
Multi-Enzymatic Cascades in the Synthesis of Modified Nucleosides: Comparison of the Thermophilic and Mesophilic Pathways

 , ,
, ,  , , , ,
, , , ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Procedures
2.2. Purification of Recombinant Enzymes
2.3. Cloning, Expression, and Purification of Recombinant TthPPM
2.4. Analytical Methods
2.5. Enzyme Assay
2.6. Determination of Kinetic Parameters of Phosphopentomutase Thermus Thermophilus HB27 (TthPPM)
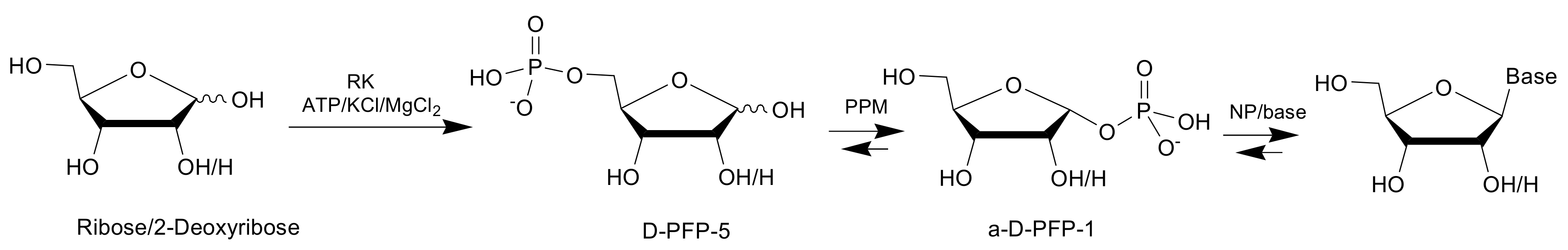
2.7. Cascade Reactions
2.8. Nucleosides Synthesis
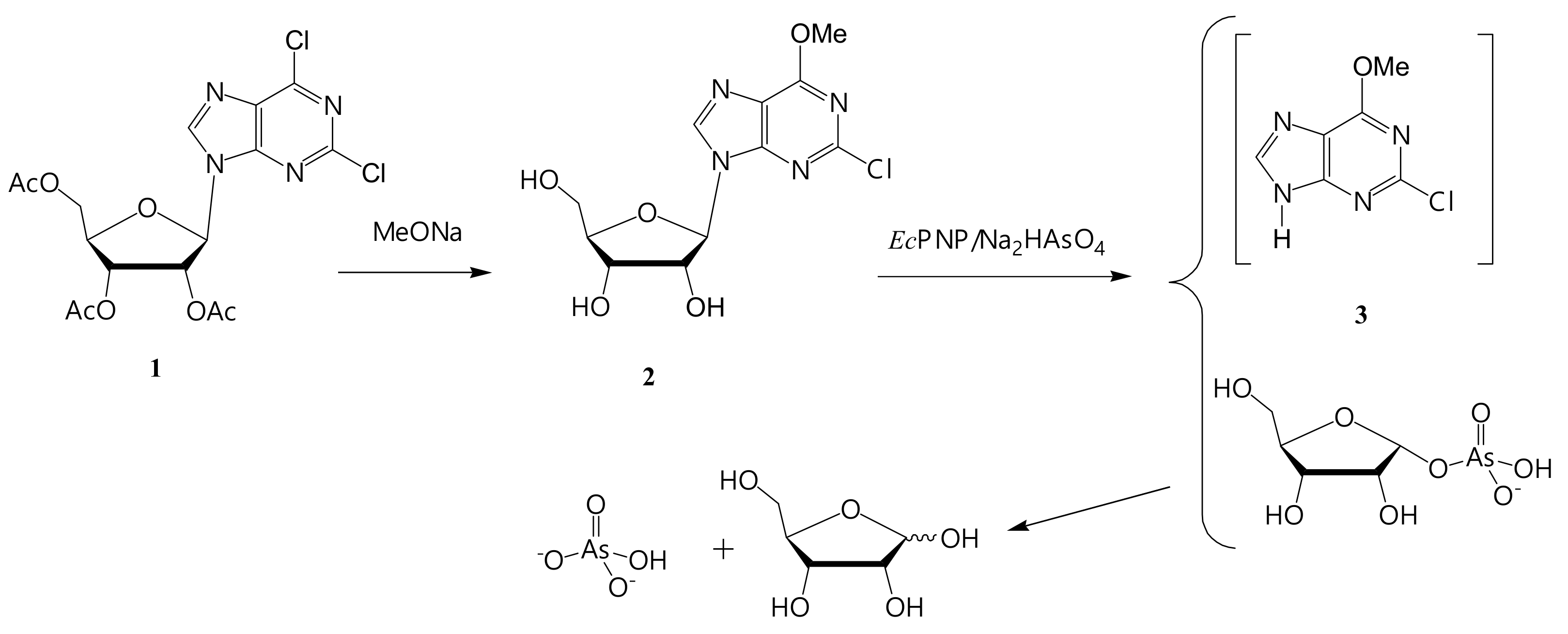
2.8.1. 2-Chloro-6-methoxy-9-(β-D-ribofuranosyl)purine (2)
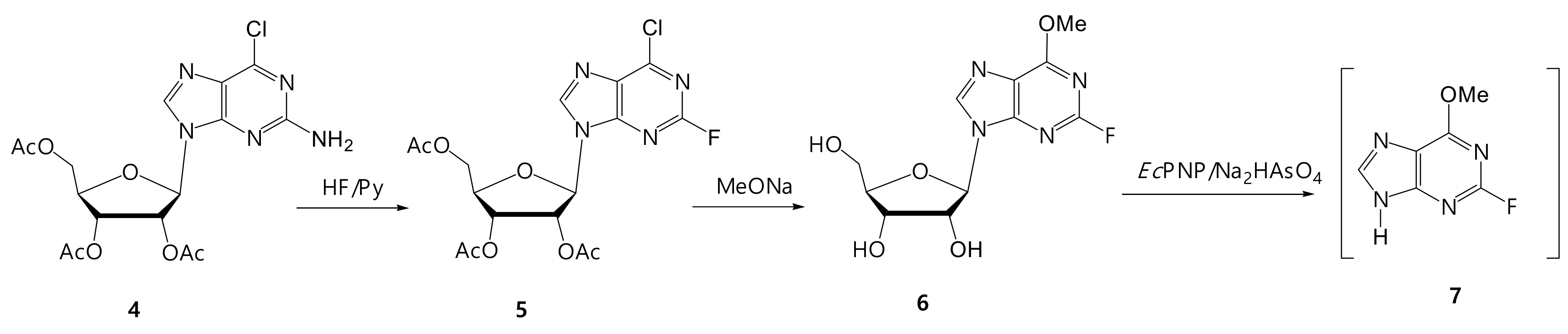
2.8.2. 2-Fluoro-6-chloro-9-(2’,3,’5′-tri-O-acetyl-β-D-ribofuranosyl)purine (5)
2.8.3. 2-Fluoro-6-methoxy-9-(β-D-ribofuranosyl)purine (6)
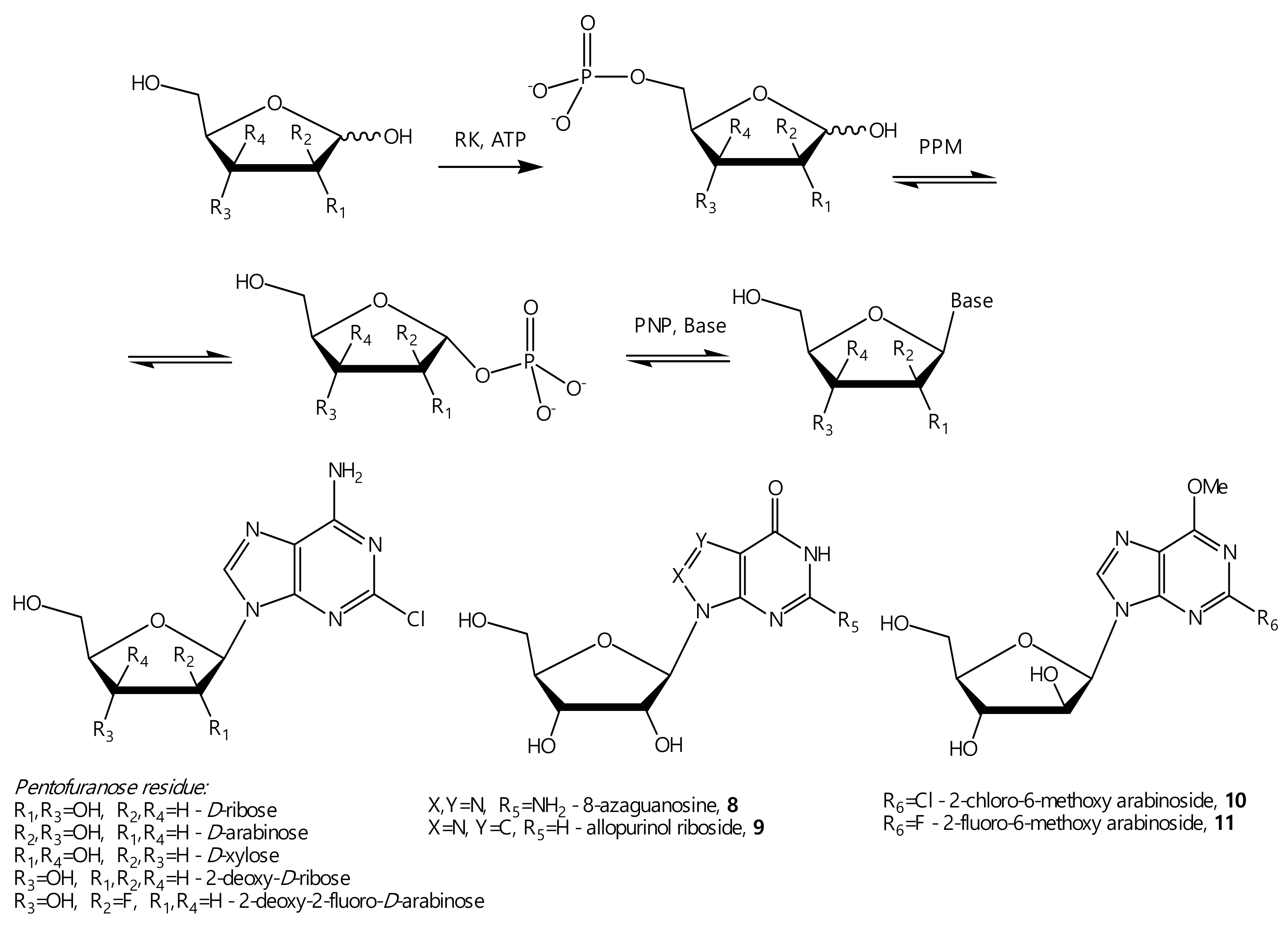
2.8.4. 5-Amino-3-(β-D-ribofuranosyl)triazolo [4,5-d]pyrimidin-7-one (8)
2.8.5. 1-(β-D-Ribofuranosyl)pyrazolo[3,4-d]pyrimidine-4-one (9)
2.8.6. 9-(β-D-Arabinofuranosyl)-2-chloro-6-methoxypurine (10)
2.8.7. 9-(β-D-Arabinofuranosyl)-2-fluoro-6-methoxypurine (11)
3. Results
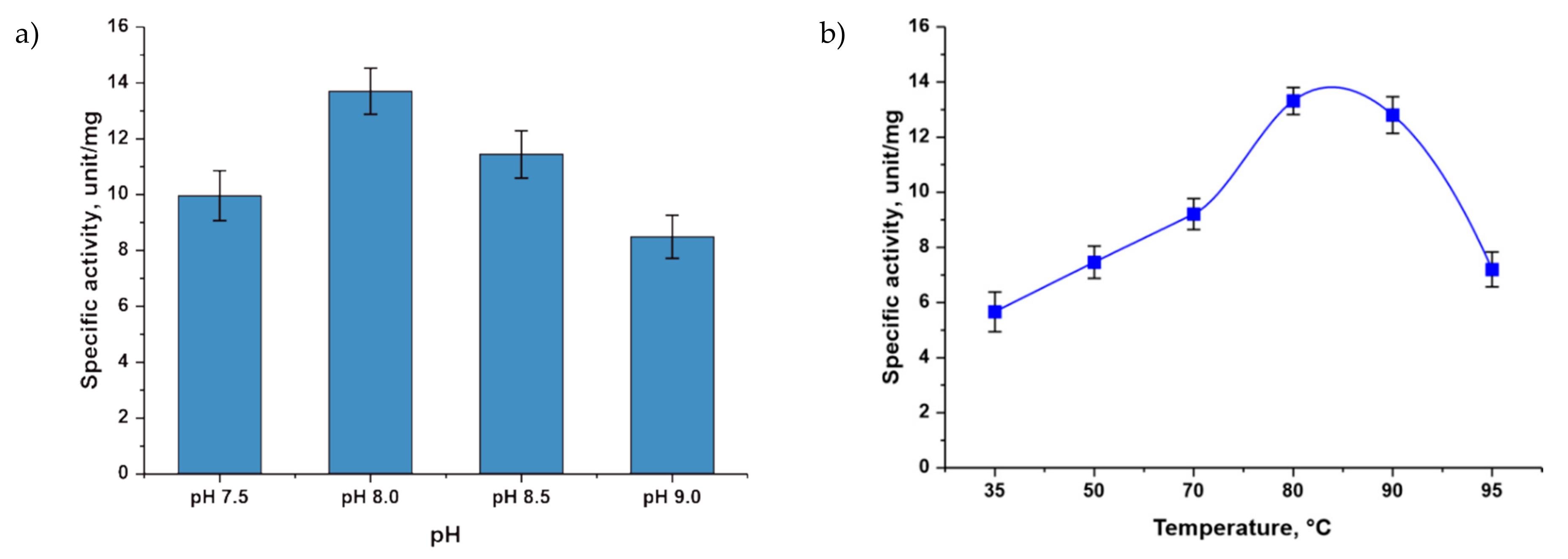
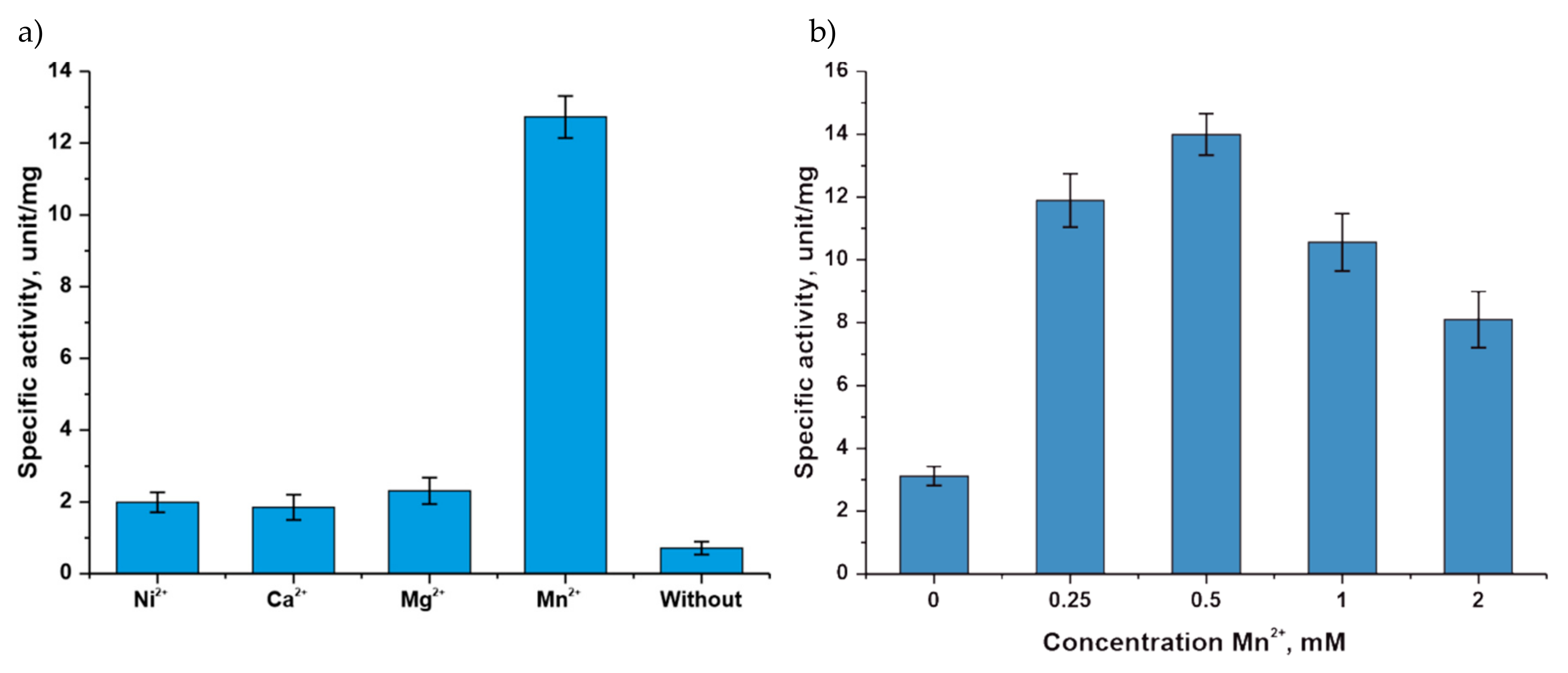
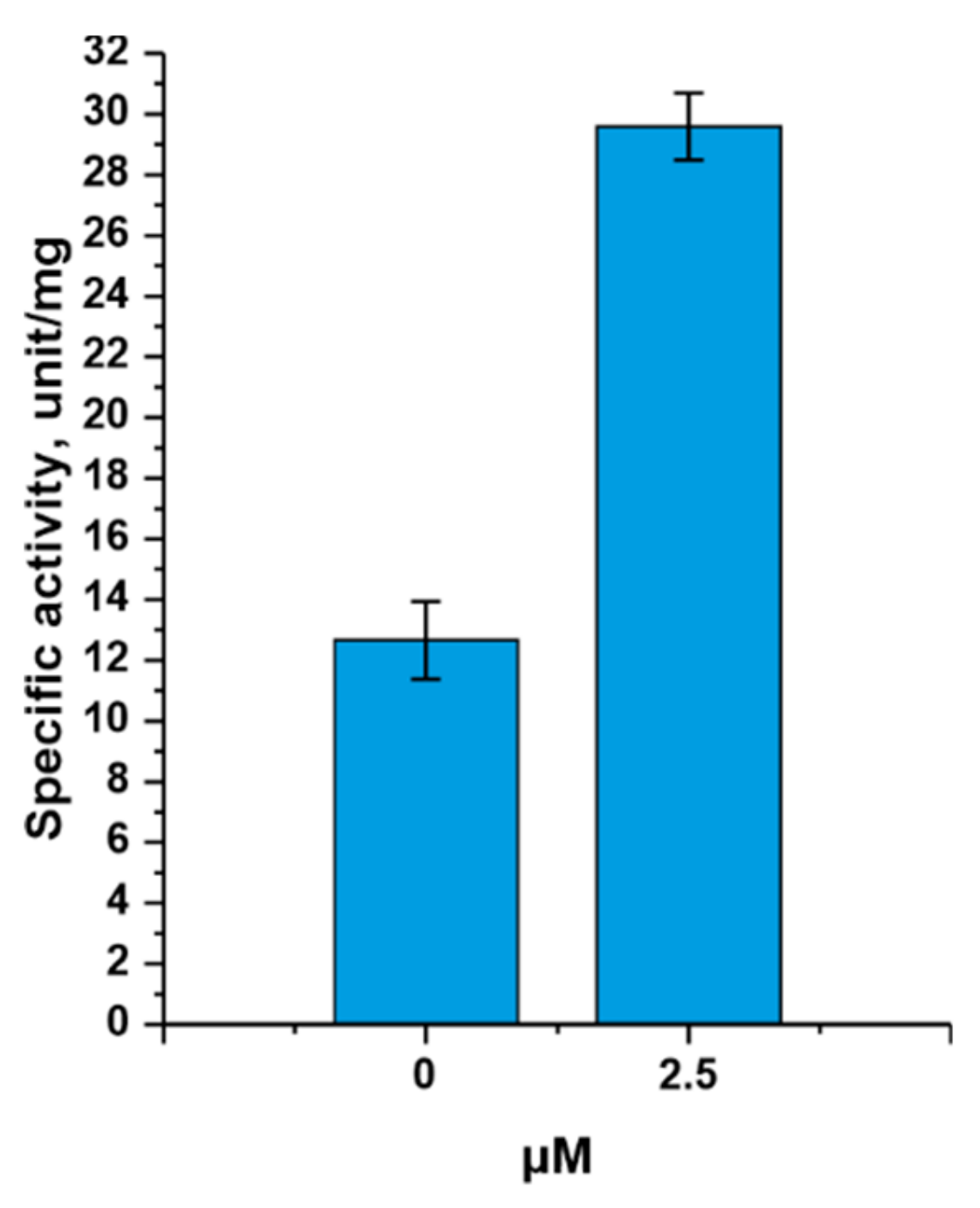
3.1. Study of the Influence of Various Factors on TthPPM Activity
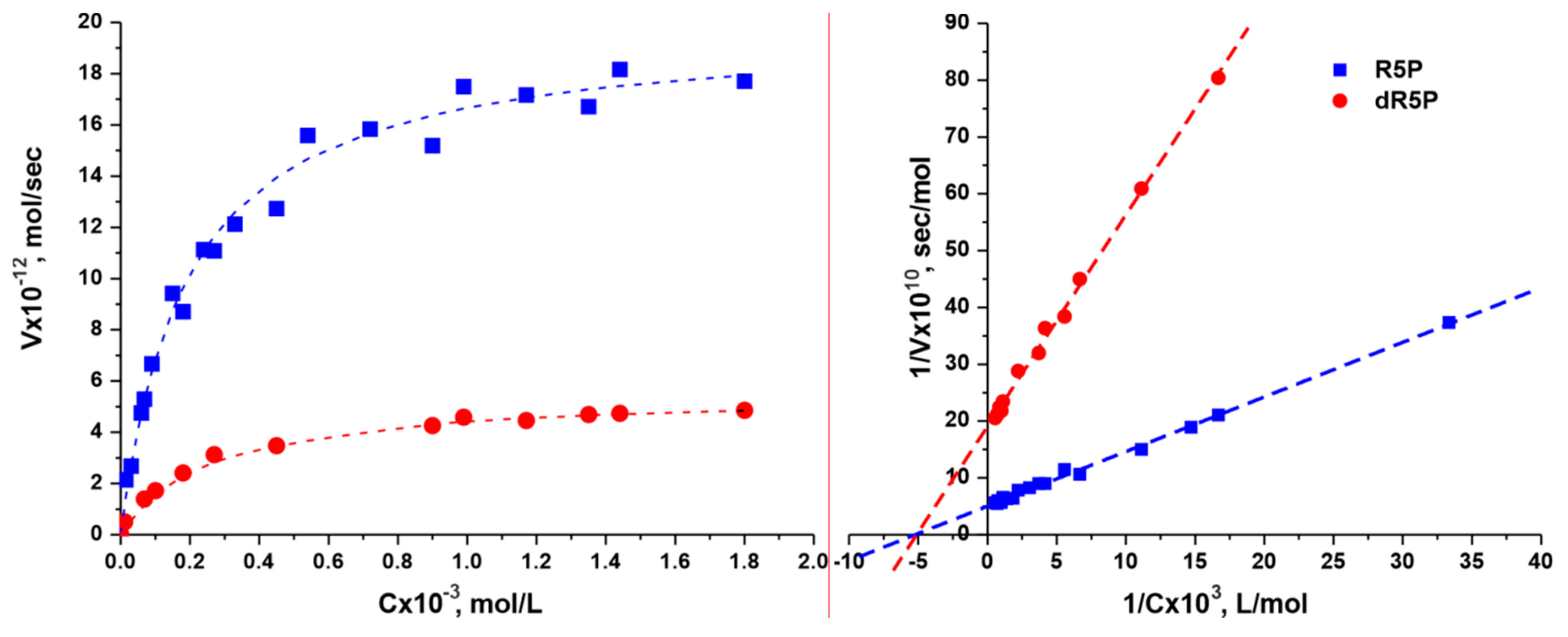
3.2. Determination of the Kinetic Parameters for TthPPM
3.3. Synthesis of Modified Heterocyclic Bases for Cascade Synthesis of Nucleosides
3.3.1. Synthesis of 2-Chloro-6-methoxypurine (3)
3.3.2. Synthesis of 2-Fluoro-6-methoxypurine (7)
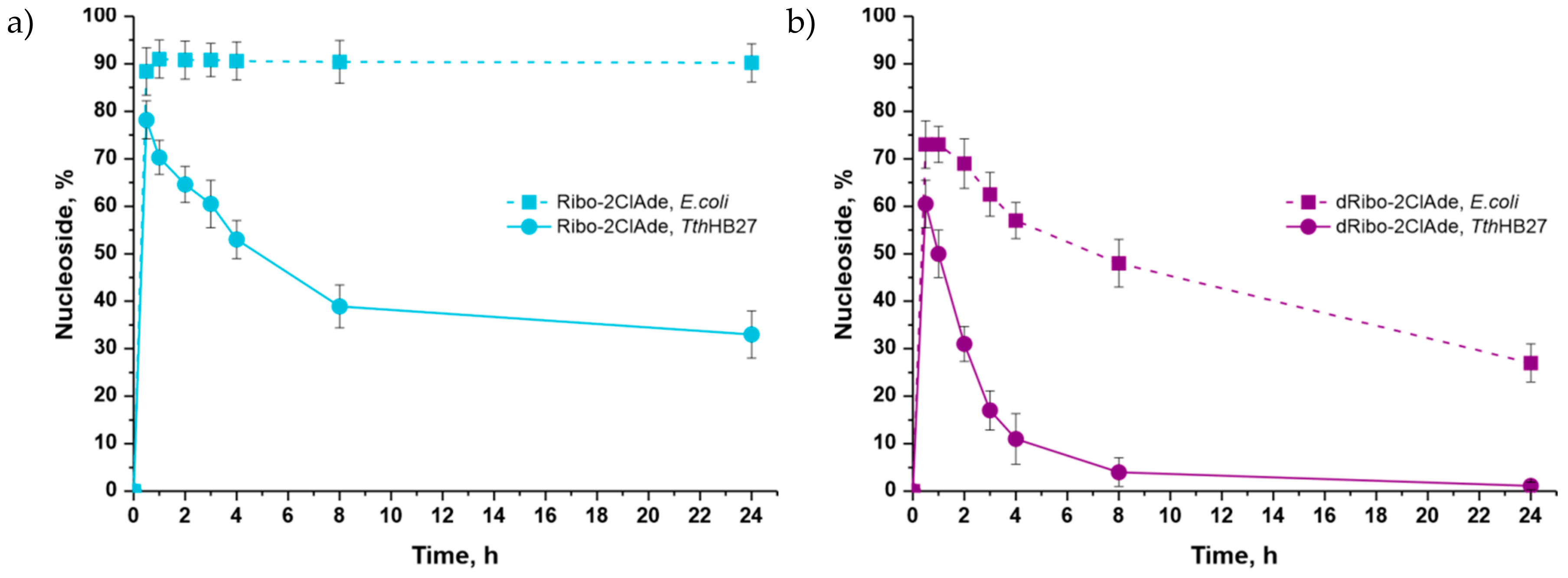
3.4. Conditions for Poly-Enzymatic Cascades Using Mesophilic and Thermophilic Enzymes
3.5. Synthesis of Nucleosides (8–11)
4. Discussion
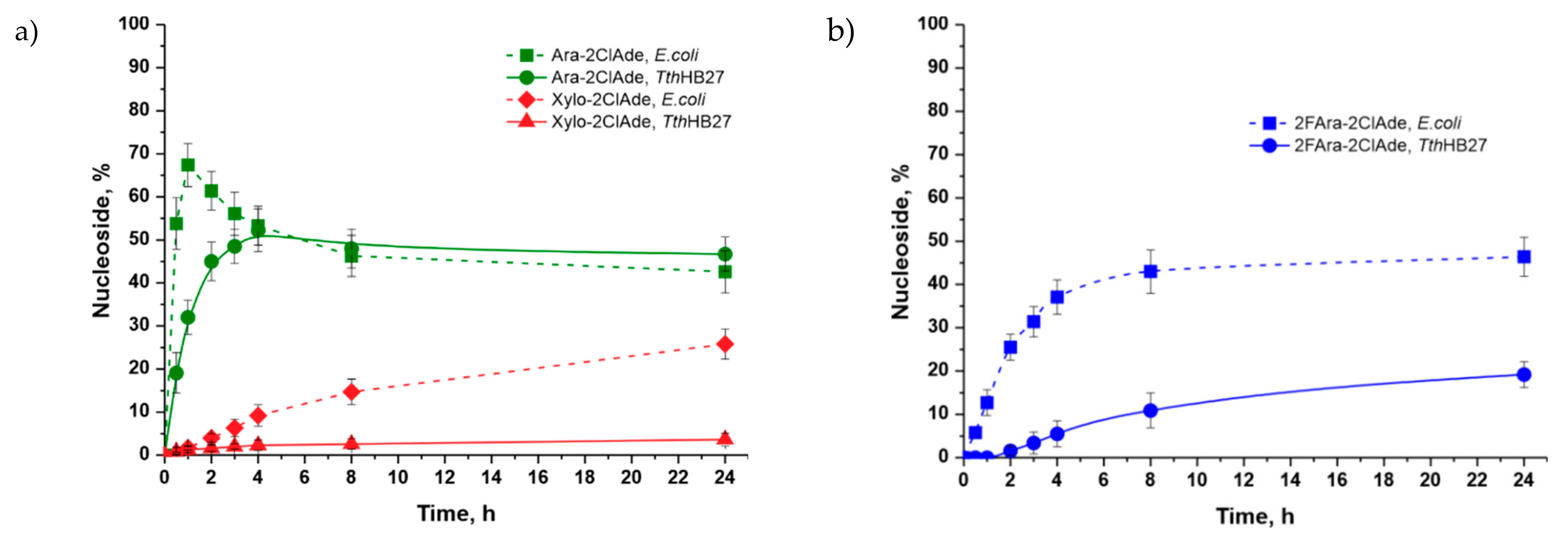
4.1. Possibilities of Using Various D-Pentoses in the Synthesis of 2-Chloroadenine Nucleosides
4.2. Non-Natural Heterocyclic Bases in the Cascade Synthesis of Nucleosides
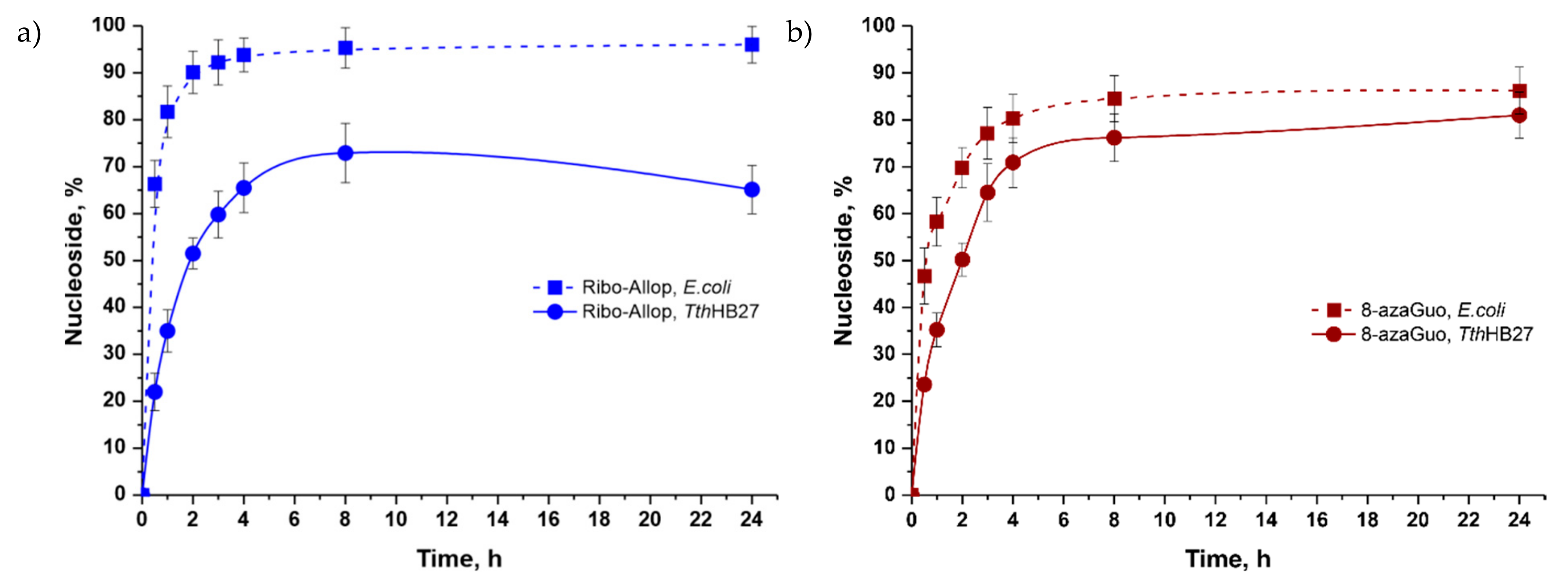
Synthesis of 8-Azaguanine and Allopurinol Ribosides
