
Interferon Regulatory Factor 5 (IRF5) Gene Haplotypes Are Associated with Premature Coronary Artery Disease. Association of the IRF5 Polymorphisms with Cardiometabolic Parameters. The Genetics of Atherosclerotic Disease (GEA) Mexican Study

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Subjects
2.3. Genetic Analysis
2.4. Statistical Analysis
3. Results
3.1. Demographic, Clinical, Biochemical, and Lifestyle Characteristics
3.2. Association of IRF5 Polymorphisms with pCAD
3.3. Distribution of IRF5 Haplotypes in pCAD Patients and Healthy Controls
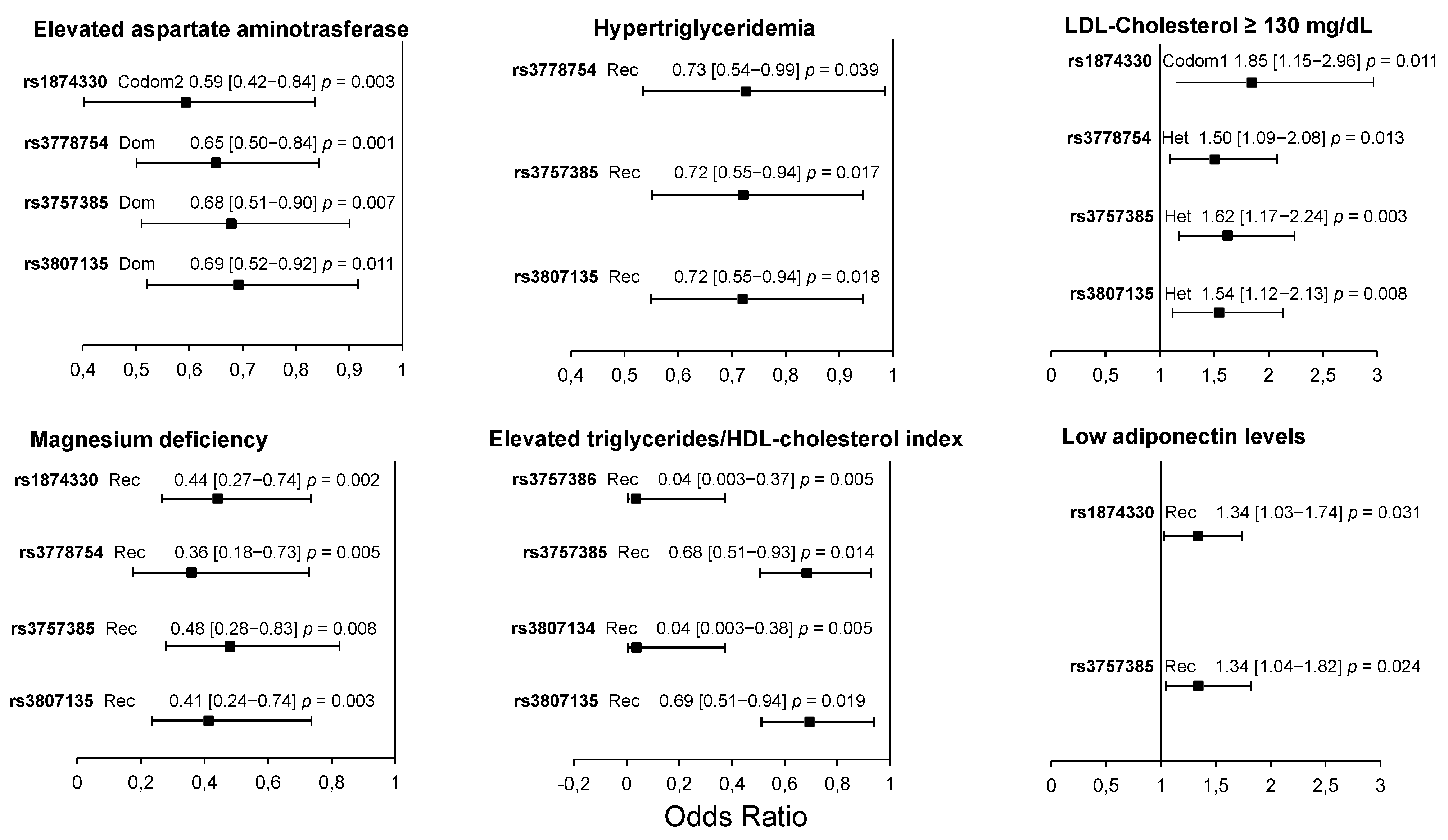
3.4. Association of IRF5 Polymorphisms with Cardiometabolic Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Globalburdenofcoronaryheartdisease. Geneva: World Health Organization. 2004. Available online: http://www.who.int/cardiovascular_diseases/en/cvd_atlas_13_coronaryHD.pdf (accessed on 10 January 2007).
