Y.M. Transgenic rice lines expressing maize C1 and R-S regulatory genes produce various flavonoids in the endosperm. Plant Biotechnol. J. 2006, 4, 303–315. [Google Scholar] [CrossRef]
- Kawahigashi, H.; Hirose, S.; Iwai, T.; Ohashi, Y.; Sakamoto, W.; Maekawa, M.; Ohkawa, Y. Chemically induced expression of rice OSB2 under the control of the OsPR1.1 promoter confers increased anthocyanin accumulation in transgenic rice. J. Agric. Food Chem. 2007, 55, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Sakulsingharoj, C.; Inta, P.; Sukkasem, R.; Pongjaroenkit, S.; Chowpongpang, S.; Sangtong, V. Overexpression of OSB2 gene in transgenic rice up-regulated expression of structural genes in anthocyanin biosynthesis pathway. Genomics 2014, 7, 173–182. [Google Scholar] [CrossRef]
- Van Long, H.; Yabe, M. The Impact of Environmental Factors on the Productivity and Efficiency of Rice Production: A Study in Vietnam’s Red River Delta. Eur. J. Soc. Sci. 2011, 26, 218–230. [Google Scholar]
- Singh, T.; Pun, K.; Bhagat, K.; Lal, B.; Satapathy, B.; Sadawarti, M.; Katara, J.; Gautam, S.L. Abiotic stress in rice: Mechanism of adaptation. In Challenges and Prospective of Plant Abiotic Stress; 4436/7; Today & Tomorrow’s Printers and Publishers: New Delhi, India, 2015; Volume 1, pp. 259–360. [Google Scholar]
- Mishra, A.K.; Mottaleb, K.A.; Khanal, A.R.; Mohanty, S. Abiotic stress and its impact on production efficiency: The case of rice farming in Bangladesh. J. Agric. Ecosyst. Environ. 2015, 199, 146–153. [Google Scholar] [CrossRef]
- Li, X.; Wu, L.; Geng, X.; Xia, X.; Wang, X.; Xu, Z.; Xu, Q. Deciphering the environmental impacts on rice quality for different rice cultivated areas. Rice Sci. 2018, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anami, B.S.; Malvade, N.N.; Palaiah, S. Classification of yield affecting biotic and abiotic paddy crop stresses using field images. J. Inf. Proc. Agric. 2019. [Google Scholar] [CrossRef]
- Woodward, G. Anthocyanin Stability, Metabolic Conjugation and in vitro Modulation of Endothelial Superoxide Production. Ph.D. Thesis, University of East Anglia, Norwich, UK, 2010. [Google Scholar]
- Bakhshayeshi, M.; Khayami, M.; Heidari, R.; Jamei, R. The effects of light, storage temperature, pH and variety on stability of anthocyanin pigments in four Malus varieties. Pak. J. Biol. Sci. 2006, 9, 428–433. [Google Scholar] [CrossRef] [Green Version]
- Bridle, P.; Timberlake, C. Anthocyanins as natural food colours—selected aspects. J. Food Chem. 1997, 58, 103–109. [Google Scholar] [CrossRef]
- Lv, X.L.; Sun, H.L.; Ji, Z.Y. Acylation of anthocyanins from black rice and their stability properties. Adv. Mater. Res. 2011, 204–210, 750–754. [Google Scholar] [CrossRef]
- Shaked-Sachray, L.; Weiss, D.; Reuveni, M.; Nissim-Levi, A.; Oren-Shamir, M. Increased anthocyanin accumulation in aster flowers at elevated temperatures due to magnesium treatment. J. Physiol. Plant 2002, 114, 559–565. [Google Scholar] [CrossRef]
- Jiang, M.; Liu, Y.; Ren, L.; Lian, H.; Chen, H. Molecular cloning and characterization of anthocyanin biosynthesis genes in eggplant (Solanum melongena L.). Acta Physiol. Plant 2016, 38, 163. [Google Scholar] [CrossRef]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot berries. Am. J. Enol. Vitic. 2002, 53, 171–182. [Google Scholar]
- Movahed, N.; Pastore, C.; Cellini, A.; Allegro, G.; Valentini, G.; Zenoni, S.; Cavallini, E.; D’Incà, E.; Tornielli, G.B.; Filippetti, I. The grapevine VviPrx31 peroxidase as a candidate gene involved in anthocyanin degradation in ripening berries under high temperature. J. Plant Res. 2016, 129, 513–526. [Google Scholar] [CrossRef]