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| a-D-PFP-1 | 1-α-phosphate D-pentofuranose |
| Allop | allopurinol |
| Ara | D-arabinose |
| Ara-2ClAde | 2-chloradenine arabinoside |
| 8-Aza-Gua | 8-azaguanie |
| BSA | bovine serum albumin |
| dNTP | deoxynucleoside triphosphate mix |
| D-PFP-5 | 5-phosphate D-pentofuranose |
| dRibo, dRib | 2-deoxy-D-ribose |
| DTT | dithiothreitol |
| Ec | Escherichia coli |
| EDTA | ethylenediaminetetraacetic acid |
| F-Ara | 2-deoxy-2-fluoro-D-arabinose |
| HMBC | heteronuclear multiple bond correlation spectroscopy |
| HSQC | heteronuclear single quantum correlation spectroscopy |
| kDa | kilodalton |
| IPTG | isopropyl β-D-1-thiogalactopyranoside |
| NP | nucleoside phosphorylase |
| PMSF | phenylmethylsulphonyl fluoride |
| PNP | purine nucleoside phosphorylase |
| PPM | phosphopentomutase |
| PyNP | pyrimidine nucleoside phosphorylase |
| Ribo, Rib | D-ribose |
| RK | ribokinase |
| TEMED | tetramethylethylenediamine |
| TP | thymidine phosphorylase |
| Tsp | Thermus species 2.9 |
| Tth | Thermus thermophilus |
| UP | uridine phosphorylase |
| Xyl | D-xylose |
| Xylo-2ClAde | 2-chloroadenine xyloside |
References
- Fernández-Lucas, J.; Fresco-Taboada, A.; Acebal, C.; de la Mata, I.; Arroyo, M. Enzymatic synthesis of nucleoside analogues using immobilized 2′-deoxyribosyltransferase from Lactobacillus reuteri. Appl. Microbiol. Biotechol. 2011, 91, 317–327. [Google Scholar] [CrossRef]
- Boryski, J. Reactions of transglycosylation in the nucleoside chemistry. Curr. Org. Chem. 2008, 12, 309–325. [Google Scholar] [CrossRef]
- Fernández-Lucas, J. Multienzymatic synthesis of nucleic acid derivatives: A general perspective. Appl. Microbiol. Biotechol. 2015, 99, 4615–4627. [Google Scholar] [CrossRef] [PubMed]
- Del Arco, J.; Fernández-Lucas, J. Purine and pyrimidine salvage pathway in thermophiles: A valuable source of biocatalysts for the industrial production of nucleic acid derivatives. Appl. Microbiol. Biotechnol. 2018, 102, 7805–7820. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q. Synthesis of nucleic acid derivatives by multi-enzymatic systems. In Enzymatic and Chemical Synthesis of Nucleic Acid Derivatives; Fernandez-Lucas, J., Camarasa Rius, M.J., Eds.; Wiley-VCH: Weinheim, Germany, 2019; Volume 6, pp. 129–157. [Google Scholar]
