
Antimicrobial Activity of Cyclic-Monomeric and Dimeric Derivatives of the Snail-Derived Peptide Cm-p5 against Viral and Multidrug-Resistant Bacterial Strains

 , , ,
, , ,  , ,
, ,  and add
Show full author list
and add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Mammalian Cell Lines and Culture Conditions
2.3. Peptide Synthesis
2.4. Antibacterial Activity
2.4.1. Agar Overlay Assay
2.4.2. Minimal Inhibitory Concentration (MIC) Determinations
2.5. H-Uracil Proliferation Assay
2.6. Effect of Peptides on Zika Virus Infection
2.7. Effect of Peptides on HSV Infection
2.8. Effect of Peptides on SARS-CoV-2 Pseudo Particles
2.8.1. Pseudotyping of VSV with SARS-CoV-2S
2.8.2. Transduction Experiments with SARS-CoV-2 Spike-Pseudoparticles
2.8.3. Cell Viability Assay
3. Results
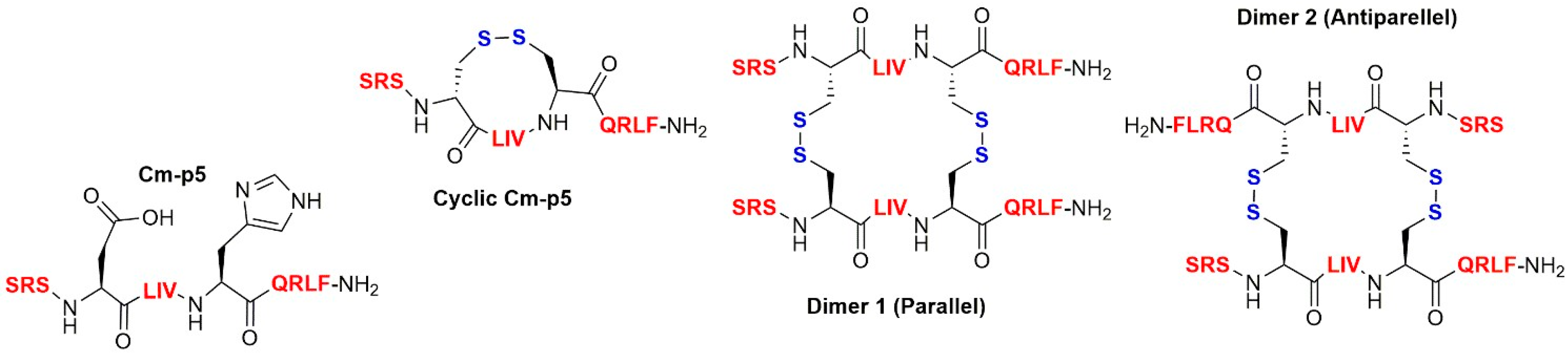
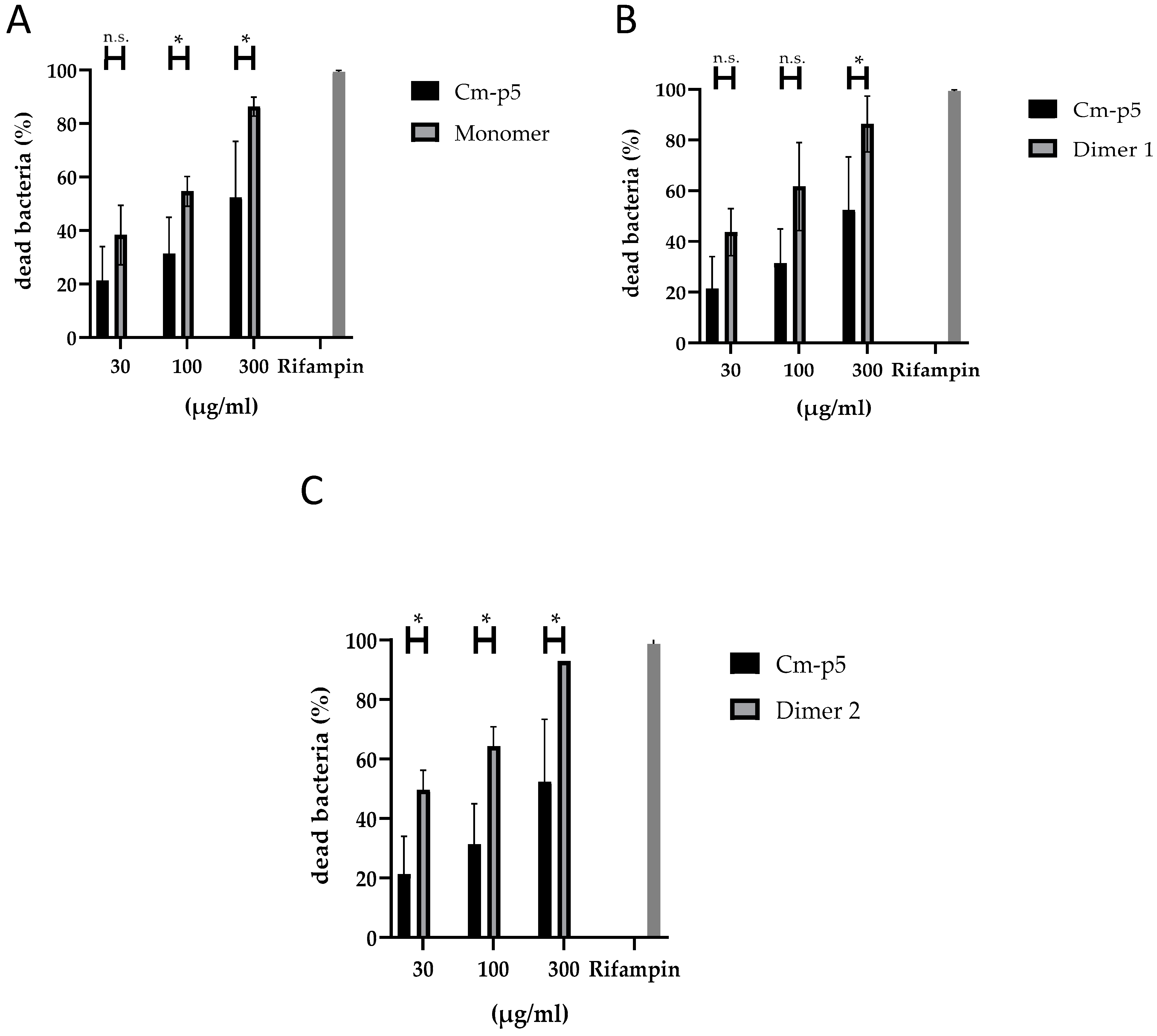
3.1. Antibacterial Activity
3.2. Antimycobacterial Activity
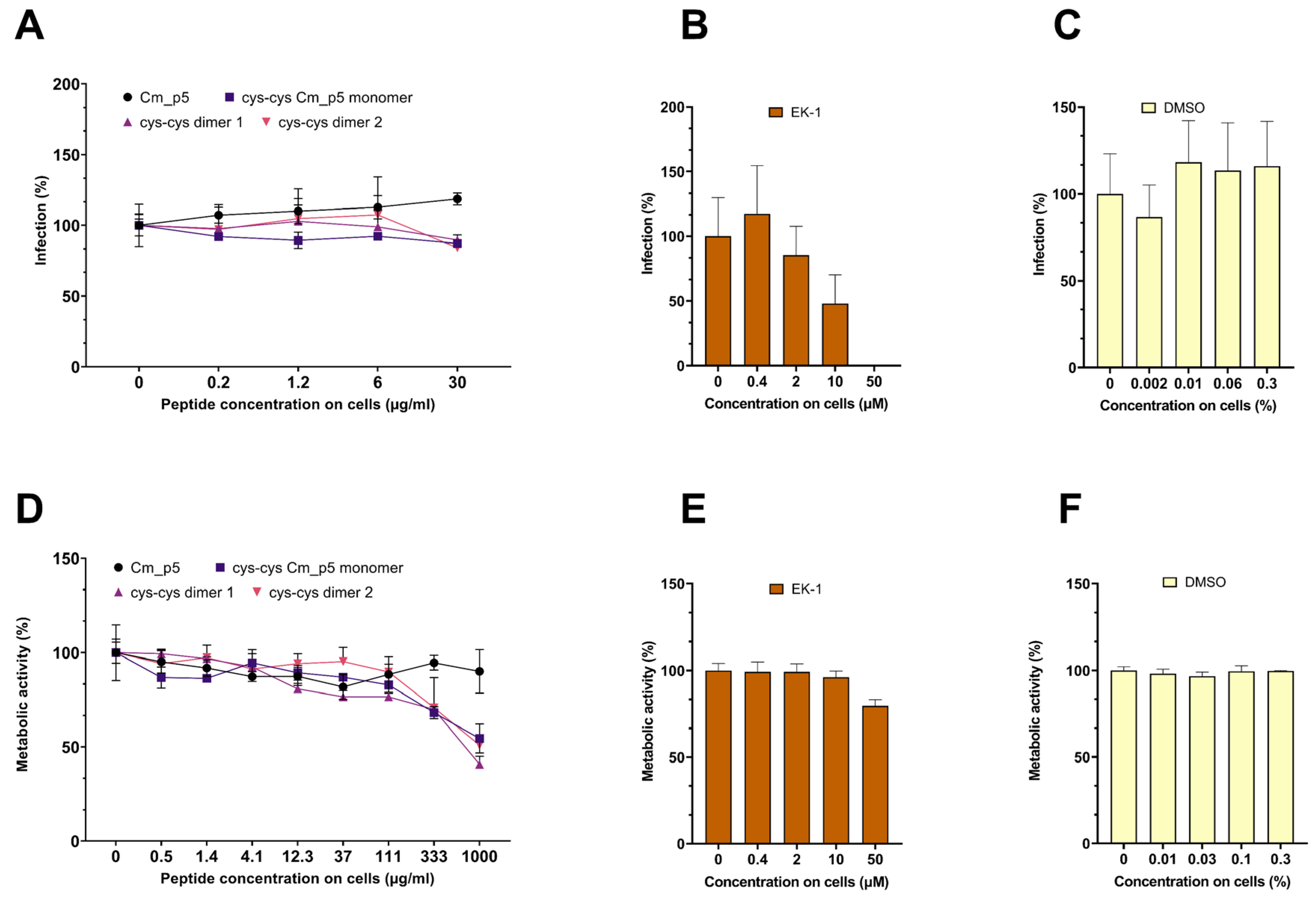
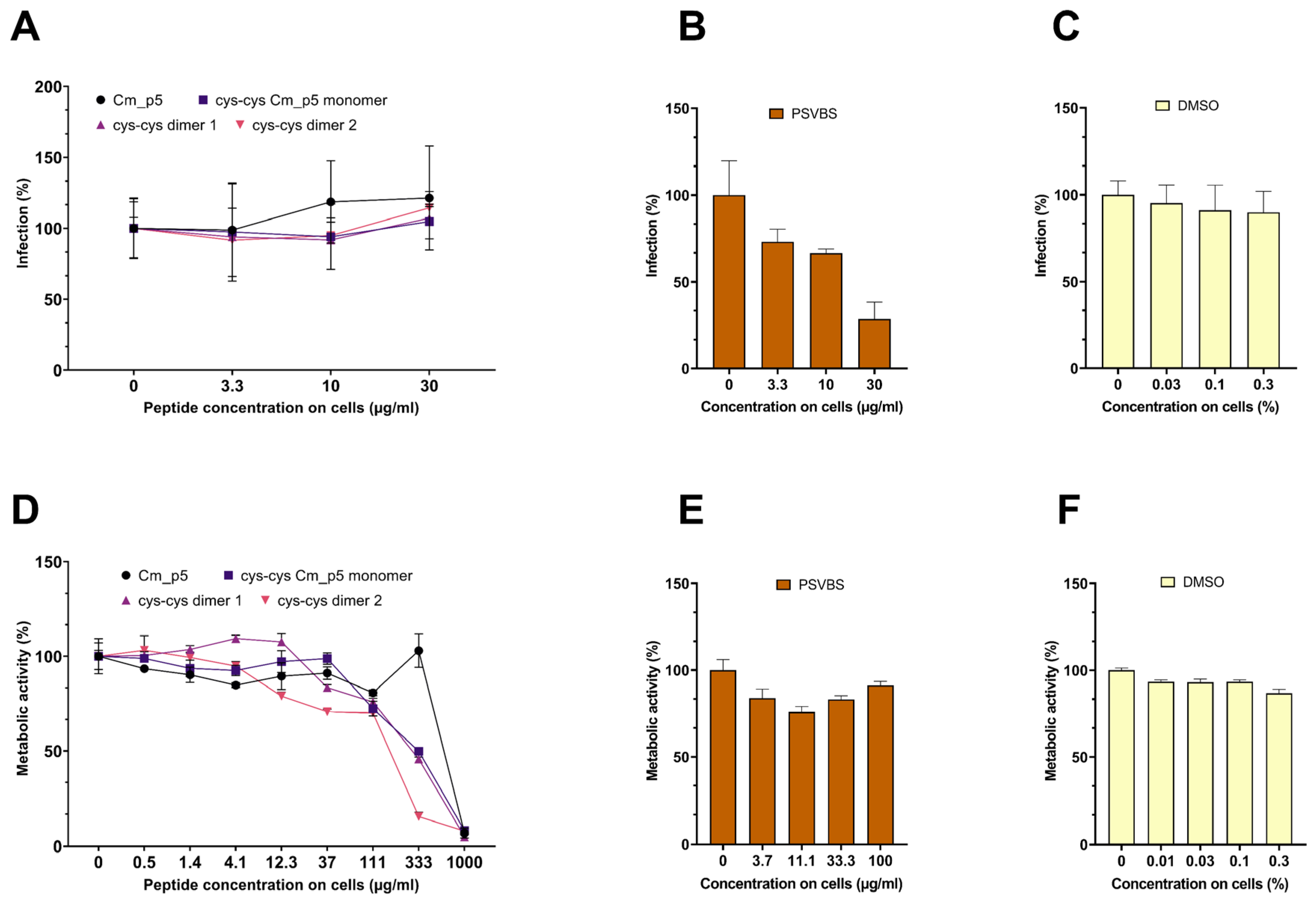
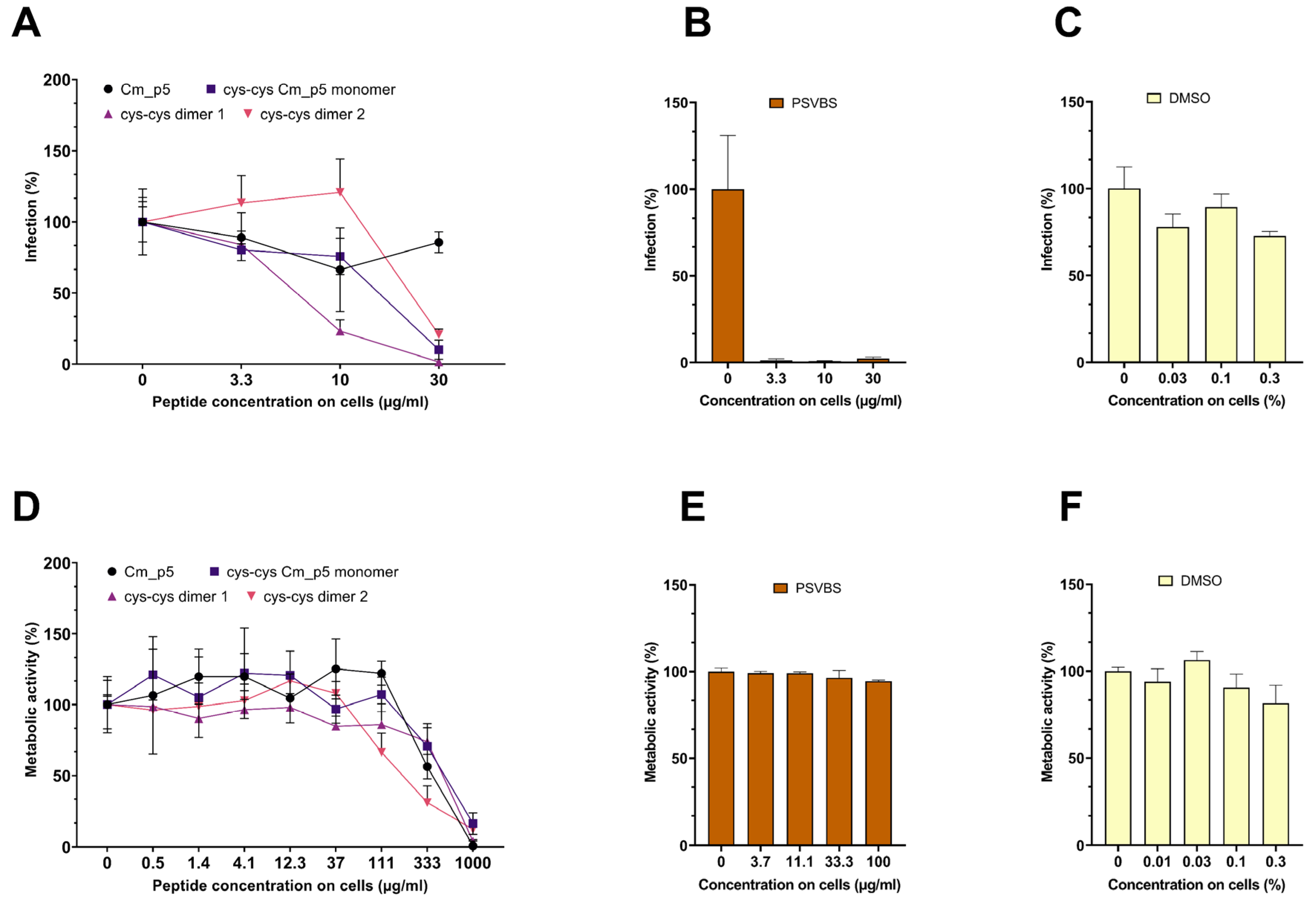
3.3. Antiviral Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- López-Abarrategui, C.; McBeth, M.; Mandal Santi, J.; Sun Zhenyu, G.; Heffron, A.; Alba-Menéndez, L.; Migliolo, O.; Reyes-Acosta, M.; García-Villarino, D.O.; Nolasco, R.; et al. Cm-p5: An antifungal hydrophilic peptide derived from the coastal mollusk Cenchritis muricatus (Gastropoda: Littorinidae). FASEB J. 2015, 29, 3315–3325. [Google Scholar] [CrossRef] [Green Version]
