
The Effect of a Unique Region of Parvovirus B19 Capsid Protein VP1 on Endothelial Cells

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. PTEC Characterization
2.3. Immunocytochemistry
2.4. Tube Forming Assay
2.5. Acetylated Low-Density Lipoprotein Uptake
2.6. VP1u Purification
2.7. Phospholipase A2 Catalytic Activity Assay
2.8. Internalization of Recombinant GFP-VP1u
2.9. Cell Viability Assay
2.10. Real-Time qPCR
2.11. Western Blot
2.12. Statistical Analysis
3. Results
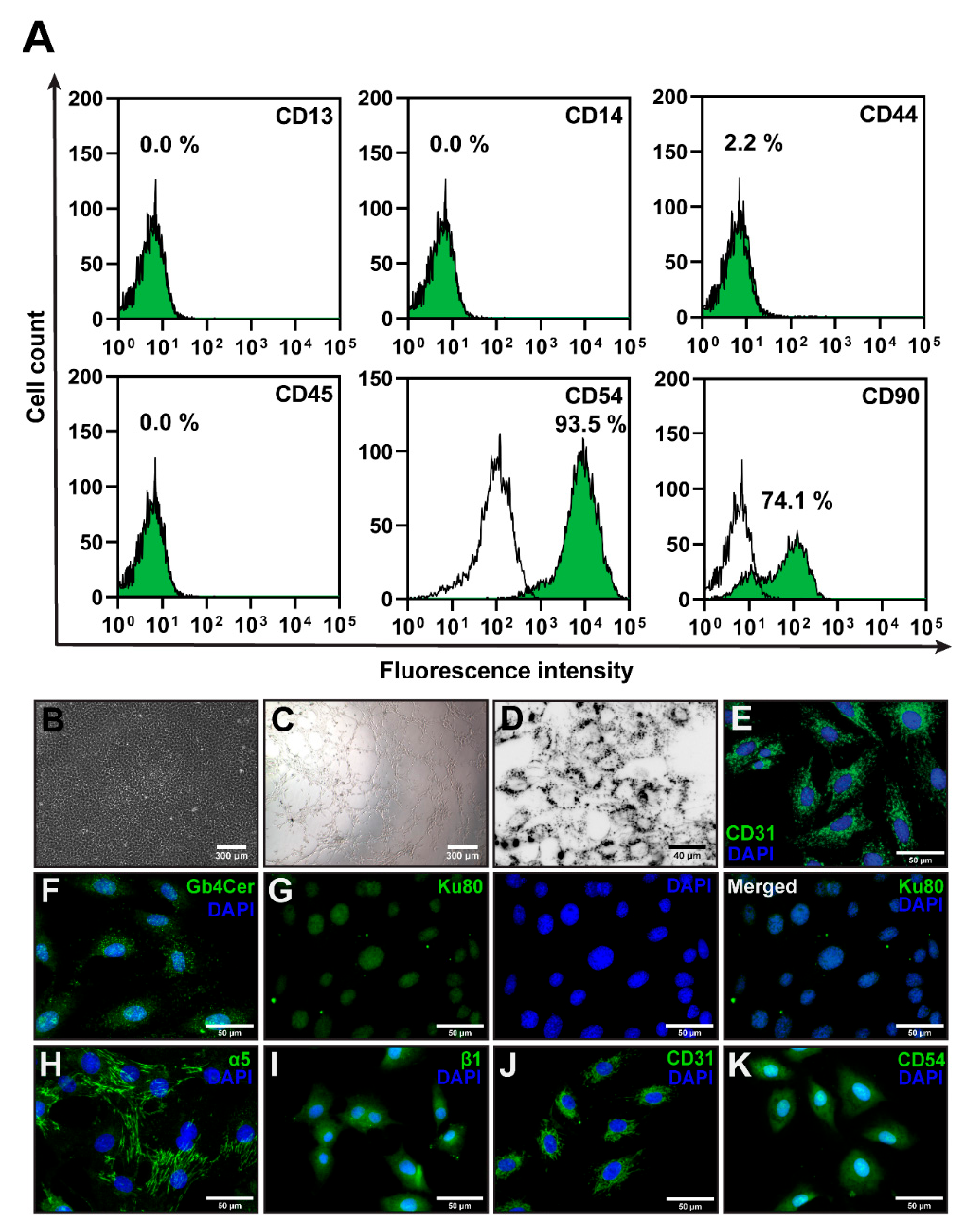
3.1. PTEC Isolation and Characterization
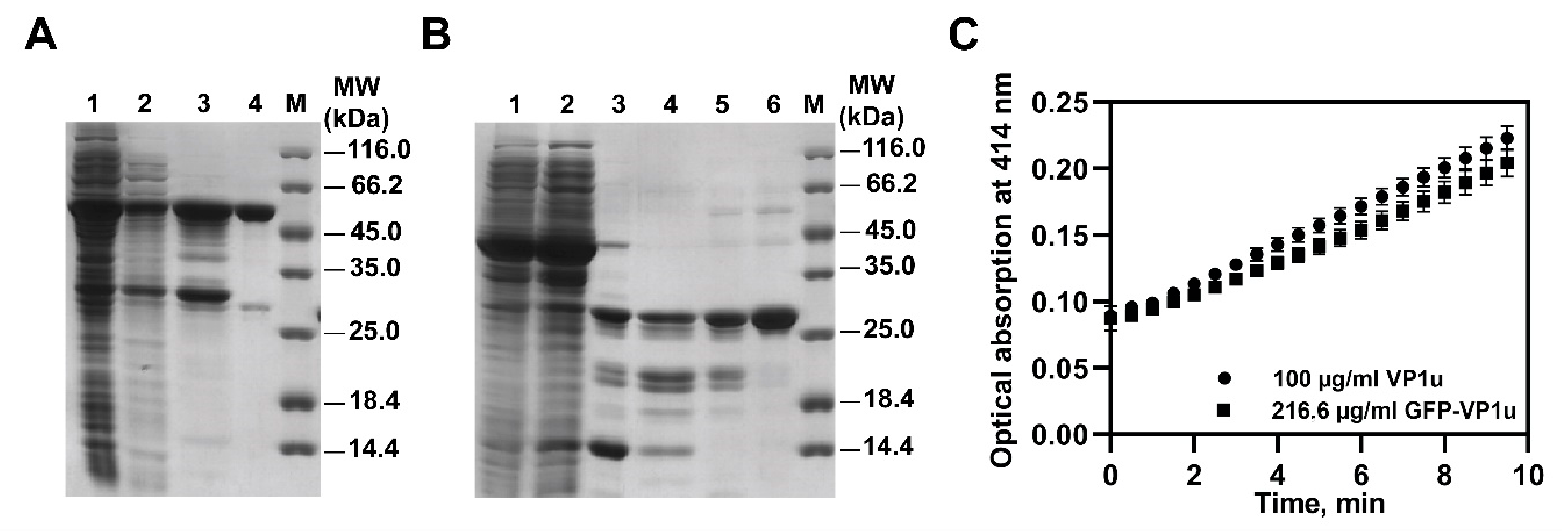
3.2. Evaluation of Recombinant VP1u
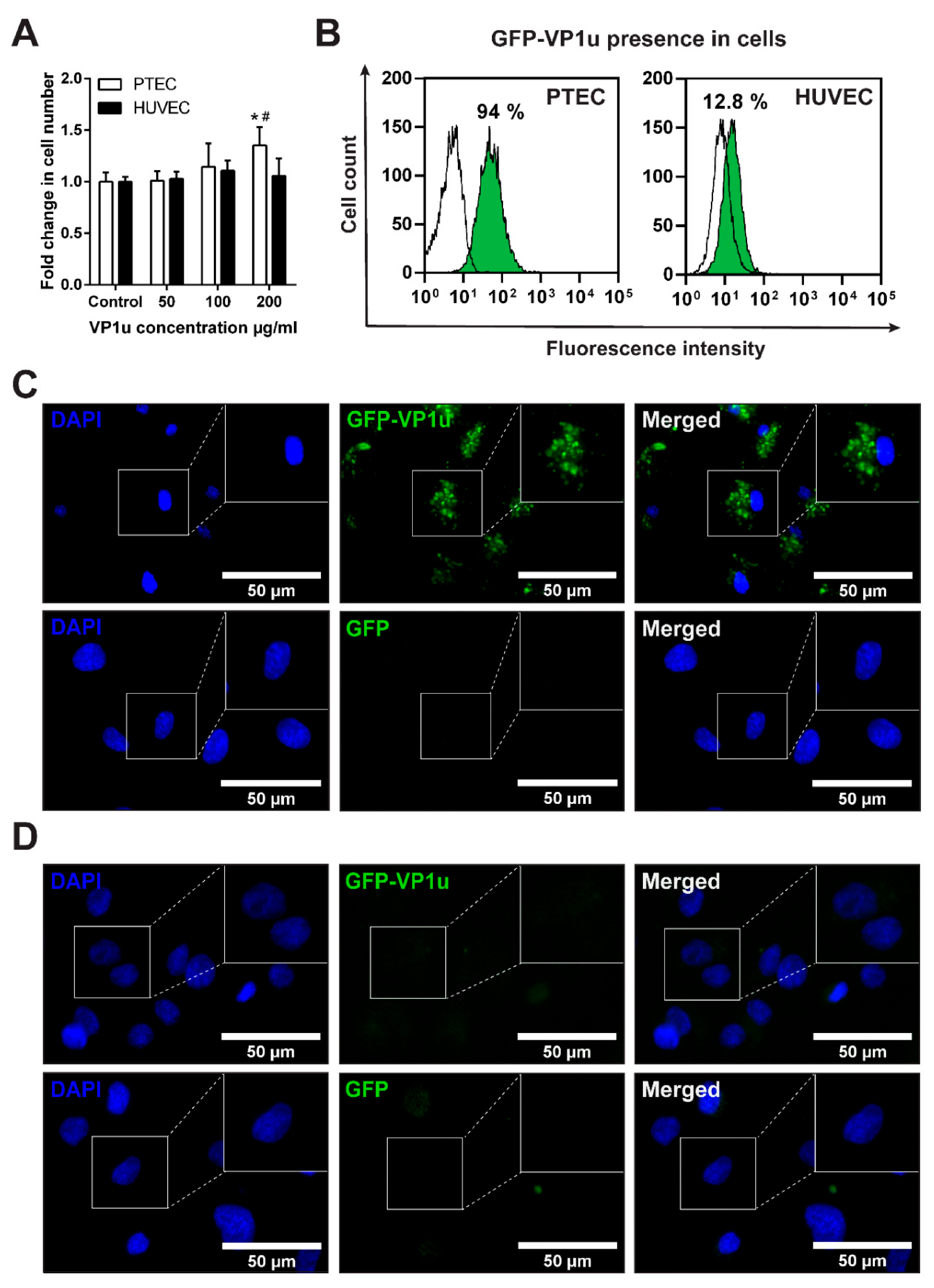
3.3. Cell Viability after VP1u Exposure
3.4. GFP-VP1u Uptake by the ECs
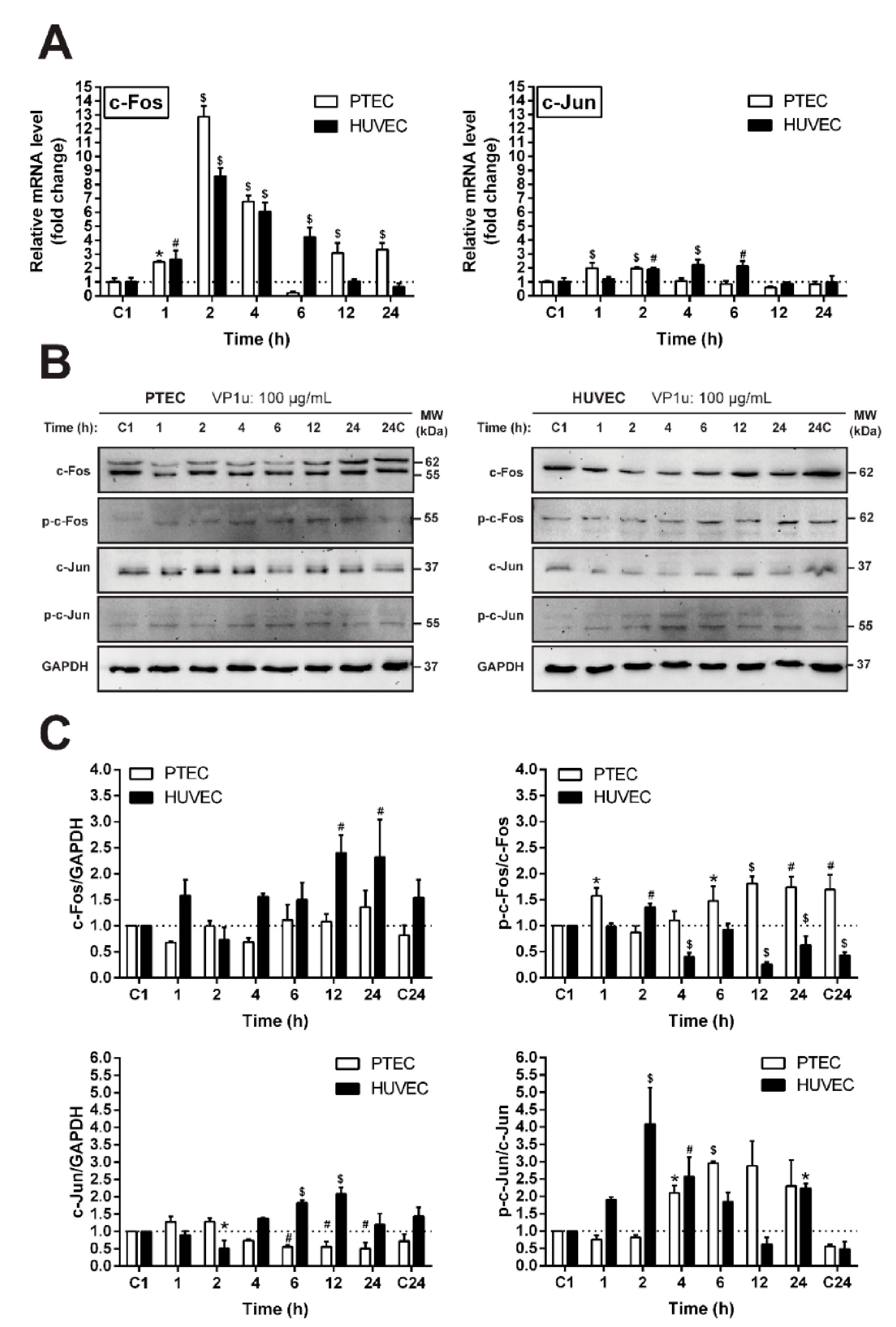
3.5. The Effect of VP1u on the Expression and Activation of c-Fos and c-Jun Proto-Oncogenes
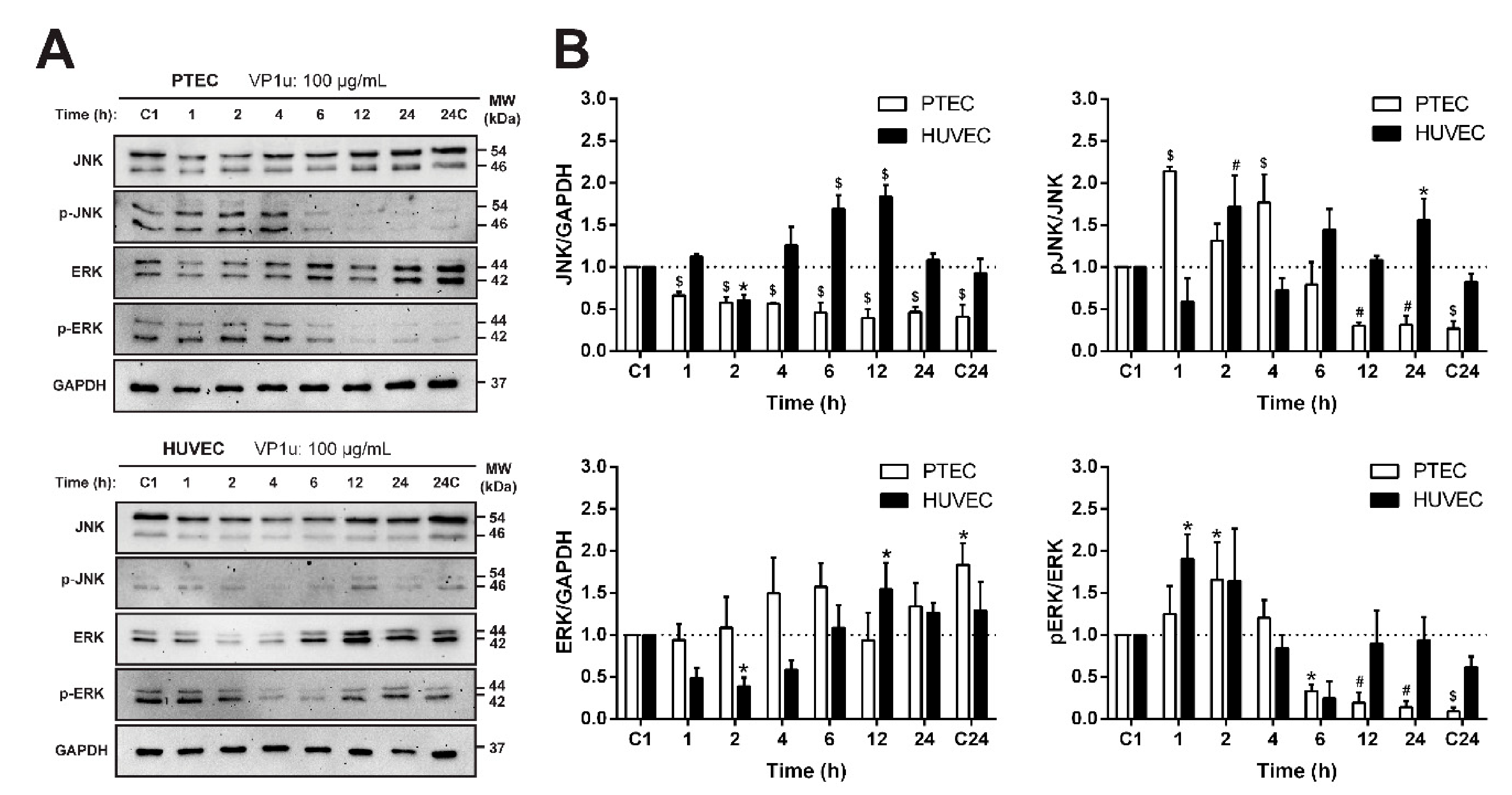
3.6. The Effect of VP1u on the Relative Abundance of Stress Response Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heegaard, E.D.; Brown, K.E. Human parvovirus B19. Clin. Microbiol. Rev. 2002, 15, 485–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufmann, B.; Simpson, A.A.; Rossmann, M.G. The structure of human parvovirus B19. Proc. Natl. Acad. Sci. USA 2004, 101, 11628–11633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.J.; Jones, S.E.; Fisher-Hoch, S.P.; Lewis, E.; Hall, S.M.; Bartlett, C.L.; Cohen, B.J.; Mortimer, P.P.; Pereira, M.S. Human parvovirus, the cause of erythema infectiosum (fifth disease)? Lancet 1983, 1, 1378. [Google Scholar] [CrossRef]
- Cohen, B.J.; Buckley, M.M. The prevalence of antibody to human parvovirus B 19 in England and Wales. J. Med. Microbiol. 1988, 25, 151–153. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, K.; Kurtzman, G.; Young, N. Replication of the B19 parvovirus in human bone marrow cell cultures. Science 1986, 233, 883–886. [Google Scholar] [CrossRef]