- Libby, P.; Buring, J.; Badimon, L.; Hansson, G.; Deanfield, J.; Bittencourt, M.; Tokgözoğlu, L.; Lewis, E. Atherosclerosis. Nat. Rev. Dis. Primers 2019, 5, 56. [Google Scholar] [CrossRef]
- Gencer, S.; Evans, B.; van der Vorst, E.; Döring, Y.; Weber, C. Inflammatory Chemokines in Atherosclerosis. Cells 2021, 10, 226. [Google Scholar] [CrossRef] [PubMed]
- Takaoka, A.; Yanai, H.; Kondo, S.; Duncan, G.; Negishi, H.; Mizutani, T.; Kano, S.; Honda, K.; Ohba, Y.; Mak, T.; et al. Integral role of IRF-5 in the gene induction programme activated by Toll-like receptors. Nature 2005, 434, 243–249. [Google Scholar] [CrossRef]
- Krausgruber, T.; Blazek, K.; Smallie, T.; Alzabin, S.; Lockstone, H.; Sahgal, N.; Hussell, T.; Feldmann, M.; Udalova, I. IRF5 promotes inflammatory macrophage polarization and TH1-TH17 responses. Nat. Immunol. 2011, 13, 231–238. [Google Scholar] [CrossRef]
- Hedl, M.; Yan, J.; Abraham, C. IRF5 and IRF5 Disease-Risk Variants Increase Glycolysis and Human M1 Macrophage Polarization by Regulating Proximal Signaling and Akt2 Activation. Cell Rep. 2016, 16, 2442–2455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Biswas, S.; Chittezhath, M.; Shalova, I.; Lim, J. Macrophage polarization and plasticity in health and disease. Immunol. Res. 2012, 53, 11–24. [Google Scholar] [CrossRef]
- Seneviratne, A.; Edsfeldt, A.; Cole, J.; Kassiteridi, C.; Swart, M.; Park, I.; Green, P.; Khoyratty, T.; Saliba, D.; Goddard, M.; et al. Interferon Regulatory Factor 5 Controls Necrotic Core Formation in Atherosclerotic Lesions by Impairing Efferocytosis. Circulation 2017, 136, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Dalmas, E.; Toubal, A.; Alzaid, F.; Blazek, K.; Eames, H.; Lebozec, K.; Pini, M.; Hainault, I.; Montastier, E.; Denis, R.; et al. IRF5 deficiency in macrophages promotes beneficial adipose tissue expansion and insulin sensitivity during obesity. Nat. Med. 2015, 21, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.; Byrne, A.; Blazek, K.; Saliba, D.; Pease, J.; Perocheau, D.; Feldmann, M.; Udalova, I. IRF5 controls both acute and chronic inflammation. Proc. Natl. Acad. Sci. USA 2015, 112, 11001–11006. [Google Scholar] [CrossRef] [Green Version]
- Courties, G.; Heidt, T.; Sebas, M.; Iwamoto, Y.; Jeon, D.; Truelove, J.; Tricot, B.; Wojtkiewicz, G.; Dutta, P.; Sager, H.; et al. In vivo silencing of the transcription factor IRF5 reprograms the macrophage phenotype and improves infarct healing. J. Am. Coll. Cardiol. 2014, 63, 1556–1566. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.; Solus, J.; Oeser, A.; Li, C.; Raggi, P.; Smith, J.; Stein, C. Genetic Variation and Coronary Atherosclerosis in Patients with Systemic Lupus Erythematosus. Lupus 2014, 23, 876–880. [Google Scholar] [CrossRef] [Green Version]
- Mälarstig, A.; Sigurdsson, S.; Eriksson, P.; Paulsson-Berne, G.; Hedin, U.; Wallentin, L.; Siegbahn, A.; Hamsten, A.; Syvänen, A. Variants of the Interferon Regulatory Factor 5 Gene Regulate Expression of IRF5 mRNA in Atherosclerotic Tissue but Are Not Associated with Myocardial Infarction. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 975–982. [Google Scholar] [CrossRef] [Green Version]