- González-García, M.; Valdés, M.; Freitas, C.G.; Alba Menéndez, A.; López-Abarrategui, C.; San Juan Galán, J.; Campos Diaz, S.; Luiz Franco, O.; Otero-González, A. In vitro complementary biological activity of the antifungal peptide Cm-p5 and in silico prediction of its functional regions. Rev. Cuba. Med. Trop. 2017, 69, 1–15. [Google Scholar]
- Vicente, F.E.M.; González-Garcia, M.; Diaz Pico, E.; Moreno-Castillo, E.; Garay, H.E.; Rosi, P.E.; Jimenez, A.M.; Campos-Delgado, J.A.; Rivera, D.G.; Chinea, G.; et al. Design of a Helical-Stabilized, Cyclic, and Nontoxic Analogue of the Peptide Cm-p5 with Improved Antifungal Activity. ACS Omega 2019, 4, 19081−19095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Kizhakkedathu, J.; Straus, S. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Cusimano, M.; Spinello, A.; Barone, G.; Schillaci, D.; Cascioferro, S.; Magistrato, A.; Parrino, B.; Arizza, V.; Vitale, M. A Synthetic Derivative of Antimicrobial Peptide Holothuroidin 2 from Mediterranean Sea Cucumber (Holothuria tubulosa) in the Control of Listeria monocytogenes. Mar. Drugs 2019, 17, 159. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.; Sahl, H.G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Otero-González, A.J.; Magalhaes, B.S.; García-Villarino, M.C.; López-Abarrategui, C.; Sousa, D.A.; Dias, S.C. Antimicrobial peptides from marine invertebrates as a new frontier for microbial infection control. FASEB J. 2010, 24, 1320–1334. [Google Scholar] [CrossRef]
- Steinstraesser, L.; Kraneburg, U.; Jacobsen, F.; Al-Benna, S. Host defense peptides and their antimicrobial-immunomodulatory duality. Immunobiology 2011, 216, 322–333. [Google Scholar] [CrossRef]