- Vassias, I.; Perol, S.; Coulombel, L.; Thebault, M.C.; Lagrange, P.H.; Morinet, F. An in situ hybridization technique for the study of B19 human parvovirus replication in bone marrow cell cultures. J. Virol. Methods 1993, 44, 329–338. [Google Scholar] [CrossRef]
- Young, N.; Harrison, M.; Moore, J.; Mortimer, P.; Humphries, R.K. Direct demonstration of the human parvovirus in erythroid progenitor cells infected in vitro. J. Clin. Investig. 1984, 74, 2024–2032. [Google Scholar] [CrossRef]
- Chen, A.Y.; Kleiboeker, S.; Qiu, J. Productive Parvovirus B19 Infection of Primary Human Erythroid Progenitor Cells at Hypoxia Is Regulated by STAT5A and MEK Signaling but not HIFα. PLoS Pathog. 2011, 7, e1002088. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.E.; Anderson, S.M.; Young, N.S. Erythrocyte P antigen: Cellular receptor for B19 parvovirus. Science 1993, 262, 114–117. [Google Scholar] [CrossRef]
- Chipman, P.R.; Agbandje-McKenna, M.; Kajigaya, S.; Brown, K.E.; Young, N.S.; Baker, T.S.; Rossmann, M.G. Cryo-electron microscopy studies of empty capsids of human parvovirus B19 complexed with its cellular receptor. Proc. Natl. Acad. Sci. USA 1996, 93, 7502–7506. [Google Scholar] [CrossRef] [Green Version]
- Munakata, Y.; Saito-Ito, T.; Kumura-Ishii, K.; Huang, J.; Kodera, T.; Ishii, T.; Hirabayashi, Y.; Koyanagi, Y.; Sasaki, T. Ku80 autoantigen as a cellular coreceptor for human parvovirus B19 infection. Blood 2005, 106, 3449–3456. [Google Scholar] [CrossRef]
- Weigel-Kelley, K.A.; Yoder, M.C.; Srivastava, A. Alpha5beta1 integrin as a cellular coreceptor for human parvovirus B19: Requirement of functional activation of beta1 integrin for viral entry. Blood 2003, 102, 3927–3933. [Google Scholar] [CrossRef]
