
Non-Tumor CCAAT/Enhancer-Binding Protein Delta Potentiates Tumor Cell Extravasation and Pancreatic Cancer Metastasis Formation

 , , , and
, , , and 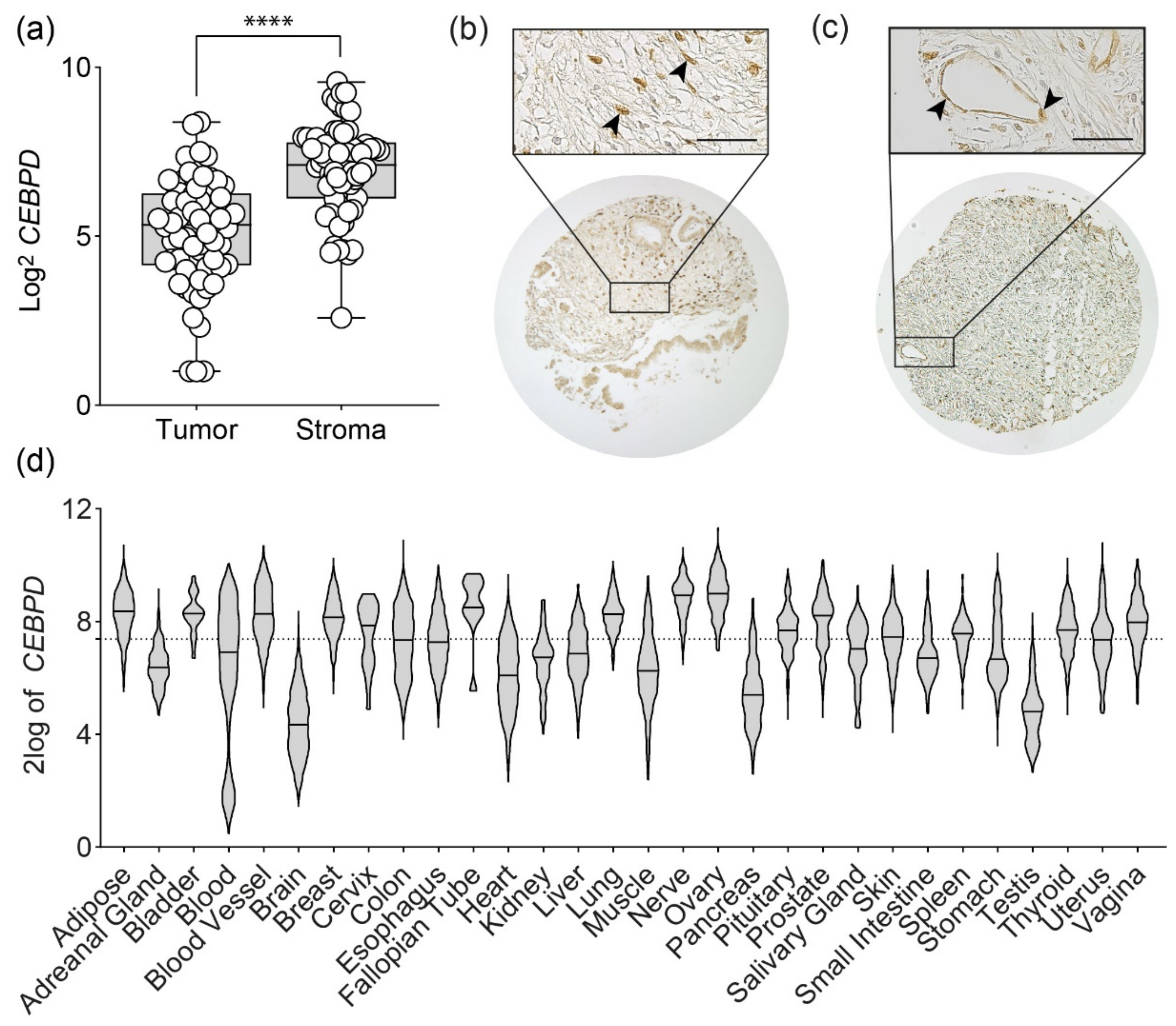{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Microarray (TMA)
2.2. Mining of Publicly Available RNA Microarray Datasets
2.3. Mice
2.4. Cells and Cell Culture
2.5. Orthotopic Pancreatic Cancer Model
2.6. Experimental Extravasation Model
2.7. Data Analysis
3. Results
3.1. C/EBPδ Is Not Tumor-Specific and Is Widely Expressed in Different Tissues
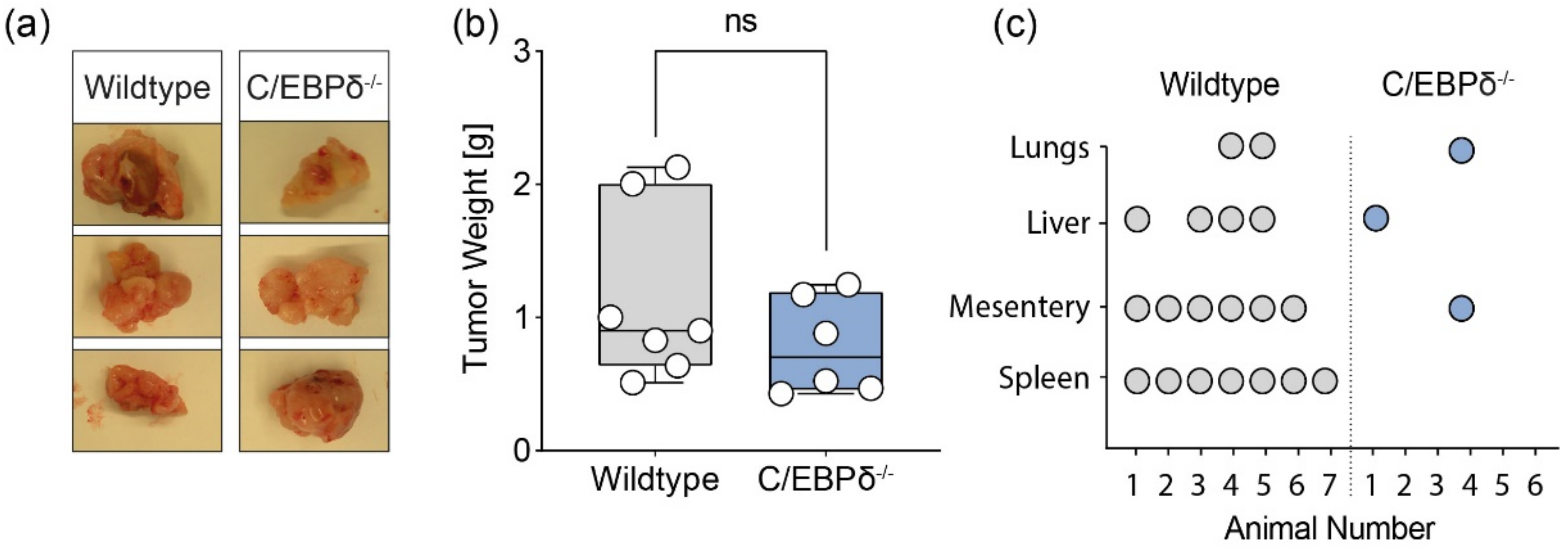
3.2. Non-Tumor C/EBPδ Potentiates Pancreatic Cancer Metastases
3.3. C/EBPδ Enhances Experimental Cancer Cell Extravasation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Rawla, P.; Sunkara, T.; Gaduputi, V. Epidemiology of Pancreatic Cancer: Global Trends, Etiology and Risk Factors. World J. Oncol. 2019, 10, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Orth, M.; Metzger, P.; Gerum, S.; Mayerle, J.; Schneider, G.; Belka, C.; Schnurr, M.; Lauber, K. Pancreatic ductal adenocarcinoma: Biological hallmarks, current status, and future perspectives of combined modality treatment approaches. Radiat. Oncol. 2019, 14, 1–20. [Google Scholar] [CrossRef]
- De Vita, F.; Ventriglia, J.; Febbraro, A.; Laterza, M.M.; Fabozzi, A.; Savastano, B.; Petrillo, A.; Diana, A.; Giordano, G.; Troiani, T.; et al. NAB-paclitaxel and gemcitabine in metastatic pancreatic ductal adenocarcinoma (PDAC): From clinical trials to clinical practice. BMC Cancer 2016, 16, 709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lekstrom-Himes, J.; Xanthopoulos, K.G. Biological role of the CCAAT/enhancer-binding protein family of transcription factors. J. Biol. Chem. 1998, 273, 28545–28548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gery, S.; Tanosaki, S.; Hofmann, W.K.; Koppel, A.; Koeffler, H.P. C/EBPδ expression in a BCR-ABL-positive cell line induces growth arrest and myeloid differentiation. Oncogene 2005, 24, 1589–1597. [Google Scholar] [CrossRef] [Green Version]
- Stein, T.; Salomonis, N.; Nuyten, D.S.A.; Vijver, M.J.; Gusterson, B.A. A mouse mammary gland involution mRNA signature identifies biological pathways potentially associated with breast cancer metastasis. J. Mammary Gland Biol. Neoplasia 2009, 14, 99–116. [Google Scholar] [CrossRef]
- Ikezoe, T.; Gery, S.; Yin, D.; Kelly, J.O.; Binderup, L.; Lemp, N.; Taguchi, H.; Koeffler, H.P.; O’Kelly, J.; Binderup, L.; et al. CCAAT/enhancer-binding protein delta: A molecular target of 1,25-dihydroxyvitamin D3 in androgen-responsive prostate cancer LNCaP cells. Cancer Res. 2005, 65, 4762–4768. [Google Scholar] [CrossRef] [Green Version]