- Lewkowicz, E.S.; Iribarren, A.M. Nucleoside phosphorylases. Cur. Org. Chem. 2006, 10, 1197–1215. [Google Scholar] [CrossRef]
- Michailidou, F.; Burnett, D.; Sharma, S.V.; Van Lanen, S.G.; Goss, R.J.M. Natural products incorporating pyrimidine nucleosides. Compr. Nat. Prod. III Third Ed. 2020, 2, 500–536. [Google Scholar]
- Michailidou, F.; Chung, C.; Brown, M.J.B.; Bent, A.F.; Naismith, J.H.; Leavens, W.J.; Lynn, S.M.; Sharma, S.V.; Goss, R.J.M. Pac13 is a small, monomeric dehydratase that mediates the formation of the 3′-deoxy nucleoside of pacidamycins. Angew. Chem. Int. Ed. 2017, 56, 12492–12497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikhailopulo, I.A.; Miroshnikov, A.I. Biologically important nucleosides: Modern trends in biotechnology and application. Mendeleev Commun. 2011, 21, 57–68. [Google Scholar] [CrossRef]
- Lapponi, M.J.; Rivero, C.W.; Zinni, M.A.; Britos, C.N.; Trelles, J.A. New developments in nucleoside analogues biosynthesis: A review. J. Mol. Catal. B Enzym. 2016, 133, 218–233. [Google Scholar] [CrossRef]
- De Benedetti, E.C.; Rivero, C.W.; Britos, C.N.; Lozano, M.E.; Trelles, J.A. Biotransformation of 2,6-diaminopurine nucleosides by immobilized Geobacillus stearothermophilus. Biotechnol. Progr. 2012, 28, 1251–1256. [Google Scholar] [CrossRef]
- Almendros, M.; Berenguer, J.; Sinisterra, J.-V. Thermus thermophilus Nucleoside phosphorylases active in the synthesis of nucleoside analogues. Appl. Environ. Microbiol. 2012, 78, 3128–3135. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, F.; Neubauer, P.; Kurreck, A. The peculiar case of the hyperthermostable pyrimidine nucleoside phosphorylase from Thermus thermophilus. Chembiochem A Eur. J. Chem. Biol. 2021, 22, 1–7. [Google Scholar] [CrossRef]
- Hori, N.; Watanabe, M.; Sunagawa, K.; Uehara, K.; Mikami, Y. Production of 5-methyluridine by immobilized thermostable purine nucleoside phosphorylase and pyrimidine nucleoside phosphorylase from Bacillus stearothermophilus JTS 85. J. Biotechnol. 1991, 17, 121–131. [Google Scholar] [CrossRef]