- Leisi, R.; Ruprecht, N.; Kempf, C.; Ros, C. Parvovirus B19 Uptake Is a Highly Selective Process Controlled by VP1u, a Novel Determinant of Viral Tropism. J. Virol. 2013, 87, 13161–13167. [Google Scholar] [CrossRef] [Green Version]
- Leisi, R.; Di Tommaso, C.; Kempf, C.; Ros, C. The Receptor-Binding Domain in the VP1u Region of Parvovirus B19. Viruses 2016, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Leisi, R.; von Nordheim, M.; Kempf, C.; Ros, C. Specific Targeting of Proerythroblasts and Erythroleukemic Cells by the VP1u Region of Parvovirus B19. Bioconjug. Chem. 2015, 26, 1923–1930. [Google Scholar] [CrossRef]
- Pasquinelli, G.; Bonvicini, F.; Foroni, L.; Salfi, N.; Gallinella, G. Placental endothelial cells can be productively infected by Parvovirus B19. J. Clin. Virol. 2009, 44, 33–38. [Google Scholar] [CrossRef]
- Bultmann, B.D.; Klingel, K.; Sotlar, K.; Bock, C.T.; Baba, H.A.; Sauter, M.; Kandolf, R. Fatal parvovirus B19-associated myocarditis clinically mimicking ischemic heart disease: An endothelial cell-mediated disease. Hum. Pathol. 2003, 34, 92–95. [Google Scholar] [CrossRef]
- Bultmann, B.D.; Klingel, K.; Sotlar, K.; Bock, C.T.; Kandolf, R. Parvovirus B19: A pathogen responsible for more than hematologic disorders. Virchows. Arch. 2003, 442, 8–17. [Google Scholar] [CrossRef]
- Spartalis, M.; Tzatzaki, E.; Spartalis, E.; Damaskos, C.; Mavrogeni, S.; Voudris, V. Parvovirus B19 Myocarditis of Fulminant Evolution. Cardiol. Res. 2017, 8, 172–175. [Google Scholar] [CrossRef] [Green Version]
- Dyrsen, M.E.; Iwenofu, O.H.; Nuovo, G.; Magro, C.M. Parvovirus B19-associated catastrophic endothelialitis with a Degos-like presentation. J. Cutan. Pathol. 2008, 35 (Suppl. 1), 20–25. [Google Scholar] [CrossRef]