- Niu, J.; Zhang, G.; Zhang, W.; Goltsev, V.; Sun, S.; Wang, J.; Li, P.; Ma, F. Anthocyanin concentration depends on the counterbalance between its synthesis and degradation in plum fruit at high temperature. Sci. Rep. 2017, 7, 7684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowan, D.D.; Cao, M.; Lin-Wang, K.; Cooney, J.M.; Jensen, D.J.; Austin, P.T.; Hunt, M.B.; Norling, C.; Hellens, R.P.; Schaffer, R. Environmental regulation of leaf colour in red 35S: PAP1 Arabidopsis thaliana. New Phytol. 2009, 182, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, W.; Zhang, P.; Pan, Q.; Zhan, J.; Huang, W. Gene transcript accumulation, tissue and subcellular localization of anthocyanidin synthase (ANS) in developing grape berries. Plant Sci. 2010, 179, 103–113. [Google Scholar] [CrossRef]
- Zaidi, S.H.R.; Zakari, S.A.; Zhao, Q.; Khan, A.R.; Shah, J.M.; Cheng, F. Anthocyanin Accumulation in Black Kernel Mutant Rice and its Contribution to ROS Detoxification in Response to High Temperature at the Filling Stage. Antiox 2019, 8, 510. [Google Scholar] [CrossRef] [Green Version]
- Kushwaha, U.; Research, A. Black rice anthocyanin content increases with increase in altitude of its plantation. Adv. Plants 2016, 5, 00170. [Google Scholar] [CrossRef] [Green Version]
- Takahama, U. Oxidation of vacuolar and apoplastic phenolic substrates by peroxidase: Physiological significance of the oxidation reactions. Phytochem. Rev. 2004, 3, 207–219. [Google Scholar] [CrossRef]
- Huang, D.; Yuan, Y.; Tang, Z.; Huang, Y.; Kang, C.; Deng, X.; Xu, Q. Retrotransposon Promoter of Ruby1 Controls Both Light- and Cold-Induced Accumulation of Anthocyanins in Blood Orange. Plant Cell Environ. 2019, 42, 3092–3104. [Google Scholar] [CrossRef]
- Chan, L.; Koay, S.; Boey, P.; Bhatt, A. Effects of abiotic stress on biomass and anthocyanin production in cell cultures of Melastoma malabathricum. J. Biol. Res. 2010, 43, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Azuma, A. Genetic and environmental impacts on the biosynthesis of anthocyanins in grapes. Hortic. J. 2018, OKD-IR02. [Google Scholar] [CrossRef] [Green Version]
- Kashif, M.H.; Wei, F.; Tang, D.; Tang, M.; Luo, D.; Hai, L.; Li, R.; Chen, P. iTRAQ-based comparative proteomic response analysis reveals regulatory pathways and divergent protein targets associated with salt-stress tolerance in kenaf (Hibiscus cannabinus L.). Ind. Crops 2020, 153, 112566. [Google Scholar] [CrossRef]
- Kashif, M.H.; Tang, D.; Li, Z.; Wei, F.; Liang, Z.; Chen, P. Comparative Cytological and Gene Expression Analysis Reveals Potential Metabolic Pathways and Target Genes Responsive to Salt Stress in Kenaf (Hibiscus cannabinus L.). J. Plant Growth Reg. 2020, 1245–1260. [Google Scholar] [CrossRef]
- Gould, K. Nature’s Swiss army knife: The diverse protective roles of anthocyanins in leaves. Biomed. Res. Int. 2004, 5, 314–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chunthaburee, S.; Sakuanrungsirikul, S.; Wongwarat, T.; Sanitchon, J.; Pattanagul, W.; Theerakulpisut, P. Changes in anthocyanin content and expression of anthocyanin synthesis genes in seedlings of black glutinous rice in response to salt stress. Asian J. Plant Sci. 2016, 15, 56–65. [Google Scholar] [CrossRef]
- Sinilal, B.; Ovadia, R.; Nissim-Levi, A.; Perl, A.; Carmeli-Weissberg, M.; Oren-Shamir, M. Increased accumulation and decreased catabolism of anthocyanins in red grape cell suspension culture following magnesium treatment. Planta 2011, 234, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Tisarum, R.; Theerawitaya, C.; Samphumphuang, T.; Cha-um, S.; Science, S. Regulation of anthocyanin accumulation in rice (Oryza sativa L. subsp. indica) using MgSO4 spraying and low temperature. Arch. Agron. Soil Sci. 2018, 64, 1663–1677. [Google Scholar] [CrossRef]
- Yamuangmorn, S.; Dell, B.; Rerkasem, B.; Prom-u-thai, C. Agriculture, Applying nitrogen fertilizer increased anthocyanin in vegetative shoots but not in grain of purple rice genotypes. J. Sci. Food Agric. 2018, 98, 4527–4532. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, J.; Waite, R.; Searchinger, T.; Hanson, C. How to Sustainably Feed 10 billion People by 2050, in 21 Charts. World Resour. Inst. 2018. Available online: https://www.wri.org/blog/2018/12/how-sustainably-feed-10-billion-people-2050-21-charts (accessed on 10 April 2020).