- Taran, S.A.; Verevkina, K.N.; Feofanov, S.A.; Miroshnikov, A.I. Enzymatic transglycosylation of natural and modified nucleosides by immobilized thermostable nucleoside phosphorylases from Geobacillus stearothermophilus. Rus. J. Bioorg. Chem. 2009, 35, 739–745. [Google Scholar] [CrossRef]
- Zhou, X.; Mikhailopulo, I.A.; Bournazou, M.N.; Neubauer, P. Immobilization of thermostable nucleoside phosphorylases on MagReSyn® epoxide microspheres and their application for the synthesis of 2,6-dihalogenated purine nucleosides. J. Mol. Catal. B Enzym. 2015, 115, 119–127. [Google Scholar] [CrossRef]
- Michailidou, F.; Lebl, T.; Slawin, A.M.Z.; Sharma, S.V.; Brown, M.J.B.; Goss, R.J.M. Fluorinated uridine analogues provide insight into a neighbouring-group participation mechanism. Molecules 2020, 25, 5513. [Google Scholar] [CrossRef]
- Miroshnikov, A.I.; Esipov, R.S.; Muravyova, T.I.; Konstantinova, I.D.; Fateev, I.V.; Mikhailopulo, I.A. A new strategy for the synthesis of nucleosides: One-pot enzymatic transformation of D-pentoses into nucleosides. Open Conf. Proc. J. 2010, 1, 98–102. [Google Scholar] [CrossRef] [Green Version]
- Panosian, T.D.; Nannemann, D.P.; Watkins, G.R.; Phelan, V.V.; McDonald, W.H.; Wadzinski, B.E.; Iverson, T. M Bacillus cereus phosphopentomutase is an alkaline phosphatase family member that exhibits an altered entry point into the catalytic cycle. J. Biol. Chem. 2011, 286, 8043–8054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taverna-Porro, M.; Bouvier, L.A.; Pereira, C.A.; Montserrat, J.M.; Iribarren, A.M. Chemoenzymatic preparation of nucleosides from furanoses. Tetrahedron Lett. 2008, 49, 2642–2645. [Google Scholar] [CrossRef]
- Stepchenko, V.A.; Seela, F.; Esipov, R.S.; Miroshnikov, A.I.; Sokolov, Y.A.; Mikhailopulo, I.A. Enzymatic synthesis of 2′-deoxy-β-d-ribonucleosides of 8-azapurines and 8-aza-7-deazapurines. Synlett 2012, 23, 1541–1545. [Google Scholar]
- Esipov, R.S.; Abramchik, Y.A.; Fateev, I.V.; Konstantinova, I.D.; Kostromina, M.A.; Muravyova, T.I.; Artemova, K.G.; Miroshnikov, A.I. A cascade of thermophilic enzymes as an approach to the synthesis of modified nucleotides. Acta Nat. 2016, 8, 161–168. [Google Scholar] [CrossRef]