- Zakrzewska, K.; Cortivo, R.; Tonello, C.; Panfilo, S.; Abatangelo, G.; Giuggioli, D.; Ferri, C.; Corcioli, F.; Azzi, A. Human parvovirus B19 experimental infection in human fibroblasts and endothelial cells cultures. Virus Res. 2005, 114, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Von Kietzell, K.; Pozzuto, T.; Heilbronn, R.; Grössl, T.; Fechner, H.; Weger, S. Antibody-mediated enhancement of parvovirus B19 uptake into endothelial cells mediated by a receptor for complement factor C1q. J. Virol. 2014, 88, 8102–8115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozzuto, T.; von Kietzell, K.; Bock, T.; Schmidt-Lucke, C.; Poller, W.; Zobel, T.; Lassner, D.; Zeichhardt, H.; Weger, S.; Fechner, H. Transactivation of human parvovirus B19 gene expression in endothelial cells by adenoviral helper functions. Virology 2011, 411, 50–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalvelyte, A.; Krestnikova, N.; Stulpinas, A.; Bukelskiene, V.; Bironaite, D.; Baltriukiene, D.; Imbrasaite, A. Long-term muscle-derived cell culture: Multipotency and susceptibility to cell death stimuli. Cell Biol. Int. 2013, 37, 292–304. [Google Scholar] [CrossRef]
- Bogomolovas, J.; Simon, B.; Sattler, M.; Stier, G. Screening of fusion partners for high yield expression and purification of bioactive viscotoxins. Protein Expr. Purif. 2009, 64, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Azzi, A.; Zakrzewska, K.; Bertoni, E.; Guidi, S.; Salvadori, M. Persistent parvovirus B19 infections with different clinical outcomes in renal transplant recipients: Diagnostic relevance of polymerase chain reaction (PCR) and of quantification of B19 DNA in sera. Clin. Microbiol. Infect. 1996, 2, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Jordan, J.A. Diagnosing human parvovirus B19 infection: Guidelines for test selection. Mol. Diagn. 2001, 6, 307–312. [Google Scholar] [CrossRef]