- Sivko, G.S.; Sanford, D.C.; Dearth, L.D.; Tang, D.; DeWille, J.W. CCAAT/enhancer binding protein δ (C/EBPδ) regulation and expression in human mammary epithelial cells: II. Analysis of activating signal transduction pathways, transcriptional, post-transcriptional, and post-translational control. J. Cell. Biochem. 2004, 93, 844–856. [Google Scholar] [CrossRef]
- Tang, D.; Sivko, G.S.S.; De Wille, J.W. Promoter methylation reduces C/EBPδ (CEBPD) gene expression in the SUM-52PE human breast cancer cell line and in primary breast tumors. Breast Cancer Res. Treat. 2006, 95, 161–170. [Google Scholar] [CrossRef]
- Porter, D.; Weremowicz, S.; Chin, K.; Seth, P.; Keshaviah, A.; Lahti-Domenici, J.; Bae, Y.K.; Monitto, C.L.; Merlos-Suarez, A.; Chan, J.; et al. A neural survival factor is a candidate oncogene in breast cancer. Proc. Natl. Acad. Sci. USA 2003, 100, 10931–10936. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S.; Hofmann, W.-K.; Tidow, N.; Ehrich, M.; van den Boom, D.; Koschmieder, S.; Berdel, W.E.; Serve, H.; Müller-Tidow, C. The C/EBPδ tumor suppressor is silenced by hypermethylation in acute myeloid leukemia. Blood 2007, 109, 3895–3905. [Google Scholar] [CrossRef]
- Ko, C.Y.; Hsu, H.C.; Shen, M.R.; Chang, W.C.; Wang, J.M. Epigenetic silencing of CCAAT/enhancer-binding protein δ activity by YY1/polycomb group/DNA methyltransferase complex. J. Biol. Chem. 2008, 283, 30919–30932. [Google Scholar] [CrossRef] [Green Version]
- Barresi, V.; Vitarelli, E.; Cerasoli, S.; Barresi, G. The cell growth inhibitory transcription factor C/EBPd is expressed in human meningiomas in association with low histological grade and proliferation index. J. Neurooncol. 2010, 97, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Villanueva, D.; Balamurugan, K.; Ali, H.R.; Kim, S.-R.; Sharan, S.; Johnson, R.C.; Merchant, A.S.; Caldas, C.; Landberg, G.; Sterneck, E. The C/EBPδ protein is stabilized by estrogen receptor α activity, inhibits SNAI2 expression, and associates with good prognosis in breast cancer. Oncogene 2016, 35, 6166–6176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, C.; Monteverde, M.; Lattanzio, L.; Gojis, O.; Rudraraju, B.; Fortunato, M.; Syed, N.; Thompson, A.; Garrone, O.; Merlano, M.; et al. Site-specific CpG methylation in the CCAAT/enhancer binding protein delta (CEBPD) CpG island in breast cancer is associated with metastatic relapse. Br. J. Cancer 2012, 107, 732–738. [Google Scholar] [CrossRef] [Green Version]