- Shurko, J.F.; Galega, R.S.; Li, C.; Lee, G.C. Evaluation of LL-37 antimicrobial peptide derivatives alone and in combination with vancomycin against S. aureus. J Antibiot. 2018, 71, 971–974. [Google Scholar] [CrossRef] [Green Version]
- Graham, F.L.; Smiley, J.; Russell, W.C.; Nairn, R. Characteristics of a human cell line transformed by DNA from human adenovirus type 5. J. Gen. Virol. 1977, 36, 59–74. [Google Scholar] [CrossRef]
- Proffitt, M.R.; Schindler, S.A. Rapid detection of HSV with an enzyme linked virus inducible system (ELVIS) employing a genetically modified cell line. Clin. Diagn. Virol. 1995, 4, 175–182. [Google Scholar] [CrossRef]
- CLSI. Instruction for susceptibility assay for bacteria. In Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika virus. I. Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Schandock, F.; Riber, C.F.; Röcker, A. Macromolecular Antiviral Agents against Zika, Ebola, SARS, and Other Pathogenic Viruses. Adv. Healthc. Mater. 2017, 6, 1700748. [Google Scholar] [CrossRef] [Green Version]
- Aubry, M.; Richard, V.; Green, J.; Broult, J.; Musso, D. Inactivation of Zika virus in plasma with amotosalen and ultraviolet A illumination. Transfusion 2016, 56, 33–40. [Google Scholar] [CrossRef]
- Müller, J.A.; Harms, M.; Krüger, F. Semen inhibits Zika virus infection of cells and tissues from the anogenital region. Nat. Commun. 2018, 9, 2207. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Berger Rentsch, M.; Zimmer, G. A vesicular stomatitis virus replicon-based bioassay for the rapid and sensitive determination of multi-species type I interferon. PLoS ONE 2011, 6, e25858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, S.; Zhu, Y.; Liu, M. Fusion mechanism of 2019-nCoV and fusion inhibitors targeting HR1 domain in spike protein. Cell. Mol. Immunol. 