- Hu, Y.; Cheng, L.; Hochleitner, B.W.; Xu, Q. Activation of mitogen-activated protein kinases (ERK/JNK) and AP-1 transcription factor in rat carotid arteries after balloon injury. Arter. Thromb. Vasc. Biol. 1997, 17, 2808–2816. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.; Zhi, N.; Filippone, C.; Keyvanfar, K.; Kajigaya, S.; Brown, K.E.; Young, N.S. Ex vivo-generated CD36+ erythroid progenitors are highly permissive to human parvovirus B19 replication. J. Virol. 2008, 82, 2470–2476. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.C.; Shi, Y.F.; Yang, J.J.; Hsiao, Y.C.; Tzang, B.S.; Hsu, T.C. Effects of Human Parvovirus B19 and Bocavirus VP1 Unique Region on Tight Junction of Human Airway Epithelial A549 Cells. PLoS ONE 2014, 9, e107970. [Google Scholar] [CrossRef]
- Bausinger, H.; Lipsker, D.; Ziylan, U.; Manie, S.; Briand, J.P.; Cazenave, J.P.; Muller, S.; Haeuw, J.F.; Ravanat, C.; de la Salle, H.; et al. Endotoxin-free heat-shock protein 70 fails to induce APC activation. Eur. J. Immunol. 2002, 32, 3708–3713. [Google Scholar] [CrossRef]
- Henderson, B.; Poole, S.; Wilson, M. Bacterial modulins: A novel class of virulence factors which cause host tissue pathology by inducing cytokine synthesis. Microbiol. Rev. 1996, 60, 316. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.F.; Dong, Y.M.; Yi, Q.H.; Huang, Y.; Zhao, D.; Yang, Y.B.; Tijssen, P.; Qiu, J.M.; Liu, K.Y.; Li, Y. The Determinants for the Enzyme Activity of Human Parvovirus B19 Phospholipase A2 (PLA2) and Its Influence on Cultured Cells. PLoS ONE 2013, 8, e61440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzang, B.S.; Chiu, C.C.; Tsai, C.C.; Lee, Y.J.; Lu, I.J.; Shi, J.Y.; Hsu, T.C. Effects of human parvovirus B19 VP1 unique region protein on macrophage responses. J. Biomed. Sci. 2009, 16, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, S.; de Fougerolles, A.R.; Blaikie, P.; Khan, L.; Pepe, A.; Green, C.D.; Koteliansky, V.; Giancotti, F.G. α5β1 Integrin Activates an NF-κB-Dependent Program of Gene Expression Important for Angiogenesis and Inflammation. Mol. Cell. Biol. 2002, 22, 5912–5922. [Google Scholar] [CrossRef] [Green Version]