- Hartl, L.; Duitman, J.; Aberson, H.L.; Chen, K.; Dijk, F.; Roelofs, J.J.T.H.; Dings, M.P.G.; Hooijer, G.K.J.; Hernanda, P.Y.; Pan, Q.; et al. CCAAT/Enhancer-Binding Protein Delta (C/EBPδ): A previously unrecognized tumor suppressor that limits the oncogenic potential of pancreatic ductal adenocarcinoma cells. Cancers 2020, 12, 2546. [Google Scholar] [CrossRef]
- Milne, G.R.; Palmer, T.M.; Yarwood, S.J. Novel control of cAMP-regulated transcription in vascular endothelial cells. Biochem. Soc. Trans. 2012, 40, 1–5. [Google Scholar] [CrossRef]
- Kolyada, A.Y.; Madias, N.E. Transcriptional regulation of the human iNOS gene by IL-1β in endothelial cells. Mol. Med. 2001, 7, 329–343. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.-C.; Kim, I.; Lye, E.; Shen, F.; Suzuki, N.; Suzuki, S.; Gerondakis, S.; Akira, S.; Gaffen, S.L.; Yeh, W.-C.; et al. Differential Role for c-Rel and C/EBPβ/δ in TLR-Mediated Induction of Proinflammatory Cytokines. J. Immunol. 2009, 182, 7212–7221. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.H.; Huang, H.S.; Wu, P.T.; Jou, I.M.; Pan, M.H.; Chang, W.C.; Wang, D.D.H.; Wang, J.M. Role of Macrophage CCAAT/Enhancer Binding Protein Delta in the Pathogenesis of Rheumatoid Arthritis in Collagen-Induced Arthritic Mice. PLoS ONE 2012, 7, e45378. [Google Scholar] [CrossRef]
- Sterneck, E.; Paylor, R.; Jackson-Lewis, V.; Libbey, M.; Przedborski, S.; Tessarollo, L.; Crawley, J.N.; Johnson, P.F. Selectively enhanced contextual fear conditioning in mice lacking the transcriptional regulator CCAAT/enhancer binding protein δ. Neurobiology 1998, 95, 10908–10913. Available online: www.pnas.org (accessed on 3 February 2021). [CrossRef] [PubMed]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Athar, A.; Füllgrabe, A.; George, N.; Iqbal, H.; Huerta, L.; Ali, A.; Snow, C.; Fonseca, N.A.; Petryszak, R.; Papatheodorou, I.; et al. ArrayExpress update—From bulk to single-cell expression data. Nucleic Acids Res. 2019, 47, D711–D715. [Google Scholar] [CrossRef] [PubMed]
- Koster, J. R2: Genomics Analysis and Visualization Platform. Available online: http://r2.amc.nl (accessed on 8 February 2021).
- Renz, B.W.; Takahashi, R.; Tanaka, T.; Macchini, M.; Hayakawa, Y.; Dantes, Z.; Maurer, H.C.; Chen, X.; Jiang, Z.; Ilmer, M.; et al. β2 Adrenergic-Neurotrophin Feedforward Loop Promotes Pancreatic Cancer. Cancer Cell 2018, 33, 75–90.e7. [Google Scholar] [CrossRef] [Green Version]