| Groups | Protein | Gene Name | Locus ID | Reference |
|---|---|---|---|---|
| Phenylalanine Pathway Structural Genes | Phenylalanine Ammonia-Lyase (PAL) | OsPAL | Os02g0627100 | [62] |
| Cinnamate 4-Hydroxylase (C4H) | OsC4H | Os05g0320700 | [61,63] | |
| 4-Coumaroyl CoA Ligase (4CL) | Os4CL | Os02g0177600 | [64] | |
| Anthocyanin Pathway Structural Genes | Chalcone Synthase (CHS) | OsCHS | Os11g0530600 | [65] |
| Chalcone Isomerase (CHI) | OsCHI | Os03g0819600 | [65] | |
| Flavanone 3-Hydroxylase (F3H) | OsF3H | Os04g0662600 | [65] | |
| Flavanone 3′-Hydroxylase (F3′H) | OsF3′H | Os10g0320100 | [65] | |
| flavonoid 3′-5′ hydroxylase (F 3′5′H) | OsF3′5′H | Os03g0367200 | [61] | |
| dihydroflavonol 4-reductase (DFR) | OsDFR | Os01g0633500 | [65] | |
| Anthocyanidin synthase (ANS) | OsANS | Os01g0372500 | [65] | |
| Leucoanthocyanidin reductase (LAR) | OsLAR | Os03g0259400b | [64] | |
| anthocyanidin reductase (ANR) | OsANR | Os04g0630800 | [66] | |
| Decorating Genes | Anthocyanin 3-O-glycosyltransferases | Os3GT | Os06g0192100 | [67] |
| Anthocyanin 3′-O-Methyltransferase | Os3′MT | Os08g0157500 | [68] |
| Protein | Gene Name | Locus ID | Description | Reference |
|---|---|---|---|---|
| Glutathione-S-transferase U 34 | OsGSTU34 | Os10g0395400 | Thioredoxin fold domain-containing protein (characterized) | [83] |
| Multidrug resistance-associated proteins 15 | OsMRP15 | Os06g0158900 | Similar to Multidrug-resistance associated protein 3 (characterized) | [83] |
| Multidrug and toxic compound extrusion 7 | OsMATE7 | Os02g0821600 | Similar to mate efflux family protein (uncharacterized) | [76] |
| Multidrug and toxic compound extrusion 34 | OsMATE34 | Os08g0562800 | Similar to Transparent testa 12 protein (uncharacterized) | [76] |
| Multidrug and toxic compound extrusion 33 | OsMATE33 | Os08g0550200 | Multi antimicrobial extrusion protein MatE family protein (uncharacterized) | [76] |
| Multidrug and toxic compound extrusion 3 | OsMATE3 | Os01g0766000 | Multi antimicrobial extrusion protein MatE family protein (uncharacterized) | [76] |
| Multidrug and toxic compound extrusion 39 | OsMATE39 | Os10g0195000 | Multi antimicrobial extrusion protein MatE family protein (uncharacterized) | [76] |
| Multidrug and toxic compound extrusion 16 | OsMATE16 | Os03g0626700 | Multi antimicrobial extrusion protein MatE family protein (uncharacterized) | [76] |
| Protein | Gene | Locus ID | Reference |
|---|---|---|---|
| bHLH TF | OSB1 | Os04g0557800 | [92] |
| OSB2 | Os04g0557500 | [92] | |
| Kala4 | Os04g0557500 | [87] | |
| OSRc | Os07g0211500 | [93] | |
| R2R3-MYB TF | OsC1 | Os06g0205100 | [94] |
| OsMYB3 | Kala 3 | Os03t0410000 | [95] |
| WD40 repeat | OsWD40 | Os02g0682500 | [61] |
| Phenotype | Name | Location | Fine Mapping | Reference |
|---|---|---|---|---|
| Colored apiculus | C-gene | RM19552-RM19565 or RM 111-RM253 | C Locus was found to be about 59.3 kb between the SSR markers RM111 and RM 253 with a genetic distance of 0.7 and 0.4, respectively. | [111,112] |