- Weigel-Kelley, K.A.; Yoder, M.C.; Srivastava, A. Recombinant human parvovirus B19 vectors: Erythrocyte P antigen is necessary but not sufficient for successful transduction of human hematopoietic cells. J. Virol. 2001, 75, 4110–4116. [Google Scholar] [CrossRef] [Green Version]
- Gomez, D.; Reich, N.C. Stimulation of primary human endothelial cell proliferation by IFN. J. Immunol. 2003, 170, 5373–5381. [Google Scholar] [CrossRef] [Green Version]
- Maloney, J.P.; Gao, L. Proinflammatory Cytokines Increase Vascular Endothelial Growth Factor Expression in Alveolar Epithelial Cells. Mediat. Inflamm. 2015. [Google Scholar] [CrossRef] [Green Version]
- Hess, J.; Angel, P.; Schorpp-Kistner, M. AP-1 subunits: Quarrel and harmony among siblings. J. Cell Sci. 2004, 117, 5965–5973. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Ishii, K.K.; Munakata, Y.; Saitoh, T.; Kaku, M.; Sasaki, T. Regulation of Tumor Necrosis Factor Alpha Promoter by Human Parvovirus B19 NS1 through Activation of AP-1 and AP-2. J. Virol. 2002, 76, 5395–5403. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yin, S.; Chen, Y.; Wu, Y.; Zheng, W.; Dong, H.; Bai, Y.; Qin, Y.; Li, J.; Feng, S.; et al. LPS-induced proinflammatory cytokine expression in human airway epithelial cells and macrophages via NF-κB, STAT3 or AP-1 activation. Mol. Med. Rep. 2018, 17, 5484–5491. [Google Scholar] [CrossRef] [Green Version]
- Hogan, N.T.; Whalen, M.B.; Stolze, L.K.; Hadeli, N.K.; Lam, M.T.; Springstead, J.R.; Glass, C.K.; Romanoski, C.E. Transcriptional networks specifying homeostatic and inflammatory programs of gene expression in human aortic endothelial cells. eLife 2017, 6, e22536. [Google Scholar] [CrossRef]