- Pilarsky, C.; Ammerpohl, O.; Sipos, B.; Dahl, E.; Hartmann, A.; Wellmann, A.; Braunschweig, T.; Jesnowski, R.; Friess, H.; Wente, M.N.; et al. Activation of Wnt signalling in stroma from pancreatic cancer identified by gene expression profiling. J. Cell. Mol. Med. 2008, 12, 2823–2835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, K.; Queiroz, K.C.S.; Roelofs, J.J.T.H.; van Noesel, C.J.M.; Richel, D.J.; Spek, C.A. Protease-activated receptor 2 suppresses lymphangiogenesis and subsequent lymph node metastasis in a murine pancreatic cancer model. J. Pathol. 2014, 234, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, K.C.S.; Shi, K.; Duitman, J.W.; Aberson, H.L.; Wilmink, J.W.; van Noesel, C.J.M.; Richel, D.J.; Spek, C.A. Protease-activated receptor-1 drives pancreatic cancer progression and chemoresistance. Int. J. Cancer. 2014, 135, 2294–2304. [Google Scholar] [CrossRef] [PubMed]
- Sowamber, R.; Chehade, R.; Bitar, M.; Dodds, L.V.; Milea, A.; Slomovitz, B.; Shaw, P.A.; George, S.H.L. CCAAT/enhancer binding protein delta (C/EBPδ) demonstrates a dichotomous role in tumor initiation and promotion of epithelial carcinoma. EBioMedicine 2019, 44, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Lili, L.N.; Matyunina, L.V.; Walker, L.D.; Benigno, B.B.; McDonald, J.F. Molecular profiling predicts the existence of two functionally distinct classes of ovarian cancer stroma. BioMed Res. Int. 2013, 2013, 846387. [Google Scholar] [CrossRef] [Green Version]
- Casey, T.; Bond, J.; Tighe, S.; Hunter, T.; Lintault, L.; Patel, O.; Eneman, J.; Crocker, A.; Wthite, J.; Tessitore, J.; et al. Molecular signatures suggest a major role for stromal cells in development of invasive breast cancer. Breast Cancer Res. Treat. 2009, 114, 47–62. [Google Scholar] [CrossRef]
- Horowitz, N.A.; Blevins, E.A.; Miller, W.M.; Perry, A.R.; Talmage, K.E.; Mullins, E.S.; Flick, M.J.; Queiroz, K.C.S.; Shi, K.; Spek, C.A.; et al. Thrombomodulin is a determinant of metastasis through a mechanism linked to the thrombin binding domain but not the lectin-like domain. Blood 2011, 118, 2889–2895. [Google Scholar] [CrossRef]
- Van Sluis, G.L.; Niers, T.M.H.; Esmon, C.T.; Tigchelaar, W.; Richel, D.J.; Buller, H.R.; van Noorden, C.J.F.; Spek, C.A. Endogenous activated protein C limits cancer cell extravasation through sphingosine-1-phosphate receptor 1-mediated vascular endothelial barrier enhancement. Blood 2009, 114, 1968–1973. [Google Scholar] [CrossRef] [Green Version]
- Im, S.-Y.; Ko, H.-M.; Ko, Y.-S.; Kim, J.-W.; Lee, H.-K.; Ha, T.-Y.; Lee, H.B.; Oh, S.-J.; Bai, S.; Chung, K.-C.; et al. Augmentation of Tumor Metastasis by Platelet-activating Factor. Cancer Res. 1996, 56, 2662–2665. [Google Scholar] [PubMed]