- Esipov, R.S.; Abramchik, Y.A.; Fateev, I.V.; Muravyova, T.I.; Scoblov, Y.S.; Kostromina, M.A.; Miroshnikov, A.I. Preparation and study of the substrate specificity of thermophilic ribokinase from Thermus sp. 2.9. Russ. J. Biopharm. 2016, 8, 3–12. [Google Scholar]
- Timofeev, V.I.; Fateev, I.V.; Kostromina, M.A.; Abramchik, Y.A.; Konstantinova, I.D.; Volkov, V.V.; Lykoshin, D.D.; Mikheeva, O.O.; Muravieva, T.I.; Esipov, R.S.; et al. The comparative analysis of the properties and structures of purine nucleoside phosphorylases from thermophilic bacterium Thermus thermophilus HB27. J. Biomol. Struct. Dyn. 2020. [Google Scholar] [CrossRef] [PubMed]
- Robins, M.J.; Uznański, B. Nucleic acid related compounds. 33. Conversions of adenosine and guanosine to 2, 6-dichloro, 2-amino-6-chloro, and derived purine nucleosides. Can. J. Chem. 1981, 59, 2601–2607. [Google Scholar] [CrossRef]
- Robins, M.J.; Zou, R.; Hansske, F.; Madej, D.; Tyrrell, D.L.J. Synthesis, transformation chemistry, and biological activity of guanine nucleosides and analoges. Nucleosides Nucleotides 1989, 8, 725–741. [Google Scholar] [CrossRef]
- Berzin, V.B.; Dorofeeva, E.V.; Leonov, V.N.; Miroshnikov, A.I. The preparative method for 2-fluoroadenosine synthesis. Rus. J. Bioorg. Chem. 2009, 35, 193–196. [Google Scholar] [CrossRef]
- Chuvikovsky, D.V.; Esipov, R.S.; Skoblov, Y.S.; Chupova, L.A.; Muravyova, T.I.; Miroshnikov, A.I.; Lapinjoki, S.; Mikhailopulo, I.A. Ribokinase from E. coli: Expression, purification, and substrate specificity. Bioorg. Med. Chem. 2006, 14, 6327–6332. [Google Scholar] [CrossRef]
- Esipov, R.S.; Gurevich, A.I.; Chuvikovsky, D.V.; Chupova, L.A.; Muravyova, T.I.; Miroshnikov, A.I. Overexpression of Escherichia coli genes encoding nucleoside phosphorylases in the pET/Bl21(DE3) system yields active recombinant enzymes. Protein Expr. Purif. 2002, 24, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Boil. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Konstantinova, I.D.; Fateev, I.V.; Miroshnikov, A.I. The arsenolysis reaction in the biotechnological method of synthesis of modified purine β-D-arabinonucleosides. Rus. J. Bioorg. Chem. 2016, 42, 372–380. [Google Scholar] [CrossRef]