2020, 17, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Liu, M.; Wang, C. Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. Cell Res. 2020, 30, 343–355. [Google Scholar] [CrossRef] [Green Version]
- Baindara, P.; Ghosh, A.K.; Mandal, S.M. Coevolution of Resistance against Antimicrobial Peptides. Microb. Drug Resist. 2020, 26, 880–899. [Google Scholar] [CrossRef] [PubMed]
- Pandit, G.; Biswas, K.; Ghosh, S.; Debnath, S.; Bidkar, A.P.; Satpati, P.; Bhunia, A.; Chatterjee, S. Rationally designed antimicrobial peptides: Insight into the mechanism of eleven residue peptides against microbial infections. Biomembranes 2020, 1862, 183177. [Google Scholar] [CrossRef] [PubMed]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging Strategies to Combat ESKAPE Pathogens in the Era of Antimicrobial Resistance: A Review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef]
- Girometti, N.; Lewis, R.E.; Giannella, M.; Ambretti, S.; Bartoletti, M.; Tedeschi, S.; Tumietto, F.; Cristini, F.; Trapani, F.; Gaibani, P.; et al. Klebsiella pneumoniae Bloodstream Infection: Epidemiology and Impact of Inappropriate Empirical Therapy. Medicine 2014, 93, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Meade, E.; Slattery, M.A.; Garvey, M. Bacteriocins, Potent Antimicrobial Peptides and the Fight against Multi Drug Resistant Species: Resistance Is utile? Antibiotics 2020, 9, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geitani, R.; Moubareck, C.A.; Touqui, L.; Sarkis, D.K. Cationic antimicrobial peptides: Alternatives and/or adjuvants to antibiotics active against methicillin-resistant Staphylococcus aureus and multidrug-resistant Pseudomonas aeruginosa. BMC Microbiol. 