- Kvist, H.; Chowdhury, B.; Grangård, U.; Tylén, U.; Sjöström, L. Total and visceral adipose-tissue volumes derived from measurements with computed tomography in adult men and women: Predictive equations. Am. J. Clin. Nutr. 1988, 48, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Mautner, G.C.; Mautner, S.L.; Froehlich, J.; Feuerstein, I.M.; Proschan, M.A.; Roberts, W.C.; Doppman, J.L. Coronary artery calcification: Assessment with electron beam CT and histomorphometric correlation. Radiology 1994, 192, 619–623. [Google Scholar] [CrossRef]
- Posadas-Sánchez, R.; López-Uribe, Á.R.; Posadas-Romero, C.; Pérez-Hernández, N.; Rodríguez-Pérez, J.M.; Ocampo-Arcos, W.A.; Fragoso, J.M.; Cardoso-Saldaña, G.; Vargas-Alarcón, G. Association of the I148M/PNPLA3 (rs738409) polymorphism with premature coronary artery disease, fatty liver, and insulin resistance in type 2 diabetic patients and healthy controls. The GEA study. Immunobiology 2017, 222, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Posadas-Sánchez, R.; Pérez-Hernández, N.; Angeles-Martínez, J.; López-Bautista, F.; Villarreal-Molina, T.; Rodríguez-Pérez, J.M.; Fragoso, J.M.; Posadas-Romero, C.; Vargas-Alarcón, G. Interleukin 35 Polymorphisms Are Associated with Decreased Risk of Premature Coronary Artery Disease, Metabolic Parameters, and IL-35 Levels: The Genetics of Atherosclerotic Disease (GEA) Study. Mediat. Inflamm. 2017, 2017, 6012795. [Google Scholar] [CrossRef]
- Posadas-Sánchez, R.; Posadas-Romero, C.; Cardoso-Saldaña, G.; Vargas-Alarcón, G.; Villarreal-Molina, M.T.M.T.; Pérez-Hernández, N.; Rodríguez-Pérez, J.M.J.M.; Medina-Urrutia, A.; Jorge-Galarza, E.; Juárez-Rojas, J.G.J.G.; et al. Serum magnesium is inversely associated with coronary artery calcification in the Genetics of Atherosclerotic Disease (GEA) study. Nutr. J. 2016, 15, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posadas-Romero, C.; Jorge-Galarza, E.; Posadas-Sánchez, R.; Acuña-Valerio, J.; Juárez-Rojas, J.G.J.G.; Kimura-Hayama, E.; Medina-Urrutia, A.; Cardoso-Saldaña, G.C.G.C. Fatty liver largely explains associations of subclinical hypothyroidism with insulin resistance, metabolic syndrome, and subclinical coronary atherosclerosis. Eur. J. Endocrinol. 2014, 171, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Cardoso-Saldaña, G.; Fragoso, J.; Lale-Farjat, S.; Torres-Tamayo, M.; Posadas-Romero, C.; Vargas-Alarcón, G.; Posadas-Sánchez, R. The rs10455872-G allele of the LPA gene is associated with high lipoprotein(a) levels and increased aortic valve calcium in a Mexican adult population. Genet. Mol. Biol. 2019, 42, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Baecke, J.A.; Burema, J.; Frijters, J.E. A short questionnaire for the measurement of habitual physical activity in epidemiological studies. Am. J. Clin. Nutr. 1982, 36, 936–942. [Google Scholar] [CrossRef]
- Silva-Zolezzi, I.; Hidalgo-Miranda, A.; Estrada-Gil, J.; Fernandez-Lopez, J.; Uribe-Figueroa, L.; Contreras, A.; Balam-Ortiz, E.; del Bosque-Plata, L.; Velazquez-Fernandez, D.; Lara, C.; et al. Analysis of genomic diversity in Mexican Mestizo populations to develop genomic medicine in Mexico. Proc. Natl. Acad. Sci. USA 2009, 106, 8611–8616. [Google Scholar] [CrossRef] [Green Version]
- Labos, C.; Thanassoulis, G. Genetic Risk Prediction for Primary and Secondary Prevention of Atherosclerotic Cardiovascular Disease: An Update. Curr. Cardiol. Rep. 2018, 20, 36. [Google Scholar] [CrossRef]