- Fateev, I.V.; Antonov, K.V.; Konstantinova, I.D.; Muravyova, T.I.; Seela, F.; Esipov, R.S.; Miroshnikov, A.I.; Mikhailopulo, I.A. The chemoenzymatic synthesis of clofarabine and related 2′-deoxyfluoroarabinosyl nucleosides: The electronic and stereochemical factors determining substrate recognition by E. coli nucleoside phosphorylases. Beilstein J. Org. Chem. 2014, 10, 1657–1669. [Google Scholar]
- Bunton, C.A.; Hummers, E. The hydrolyses of α-D-ribose and α-D-glucose-1-phosphate. J.Org. Chem. 1969, 34, 572–576. [Google Scholar] [CrossRef]
- Kamel, S.; Weiß, M.; Klare, H.F.T.; Mikhailopulo, I.A.; Neubauer, P.; Wagner, A. Chemo-enzymatic synthesis of α-D-pentofuranose-1-phosphates using thermostable pyrimidine nucleoside phosphorylases. Mol. Catal. 2018, 458, 52–59. [Google Scholar] [CrossRef]
- Bennett, E.M.; Li, C.; Allan, P.W.; Parker, W.B.; Ealick, S.E. Structural Basis for Substrate Specificity of Escherichia coli Purine Nucleoside Phosphorylase. J. Biol. Chem. 2003, 278, 47110–47118. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Enzyme | Gene | Concentration *, mg/mL | Oligomer form ** | Purity, % *** |
|---|---|---|---|---|---|
| TspRK | Ribokinase Thermus species 2.9 | QT17_05185 | 13.4 | 63 kD adimer | 94.3 |
| TthPPM | Phosphopentamutase Thermus thermophilus HB27 | TT_RS08405 | 10.1 | 43 kD amonomer | 70.5 |
| TthPNPI | Purine nucleoside phosphorylase I Thermus thermophilus HB27 | TT_RS05405 | 9.8 | 153 kD ahexamer | 96.4 |
| TthPNPII | Purine nucleoside phosphorylase II Thermus thermophilus HB27 | TT_RS00985 | 10.3 | 179 kD ahexamer | 95.0 |
| EcRK | Ribokinase E. coli K12 | AAA51476.1 | 6.9 | 66 kD adimer | 83.2 |
| EcPPM | Phosphopentamutase E. coli K12 | AAA97279.1 | 11.0 | 44 kD amonomer | 85.4 |
| EcPNP | Purine nucleoside phosphorylase E. coliK12 | AAA24401.1 | 30.1 | 155 kD ahexamer | 95.8 |
| Substrate | KM, mM | Vmax, µmol/min·mg | kcat, s−1 | kcat/KM, s−1·M−1 |
|---|---|---|---|---|