- Duitman, J.W.; Schouten, M.; Groot, A.P.; Daalhuisen, J.B.; Florquin, S.; van der Poll, T.; Spek, C.A. CCAAT/enhancer-binding protein δ facilitates bacterial dissemination during pneumococcal pneumonia in a platelet-activating factor receptor-dependent manner. Proc. Natl. Acad. Sci. USA 2012, 109, 9113–9118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, L.A.D.; Gutman, D.A.; Chisolm, C.; Appin, C.; Kong, J.; Rong, Y.; Kurc, T.; van Meir, E.G.; Saltz, J.H.; Moreno, C.S.; et al. The tumor microenvironment strongly impacts master transcriptional regulators and gene expression class of glioblastoma. Am. J. Pathol. 2012, 180, 2108–2119. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Wu, W.J.; Wang, W.J.; Huang, H.Y.; Li, W.M.; Yeh, B.W.; Wu, T.F.; Shiue, Y.L.; Sheu, J.J.C.; Wang, J.M.; et al. CEBPD amplification and overexpression in urothelial carcinoma: A driver of tumor metastasis indicating adverse prognosis. Oncotarget 2015, 6, 31069–31084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, C.Y.; Chang, W.C.; Wang, J.M. Biological roles of CCAAT/enhancer-binding protein delta during inflammation. J. Biomed. Sci. 2015, 22, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chi, J.-Y.; Hsiao, Y.-W.; Li, C.-F.; Lo, Y.-C.; Lin, Z.-Y.; Hong, J.-Y.; Liu, Y.-M.; Han, X.; Wang, S.-M.; Chen, B.-K.; et al. Targeting chemotherapy-induced PTX3 in tumor stroma to prevent the progression of drug-resistant cancers. Oncotarget 2015, 6, 23987–24001. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Zhang, H.H.; Singh, S.P.; Koo, L.; Kabat, J.; Tsang, H.; Singh, T.P.; Farber, J.M. C/EBPδ drives interactions between human MAIT cells and endothelial cells that are important for extravasation. Elife 2018, 7, e32532. [Google Scholar] [CrossRef]
- Russano, M.; Napolitano, A.; Ribelli, G.; Iuliai, M.; Simonetti, S.; Citarella, F.; Pantano, F.; Dell’Aquila, E.; Anesi, C.; Silvestris, N.; et al. Liquit biopsy and tumor heterogeneity in metastatic solid tumors: The potentiality of blood samples. J. Exp. Clin. Cancer Res. 2020, 39, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Su, G.H. Development of orthotopic pancreatic tumor mouse models. Methods Mol. Biol. 2013, 980, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partecke, L.I.; Sendler, M.; Kaeding, A.; Weiss, F.U.; Mayerle, J.; Dummer, A.; Nguyen, T.D.; Albers, N.; Speerforck, S.; Lerch, M.M.; et al. A syngeneic orthotopic murine model of pancreatic adenocarcinoma in the C57/BL6 mouse using the panc02 and 6606PDA cell lines. Eur. Surg. Res. 2011, 47, 98–107. [Google Scholar] [CrossRef]