- Daly, M.; Rioux, J.; Schaffner, S.; Hudson, T.; Lander, E. High-resolution haplotype structure in the human genome. Nat. Genet. 2001, 29, 229–232. [Google Scholar] [CrossRef]
- Drysdale, C.; McGraw, D.; Stack, C.; Stephens, J.; Judson, R.; Nandabalan, K.; Arnold, K.; Ruano, G.; Liggett, S. Complex promoter and coding region beta 2-adrenergic receptor haplotypes alter receptor expression and predict in vivo responsiveness. Proc. Natl. Acad. Sci. USA 2000, 97, 10483–10488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Yi, L.; Guo, X.; Liu, M.; Li, H.; Zou, H.; Gu, Y.; Tu, W.; Guo, G.; Yang, L.; et al. Association of the IRF5 SNP rs2004640 with systemic sclerosis in Han Chinese. Int. J. Immunopathol. Pharmacol. 2014, 27, 635–638. [Google Scholar] [CrossRef] [Green Version]
- Dieude, P.; Dawidowicz, K.; Guedj, M.; Legrain, Y.; Wipff, J.; Hachulla, E.; Diot, E.; Sibilia, J.; Mouthon, L.; Cabane, J.; et al. Phenotype-haplotype correlation of IRF5 in systemic sclerosis: Role of 2 haplotypes in disease severity. J. Rheumatol. 2010, 37, 987–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiri Jahromi, R.; Nasiri, M.; Jahromi, A. Interferon Regulatory Factor 5 Gene Polymorphisms in Iranian Women with Unexplained Recurrent Pregnancy Loss. Immunol. Invest. 2017, 46, 97–107. [Google Scholar] [CrossRef]
- Sindhu, S.; Kochumon, S.; Thomas, R.; Bennakhi, A.; Al-Mulla, F.; Ahmad, R. Enhanced Adipose Expression of Interferon Regulatory Factor (IRF)-5 Associates with the Signatures of Metabolic Inflammation in Diabetic Obese Patients. Cells 2020, 9, 730. [Google Scholar] [CrossRef] [Green Version]
- Watkins, A.; Yasuda, K.; Wilson, G.; Aprahamian, T.; Xie, Y.; Maganto-Garcia, E.; Shukla, P.; Oberlander, L.; Laskow, B.; Menn-Josephy, H.; et al. IRF5 deficiency ameliorates lupus but promotes atherosclerosis and metabolic dysfunction in a mouse model of lupus-associated atherosclerosis. J. Immunol. 2015, 194, 1467–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Study Groups | |||
|---|---|---|---|
| Controls (n = 1003) | pCAD (n = 1116) | p * | |
| Demographic and clinical characteristics | |||
| Age (years) | 51 ± 9 | 54 ± 8 | <0.001 |
| Sex (% male) | 41.6 | 81.5 | <0.001 |
| Body mass index (kg/m2) | 27.9 (25.4–30.7) | 28.3 (26.0–31.1) | 0.004 |
| Biochemical profile | |||
| HDL-cholesterol (mg/dL) | 45 (36–65) | 37 (31–44) | <0.001 |
| LDL-cholesterol (mg/dL) | 115 (95–134) | 91 (68–116) | <0.001 |
| Triglycerides (mg/dL) | 144 (107–203) | 162 (120–219) | <0.001 |
| Non-HDL-cholesterol (mg/dL) | 142 (121–164) | 120 (93–151) | <0.001 |
| Apolipoprotein B | 94 (76–113) | 80 (63–103) | <0.001 |
| Triglycerides/HDL-cholesterol | 3.2 (2.1–5.3) | 4.3 (3.0–6.6) | <0.001 |
| Aspartate aminotransferase (UI) | 24 (21–30) | 26 (22–31) | 0.001 |
| Adiponectin | 8.2 (5.0–12.6) | 5.2 (3.2–8.1) | <0.001 |
| Uric acid (mg/dL) | 5.4 (4.4–6.4) | 6.4 (5.4–7.4) | <0.001 |
| Tumor necrosis factor alpha (pg/mL) | 0.56 (0.01–1.81) | 0.53 (0.06–1.65) | 0.377 |
| Interleukin 6 (pg/mL) | 0.83 (0.40–1.71) | 0.93 (0.50–2.04) | 0.011 |
| Interleukin 15 (pg/mL) | 1.46 (0.34–2.94) | 1.30 (0.67–2.06) | 0.022 |
| Lifestyle | |||
| Current smoking habit (%) | 23.3 | 11.6 | <0.001 |
| Physical activity | 7.9 (7.0–8.8) | 7.5 (6.8–8.4) | <0.001 |
| Study Groups | |||