| D-ribose 5-phosphate | 0.19 ± 0.02 | 3.50 ± 0.40 | 2.50 ± 0.30 | 1.3 × 104 |
| 2-deoxy-D-ribose 5-phosphate | 0.25 ± 0.04 | 0.48 ± 0.08 | 0.35 ± 0.06 | 1.4 × 103 |
| Enzyme | Substrate | Activity, Units/mg |
|---|---|---|
| TspRK | D-ribose | 19 ± 3 |
| TthPPM | D-ribose 5-phosphate | 14 ± 3 |
| 2-deoxy-D-ribose 5-phosphate | 2.8 ± 0.7 | |
| TthPNPI | Inosine | 180 ± 30 |
| TthPNPII | Adenosine | 200 ± 30 |
| EcRK | D-ribose | 310 ± 40 |
| EcPPM | D-ribose 5-phosphate | 4.9 ± 0.9 |
| EcPNP | Inosine | 220 ± 30 |
| Adenosine | 120 ± 10 |
| Nucleoside | 2ClAde Conc., mM | ATP Conc., mM | D-Pentose Conc. | Enzyme Ratio, Units | Max. Nucleoside Content, % | ||
|---|---|---|---|---|---|---|---|
| RK | PPM | PNP | |||||
| Thermus thermophilus | |||||||
| Ribo-2ClAde | 0.34 | 2.5 | 2 mM D-ribose | 0.3 * | 0.1 | 0.5 ** | 78 (30 min) |
| dRibo-2ClAde | 2 mM 2-deoxy-D-ribose | 0.3 * | 0.1 | 0.5 ** | 61 (30 min) | ||
| Ara-2ClAde | 60 mM D-arabinose | 11 * | 1 | 10 ** | 52 (4 h) | ||
| Xylo-2ClAde | 60 mM D-xylose | 11 * | 1 | 10 ** | 4 (24 h) | ||
| Clofarabine | 10 mM 2-deoxy-2-fluoro-D-arabinose | 6 * | 1 | 10 ** | 19 (24 h) | ||
| Escherichia coli | |||||||
| Ribo-2ClAde | 0.34 | 2.5 | 2 mM D-ribose | 0.2 | 0.1 | 0.4 | 91 (1 h) |
| dRibo-2ClAde | 2 mM 2-deoxy-D-ribose | 0.2 | 0.1 | 0.4 | 73 (30 min) | ||
| Ara-2ClAde | 60 mM D-arabinose | 5 | 1 | 9 | 67 (1 h) | ||
| Xylo-2ClAde | 60 mM D-xylose | 5 | 1 | 9 | 26 (24 h) | ||
| Clofarabine | 10 mM 2-deoxy-2-fluoro-D-arabinose | 6 | 1 | 4 | 46 (24 h) | ||
| Nucleoside | D-Rib Conc., mM | ATP Conc., mM | Base, 0.5 mM | Enzyme Ratio, Units | Max. Nucleoside Content, % (LC MS) | ||
|---|---|---|---|---|---|---|---|
| RK | PPM | PNP | |||||
| Thermus thermophilus | |||||||
| Ribo-8azaGua (8) | 2 | 2.5 | 8-Aza-Gua | 0.4 * | 0.1 | 0.4 ** | 81 (24 h) |
| Ribo-Allop (9) | Allop | 0.4 * | 0.1 | 0.4 ** | 73 (8 h) | ||
| Escherichia coli | |||||||
| Ribo-8azaGua (8) | 2 | 2.5 | 8-Aza-Gua | 0.2 | 0.1 | 0.4 | 86 (24 h) |
| Ribo-Allop (9) | Allop | 0.2 | 0.1 | 0.4 | 96 (24 h) | ||
| Comp. | D-Pentose, mg (mmol) | Base, mg (mmol) | Vol., mL | Enzymes, U | Time, h /Temp, °C | Isolated Yield, mg (%) | ||