| Purple apiculus | Pa-6 | RM19556—RM19561 with 0.2–0.3 cM respectively | Pa-6 is located 41.7 kb between L02 and RM 19561 containing 11 ORFs of which ORF8 is associated with this trait. | [111] |
| Red apiculus | OSC | An interval of 70.8 kb bounded Dcap13-RM 19561 | OSC is located to an interval of 70.8 kb bounded by Dcap13-RM 19561 and contains 10 ORFs of which ORF8 consists of two introns and three exons. It contains a 10-bp deletion in the third exon, causing a frame-shift mutation and loss of function of the encoded protein. | [113] |
| Black hull | Bh1 | RM6629 and SNP marker SNP6-1 | Bh1 was fine- mapped on the long arm of Chr4 in an interval of 24.2 kb between RM 6629 and SNP6-1. | [114] |
| Purple pericarp | Pb | An interval of 25 kb of RID3 and RID4 | Pb was first mapped downstream of SSR RM3820 on Chr4 and between RID3 and RID4 after saturation with indel and CAP markers | [100] |
| Purple leaf | Plr4 | Recessive gene, Two putative candidates Os04g0577800 and Os04g0616400 | The purple leaf gene was located on Chr4 to about 27.9–31.1 Mb. | [115] |
| Purple stigma | Ps-4(t) | RM253, RM111 and RM6917 | The Ps-4(t) gene was mapped on Chr 6 at 2.5 cM, 0 cM and 4.4 cM, respectively, from RM253, RM111 and RM6917 | [116] |
| Purple leaf sheath | Purple leaf sheath | SSR markers RPM8 and RPM11 | Purple leaf sheath was first mapped to the short arm of Chr6 between RPM5 and RM402 with a genetic distance 1.1 and 10.3 cM, respectively, and then narrowed to an interval of 153 kb between RPM8-RPM11 | [117] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mackon, E.; Jeazet Dongho Epse Mackon, G.C.; Ma, Y.; Haneef Kashif, M.; Ali, N.; Usman, B.; Liu, P. Recent Insights into Anthocyanin Pigmentation, Synthesis, Trafficking, and Regulatory Mechanisms in Rice (Oryza sativa L.) Caryopsis. Biomolecules 2021, 11, 394. https://doi.org/10.3390/biom11030394
Mackon E, Jeazet Dongho Epse Mackon GC, Ma Y, Haneef Kashif M, Ali N, Usman B, Liu P. Recent Insights into Anthocyanin Pigmentation, Synthesis, Trafficking, and Regulatory Mechanisms in Rice (Oryza sativa L.) Caryopsis. Biomolecules. 2021; 11(3):394. https://doi.org/10.3390/biom11030394
Chicago/Turabian StyleMackon, Enerand, Guibeline Charlie Jeazet Dongho Epse Mackon, Yafei Ma, Muhammad Haneef Kashif, Niyaz Ali, Babar Usman, and Piqing Liu. 2021. "Recent Insights into Anthocyanin Pigmentation, Synthesis, Trafficking, and Regulatory Mechanisms in Rice (Oryza sativa L.) Caryopsis" Biomolecules 11, no. 3: 394. https://doi.org/10.3390/biom11030394
APA StyleMackon, E., Jeazet Dongho Epse Mackon, G. C., Ma, Y., Haneef Kashif, M., Ali, N., Usman, B., & Liu, P. (2021). Recent Insights into Anthocyanin Pigmentation, Synthesis, Trafficking, and Regulatory Mechanisms in Rice (Oryza sativa L.) Caryopsis. Biomolecules, 11(3), 394. https://doi.org/10.3390/biom11030394