|---|---|---|---|
| Controls (n = 1003) | pCAD (n = 1116) | p * | |
| Obesity (%) | 29.7 | 34.9 | 0.012 |
| Hypertension (%) | 18.7 | 68.0 | <0.001 |
| Type 2 diabetes mellitus (%) | 10.3 | 35.5 | <0.001 |
| Hypoalphalipoproteinemia | 51.4 | 67.8 | <0.001 |
| High LDL-cholesterol (≥130 mg/dL, %) | 29.3 | 16.5 | <0.001 |
| Hypertriglyceridemia (%) | 33.2 | 42.8 | <0.001 |
| High non-HDL-cholesterol (>160 mg/dL, %) | 27.9 | 19.6 | <0.001 |
| High apolipoprotein B (≥110 mg/dL, %) | 28.2 | 19.5 | <0.001 |
| High triglycerides/HDL-cholesterol index (>3.0, %) | 52.6 | 75.2 | <0.001 |
| Elevated aspartate aminotransferase (%) | 35.9 | 38.3 | 0.259 |
| Hypoadiponectinemia (<p25, %) | 42.7 | 58.1 | <0.001 |
| Hyperuricemia (%) | 20.4 | 36.0 | <0.001 |
| Elevated TNFα (>p75, %) | 29.9 | 23.3 | 0.001 |
| Elevated interleukin 6 (>p75, %) | 29.2 | 29.8 | 0.803 |
| Elevated interleukin 15 (>p75, %) | 36.5 | 19.4 | <0.001 |
| Magnesium deficiency (%) | 5.0 | 9.3 | <0.001 |
| Valvular calcification (%) | 10.6 | nd | |
| Subclinical hypothyroidism (%) | 17.3 | 16.3 | 0.592 |
| Haplotypes | pCAD | Controls | OR (95% CI) | p | |
|---|---|---|---|---|---|
| H1 | TGCGTCT | 0.427 | 0.374 | 1.248 (1.102–1.413) | 0.0005 |
| H2 | CCCTTTT | 0.407 | 0.379 | 1.128 (0.996–1.278) | 0.0569 |
| H3 | TCCTTTT | 0.061 | 0.048 | 1.285 (0.980–1.684) | 0.0700 |
| H4 | TCTGCCT | 0.047 | 0.004 | 10.73 (5.416–21.26) | <0.0001 |
| H5 | TCCGTCT | 0.007 | 0.042 | 0.155 (0.089–0.269) | <0.0001 |
| H6 | TCCGTCC | 0.023 | 0.018 | 1.303 (0.843–2.012) | 0.2331 |
| H7 | CGCTTTT | 0.004 | 0.032 | 0.108 (0.051–0.226) | <0.0001 |
| H8 | TCCGCCT | 0.001 | 0.031 | 0.014 (0.002–0.102) | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Posadas-Sánchez, R.; Cardoso-Saldaña, G.; Fragoso, J.M.; Vargas-Alarcón, G. Interferon Regulatory Factor 5 (IRF5) Gene Haplotypes Are Associated with Premature Coronary Artery Disease. Association of the IRF5 Polymorphisms with Cardiometabolic Parameters. The Genetics of Atherosclerotic Disease (GEA) Mexican Study. Biomolecules 2021, 11, 443. https://doi.org/10.3390/biom11030443
Posadas-Sánchez R, Cardoso-Saldaña G, Fragoso JM, Vargas-Alarcón G. Interferon Regulatory Factor 5 (IRF5) Gene Haplotypes Are Associated with Premature Coronary Artery Disease. Association of the IRF5 Polymorphisms with Cardiometabolic Parameters. The Genetics of Atherosclerotic Disease (GEA) Mexican Study. Biomolecules. 2021; 11(3):443. https://doi.org/10.3390/biom11030443
Chicago/Turabian StylePosadas-Sánchez, Rosalinda, Guillermo Cardoso-Saldaña, José Manuel Fragoso, and Gilberto Vargas-Alarcón. 2021. "Interferon Regulatory Factor 5 (IRF5) Gene Haplotypes Are Associated with Premature Coronary Artery Disease. Association of the IRF5 Polymorphisms with Cardiometabolic Parameters. The Genetics of Atherosclerotic Disease (GEA) Mexican Study" Biomolecules 11, no. 3: 443. https://doi.org/10.3390/biom11030443
APA StylePosadas-Sánchez, R., Cardoso-Saldaña, G., Fragoso, J. M., & Vargas-Alarcón, G. (2021). Interferon Regulatory Factor 5 (IRF5) Gene Haplotypes Are Associated with Premature Coronary Artery Disease. Association of the IRF5 Polymorphisms with Cardiometabolic Parameters. The Genetics of Atherosclerotic Disease (GEA) Mexican Study. Biomolecules, 11(3), 443. https://doi.org/10.3390/biom11030443