- Wang, N.; Verna, L.; Hardy, S.; Forsayeth, J.; Zhu, Y.; Stemerman, M.B. Adenovirus-mediated overexpression of c-Jun and c-Fos induces intercellular adhesion molecule-1 and monocyte chemoattractant protein-1 in human endothelial cells. Arter. Thromb. Vasc. Biol. 1999, 19, 2078–2084. [Google Scholar] [CrossRef] [Green Version]
- Tung, W.H.; Hsieh, H.L.; Yang, C.M. Enterovirus 71 induces COX-2 expression via MAPKs, NF-kappa B, and AP-1 in SK-N-SH cells: Role of PGE(2) in viral replication. Cell. Signal. 2010, 22, 234–246. [Google Scholar] [CrossRef]
- Gustems, M.; Woellmer, A.; Rothbauer, U.; Eck, S.H.; Wieland, T.; Lutter, D.; Hammerschmidt, W. c-Jun/c-Fos heterodimers regulate cellular genes via a newly identified class of methylated DNA sequence motifs. Nucleic Acids Res. 2014, 42, 3059–3072. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, M.; Zhang, X.; Chu, F.; Zhou, T. MAPK/c-Jun signaling pathway contributes to the upregulation of the anti-apoptotic proteins Bcl-2 and Bcl-xL induced by Epstein-Barr virus-encoded BARF1 in gastric carcinoma cells. Oncol. Lett. 2018, 15, 7537–7544. [Google Scholar] [CrossRef] [Green Version]
- Zádori, Z.; Szelei, J.; Lacoste, M.C.; Li, Y.; Gariépy, S.; Raymond, P.; Allaire, M.; Nabi, I.R.; Tijssen, P. A viral phospholipase A2 is required for parvovirus infectivity. Dev. Cell 2001, 1, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Dorsch, S.; Liebisch, G.; Kaufmann, B.; von Landenberg, P.; Hoffmann, J.H.; Drobnik, W.; Modrow, S. The VP1 unique region of parvovirus B19 and its constituent phospholipase A2-like activity. J. Virol. 2002, 76, 2014–2018. [Google Scholar] [CrossRef] [Green Version]
- Tokumoto, H.; Croxtall, J.D.; Choudhury, Q.; Flower, R.J. Phospholipase A2-induced stimulation of A549 lung adenocarcinoma cell line proliferation. Biochim. Biophys. Acta 1993, 1169, 236–242. [Google Scholar] [CrossRef]