2019, 19, 54. [Google Scholar] [CrossRef] [Green Version]
- Mikkelsen, H.; McMullan, R.; Filloux, A. The Pseudomonas aeruginosa reference strain PA14 displays increased virulence due to a mutation in ladS. PLoS ONE 2011, 6, e29113. [Google Scholar] [CrossRef] [Green Version]
- Bolukaoto, J.Y.; Monyama, C.M.; Chukwu, M.O.; Lekala, S.M.; Nchabeleng, M.; Maloba, M.R.B.; Mavenyengwa, R.T.; Lebelo, S.L.; Monokoane, S.T.; Tshepuwane, C.; et al. Antibiotic resistance of Streptococcus agalactiae isolated from pregnant women in Garankuwa, South Africa. BMC Res. Notes 2015, 8, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaramillo-Jaramillo, A.S.; Cobo-Ángel, C.G.; Moreno-Tolosa, Y.; Ceballos-Márquez, A. Resistencia antimicrobiana de Streptococcus agalactiae de origen humano y bovino. Rev. CES Med. Zootec. 2018, 13, 62–79. [Google Scholar] [CrossRef]
- López-Abarrategui, C.; Alba, A.; Silva, O.; Reyes-Acosta, I.M.; Vasconcelos, J.T.A.; Olivera, L.; Migliolo, M.P.; Costa, C.R.; Costa, M.R.R.; Silva, H.E.; et al. Functional characterization of a synthetic hydrophilic antifungal peptide dericed from the marine snail Cenchritis muricatus. Biochimie 2012, 94, 968–974. [Google Scholar] [CrossRef] [Green Version]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Leszczynska, K.; Namiot, D.; Byfield, F.J.; Cruz, K.; Zendzian-Piotrowska, M.; Fein, D.E.; Savage, P.B.; Diamond, S.; McCulloch, C.A.; Janmey, P.A.; et