|---|---|---|---|---|---|---|---|---|
| RK | PPM | PNP | ||||||
| Thermus thermophilus | ||||||||
| 8 | Ribose 42.6 (0.284) | 8-Aza-Gua 28 (0.184) | 142 | 11.4 * | 2.8 | 11.4 ** | 24/ 80 | 36.1 (69) |
| Escherichia coli | ||||||||
| 9 | Ribose 42.6 (0.284) | Allop 25 (0.184) | 142 | 5.7 | 2.8 | 11.4 | 24/ 50 | 41.6 (84) |
| 10 | Arabinose 1500 (10) | (2) 80 (0.25) | 167 | 1670 | 334 | 3000 | 48/ 50 | 43.0 (54 ***) |
| 11 | Arabinose 720 (4.8) | (6) 36 (0.12) | 80 | 800 | 160 | 1440 | 48/ 50 | 14.7 (41 ***) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fateev, I.V.; Kostromina, M.A.; Abramchik, Y.A.; Eletskaya, B.Z.; Mikheeva, O.O.; Lukoshin, D.D.; Zayats, E.A.; Berzina, M.Y.; Dorofeeva, E.V.; Paramonov, A.S.; et al. Multi-Enzymatic Cascades in the Synthesis of Modified Nucleosides: Comparison of the Thermophilic and Mesophilic Pathways. Biomolecules 2021, 11, 586. https://doi.org/10.3390/biom11040586
Fateev IV, Kostromina MA, Abramchik YA, Eletskaya BZ, Mikheeva OO, Lukoshin DD, Zayats EA, Berzina MY, Dorofeeva EV, Paramonov AS, et al. Multi-Enzymatic Cascades in the Synthesis of Modified Nucleosides: Comparison of the Thermophilic and Mesophilic Pathways. Biomolecules. 2021; 11(4):586. https://doi.org/10.3390/biom11040586
Chicago/Turabian StyleFateev, Ilja V., Maria A. Kostromina, Yuliya A. Abramchik, Barbara Z. Eletskaya, Olga O. Mikheeva, Dmitry D. Lukoshin, Evgeniy A. Zayats, Maria Ya. Berzina, Elena V. Dorofeeva, Alexander S. Paramonov, and et al. 2021. "Multi-Enzymatic Cascades in the Synthesis of Modified Nucleosides: Comparison of the Thermophilic and Mesophilic Pathways" Biomolecules 11, no. 4: 586. https://doi.org/10.3390/biom11040586
APA StyleFateev, I. V., Kostromina, M. A., Abramchik, Y. A., Eletskaya, B. Z., Mikheeva, O. O., Lukoshin, D. D., Zayats, E. A., Berzina, M. Y., Dorofeeva, E. V., Paramonov, A. S., Kayushin, A. L., Konstantinova, I. D., & Esipov, R. S. (2021). Multi-Enzymatic Cascades in the Synthesis of Modified Nucleosides: Comparison of the Thermophilic and Mesophilic Pathways. Biomolecules, 11(4), 586. https://doi.org/10.3390/biom11040586