- Arita, H.; Hanasaki, K.; Nakano, T.; Oka, S.; Teraoka, H.; Matsumoto, K. Novel proliferative effect of phospholipase A2 in Swiss 3T3 cells via specific binding site. J. Biol. Chem. 1991, 266, 19139–19141. [Google Scholar] [CrossRef]
- Rodriguez De Turco, E.B.; Jackson, F.R.; DeCoster, M.A.; Kolko, M.; Bazan, N.G. Glutamate signalling and secretory phospholipase A2 modulate the release of arachidonic acid from neuronal membranes. J. Neurosci. Res. 2002, 68, 558–567. [Google Scholar] [CrossRef]
- Bernstein, D.I.; El Sahly, H.M.; Keitel, W.A.; Wolff, M.; Simone, G.; Segawa, C.; Wong, S.; Shelly, D.; Young, N.S.; Dempsey, W. Safety and immunogenicity of a candidate parvovirus B19 vaccine. Vaccine 2011, 29, 7357–7363. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rinkūnaitė, I.; Šimoliūnas, E.; Bironaitė, D.; Rutkienė, R.; Bukelskienė, V.; Meškys, R.; Bogomolovas, J. The Effect of a Unique Region of Parvovirus B19 Capsid Protein VP1 on Endothelial Cells. Biomolecules 2021, 11, 606. https://doi.org/10.3390/biom11040606
Rinkūnaitė I, Šimoliūnas E, Bironaitė D, Rutkienė R, Bukelskienė V, Meškys R, Bogomolovas J. The Effect of a Unique Region of Parvovirus B19 Capsid Protein VP1 on Endothelial Cells. Biomolecules. 2021; 11(4):606. https://doi.org/10.3390/biom11040606
Chicago/Turabian StyleRinkūnaitė, Ieva, Egidijus Šimoliūnas, Daiva Bironaitė, Rasa Rutkienė, Virginija Bukelskienė, Rolandas Meškys, and Julius Bogomolovas. 2021. "The Effect of a Unique Region of Parvovirus B19 Capsid Protein VP1 on Endothelial Cells" Biomolecules 11, no. 4: 606. https://doi.org/10.3390/biom11040606
APA StyleRinkūnaitė, I., Šimoliūnas, E., Bironaitė, D., Rutkienė, R., Bukelskienė, V., Meškys, R., & Bogomolovas, J. (2021). The Effect of a Unique Region of Parvovirus B19 Capsid Protein VP1 on Endothelial Cells. Biomolecules, 11(4), 606. https://doi.org/10.3390/biom11040606