al. Antibacterial activity of the human host defence peptide LL-37 and selected synthetic cationic lipids against bacteria associated with oral and upper respiratory tract infections. J. Antimicrob. Chemother. 2013, 68, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Borrero, R.; Álvarez, N.; Reyes, F.; Sarmiento, M.E.; Acosta, A. Mycobacterium tuberculosis: Factores de virulencia. Vacci Monit. 2011, 20, 34–38. [Google Scholar]
- AlMatar, M.; Makky, E.A.; Yakici, G.; Var, I.; Kayar, B.; Koksal, F. Antimicrobial peptides as an alternative to anti-tuberculosis drugs. Pharmacol. Res. 2017, 128, 288–305. [Google Scholar] [CrossRef]
- Carroll, J.; Field, D.; O’Connor, P.M.; Cotter, P.D.; Coffey, A.; Hill, C. Gene encoded antimicrobial peptides, a template for the design of novel anti-mycobacterial drugs. Bioeng. Bugs 2010, 1, 408–412. [Google Scholar] [CrossRef] [Green Version]
- Corrales-Garcia, L.; Ortiz, E.; Castañeda-Delgado, J.; Rivas-Santiago, B.; Corzo, G. Bacterial expression and antibiotic activities of recombinant variants of human beta-defensins on pathogenic bacteria and M. tuberulosis. Protein Expr. Purif. 2013, 89, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Sala, A.; Ardizzoni, A.; Ciociola, T.; Magliani, W.; Conti, S.; Blasi, E.; Cermelli, C. Antiviral Activity of Synthetic Peptides Derived from Physiological Proteins. Intervirology 2018, 61, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Góngora-Benítez, M.; Tulla-Puche, J.; Albericio, F. Multifaceted Roles of Disulfide Bonds. Peptides as Therapeutics. Chem. Rev. 2014, 114, 901–926. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration | CMP5 cys-cys Monomer | CMP5 cys-cys Dimer1 | CMP5 cys-cys Dimer2 | LL-37 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Bacterial Species | 1000 mg/L | 100 mg/L | 20 mg/L | 1000 mg/L | 100 mg/L | 20 mg/L | 1000 mg/L | 100 mg/L | 20 mg/L | 1 mg/L | |
| Streptococcus agalactiae | 0.8 | 0.4 | - | 1 | 0.7 | - | 1.1 | 0.6 | 0.3 | n.d. | |
| Staphylococcus aureus MRSA | - | - | - | - | - | - | - | - | - | 0.1 | |
| Listeria monocytogenes | 0.9 | 0.5 | - | 1.2 | 0.7 | 0.3 | 1.2 | 0.8 | 0.3 | 1.1 | |
| Klebsiella pneumoniae ESBL | 0.6 | - | - | 0.7 | 0.5 | - | 0.9 | 0.5 | - | 0.8 | |
| Pseudomonas aeruginosa | 0.8 | 0.4 | - | 1 | 0.7 | 0.3 | 1 | 0.6 | 0.3 | 0.8 | |
| Acinetobacter baumanii ATCC 19606 | 0.7 | 0.4 | - | 1 | 0.6 | 0.4 | 0.9 | 0.6 | 0.4 | 0.7 | |
| Enterococcus faecium VRE DSM 17050 | 0.9 | 0.6 | 0.4 | 1.1 | 0.8 | 0.5 | 1.1 | 0.8 | 0.5 | 1.0 | |
| MIC Values | AMP Monomer (MIC) | AMP Dimer 1 (MIC) | AMP Dimer 2 (MIC) | |
|---|---|---|---|---|
| Bacterial Species | ||||
| Streptococcus agalactiae | >100 mg/L | >100 mg/L | >100 mg/L | |
| Listeria monocytogenes | 100 mg/L | 50 mg/L | 12.5 mg/L | |
| Pseudomonas aeruginosa | >100 mg/L | >100 mg/L | >100 mg/L | |
| Klebsiella pneumoniae ESBL | >100 mg/L | >100 mg/L | >100 mg/L | |
| Acinetobacter baumanii | 100 mg/L | 50 mg/L | 25 mg/L | |