- Butler, T.P.; Gullino, P.M. Quantitation of Cell Shedding into Efferent Blood of Mammary Adenocarcinoma. Cancer Res. 1975, 35, 512–516. [Google Scholar] [PubMed]
- Pierga, J.Y.; Bonneton, C.; Vincent-Salomon, A.; de Cremoux, P.; Nos, C.; Blin, N.; Pouillart, P.; Thiery, J.P.; Magdelénat, H. Clinical Significance of Immunocytochemical Detection of Tumor Cells Using Digital Microscopy in Peripheral Blood and Bone Marrow of Breast Cancer Patients. Clin. Cancer Res. 2004, 10, 1392–1400. [Google Scholar] [CrossRef] [Green Version]
- Chambers, A.F.; Groom, A.C.; MacDonald, I.C. Dissemination and growth of cancer cells in metastatic sites. Nat. Rev. Cancer 2002, 2, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Liu, L.; Zhang, H.; Liu, Z.L. Effect of Platelet-activating factor on barrier function of ARPE-19 cells. bioRxiv 2018, 398131. [Google Scholar] [CrossRef]
- Brailoiu, E.; Barlow, C.L.; Ramirez, S.H.; Abood, M.E.; Brailoiu, G.C. Effects of Platelet-Activating Factor on Brain Microvascular Endothelial Cells. Neuroscience 2018, 377, 105–113. [Google Scholar] [CrossRef]
- Hackler, P.C.; Reuss, S.; Konger, R.L.; Travers, J.B.; Sahu, R.P. Systemic Platelet-activating Factor Receptor Activation Augments Experimental Lung Tumor Growth and Metastasis. Cancer Growth Metastasis 2014, 7, 27–32. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duitman, J.; Hartl, L.; Roelofs, J.J.T.H.; Bijlsma, M.F.; Spek, C.A. Non-Tumor CCAAT/Enhancer-Binding Protein Delta Potentiates Tumor Cell Extravasation and Pancreatic Cancer Metastasis Formation. Biomolecules 2021, 11, 1079. https://doi.org/10.3390/biom11081079
Duitman J, Hartl L, Roelofs JJTH, Bijlsma MF, Spek CA. Non-Tumor CCAAT/Enhancer-Binding Protein Delta Potentiates Tumor Cell Extravasation and Pancreatic Cancer Metastasis Formation. Biomolecules. 2021; 11(8):1079. https://doi.org/10.3390/biom11081079
Chicago/Turabian StyleDuitman, JanWillem, Leonie Hartl, Joris J. T. H. Roelofs, Maarten F. Bijlsma, and C. Arnold Spek. 2021. "Non-Tumor CCAAT/Enhancer-Binding Protein Delta Potentiates Tumor Cell Extravasation and Pancreatic Cancer Metastasis Formation" Biomolecules 11, no. 8: 1079. https://doi.org/10.3390/biom11081079
APA StyleDuitman, J., Hartl, L., Roelofs, J. J. T. H., Bijlsma, M. F., & Spek, C. A. (2021). Non-Tumor CCAAT/Enhancer-Binding Protein Delta Potentiates Tumor Cell Extravasation and Pancreatic Cancer Metastasis Formation. Biomolecules, 11(8), 1079. https://doi.org/10.3390/biom11081079