| Enterococcus faecium VRE | 50 mg/L | 50 mg/L | 25 mg/L | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-García, M.; Morales-Vicente, F.; Pico, E.D.; Garay, H.; Rivera, D.G.; Grieshober, M.; Raluca Olari, L.; Groß, R.; Conzelmann, C.; Krüger, F.; et al. Antimicrobial Activity of Cyclic-Monomeric and Dimeric Derivatives of the Snail-Derived Peptide Cm-p5 against Viral and Multidrug-Resistant Bacterial Strains. Biomolecules 2021, 11, 745. https://doi.org/10.3390/biom11050745
González-García M, Morales-Vicente F, Pico ED, Garay H, Rivera DG, Grieshober M, Raluca Olari L, Groß R, Conzelmann C, Krüger F, et al. Antimicrobial Activity of Cyclic-Monomeric and Dimeric Derivatives of the Snail-Derived Peptide Cm-p5 against Viral and Multidrug-Resistant Bacterial Strains. Biomolecules. 2021; 11(5):745. https://doi.org/10.3390/biom11050745
Chicago/Turabian StyleGonzález-García, Melaine, Fidel Morales-Vicente, Erbio Díaz Pico, Hilda Garay, Daniel G. Rivera, Mark Grieshober, Lia Raluca Olari, Rüdiger Groß, Carina Conzelmann, Franziska Krüger, and et al. 2021. "Antimicrobial Activity of Cyclic-Monomeric and Dimeric Derivatives of the Snail-Derived Peptide Cm-p5 against Viral and Multidrug-Resistant Bacterial Strains" Biomolecules 11, no. 5: 745. https://doi.org/10.3390/biom11050745
APA StyleGonzález-García, M., Morales-Vicente, F., Pico, E. D., Garay, H., Rivera, D. G., Grieshober, M., Raluca Olari, L., Groß, R., Conzelmann, C., Krüger, F., Zech, F., Prelli Bozzo, C., Müller, J. A., Zelikin, A., Raber, H., Kubiczek, D., Rosenau, F., Münch, J., Stenger, S., ... Otero-Gonzalez, A. J. (2021). Antimicrobial Activity of Cyclic-Monomeric and Dimeric Derivatives of the Snail-Derived Peptide Cm-p5 against Viral and Multidrug-Resistant Bacterial Strains. Biomolecules, 11(5), 745. https://doi.org/10.3390/biom11050745