
Pathophysiological Roles of Histamine Receptors in Cancer Progression: Implications and Perspectives as Potential Molecular Targets

Abstract
:1. Introduction
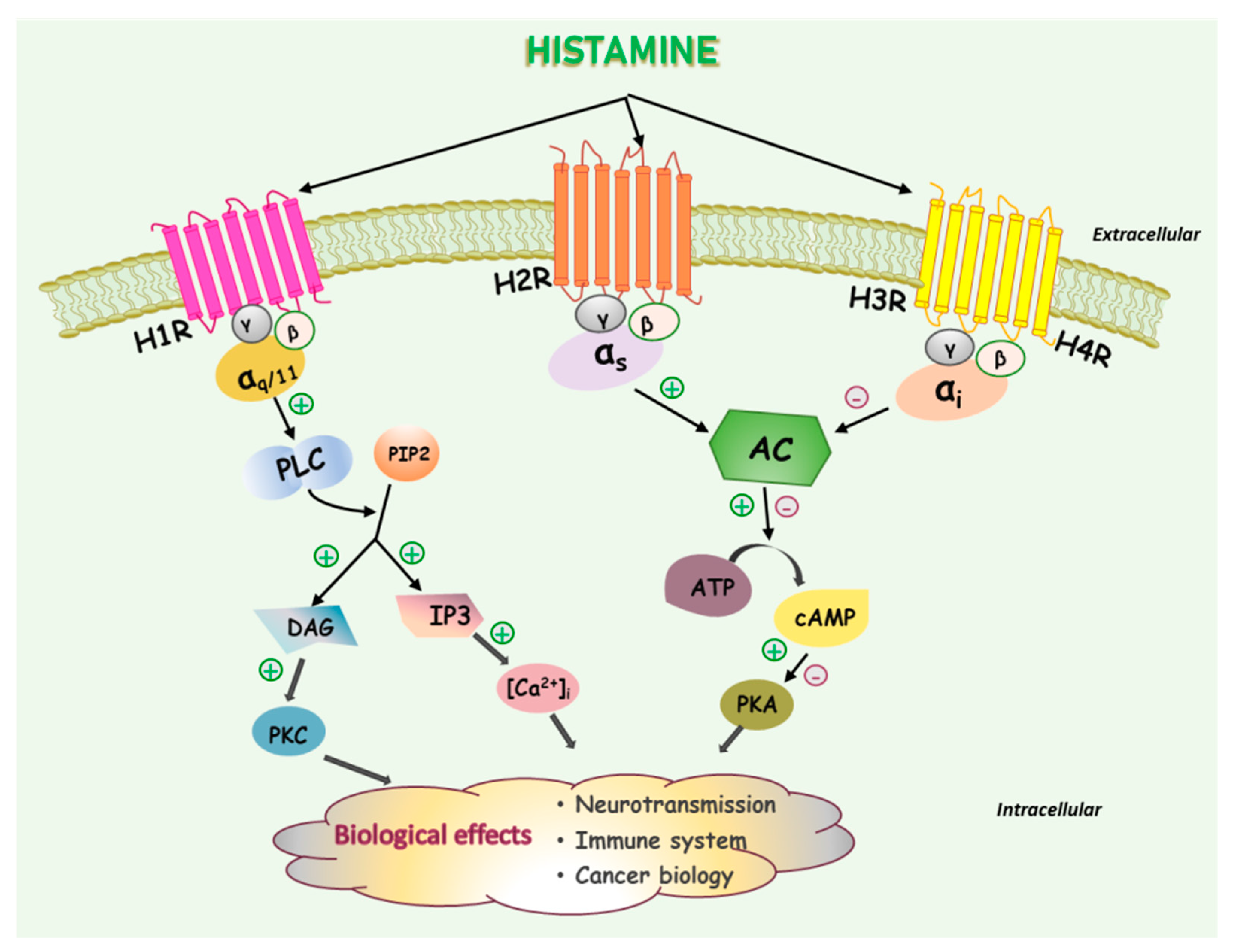
2. Histamine and Histamine Receptors
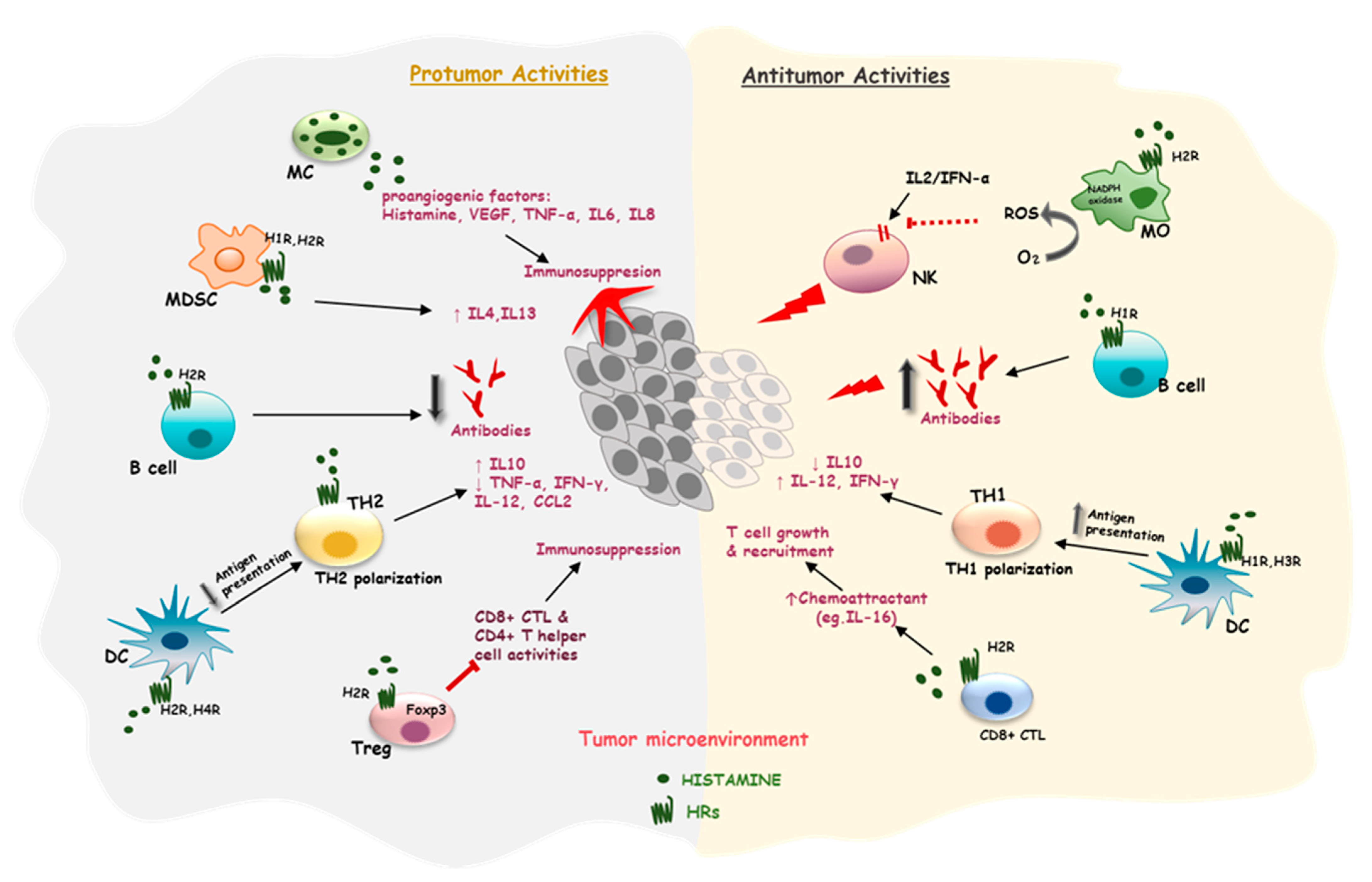
3. Effects of Histamine on the Immune Cells in a Tumor Microenvironment (TME)
3.1. Effects of Histamine on Mast Cells
3.2. Effects of Histamine on Antigen-Presenting Cells
3.3. Effects of Histamine on T Cells
3.4. Effects of Histamine on Myeloid-Derived Suppressor Cells
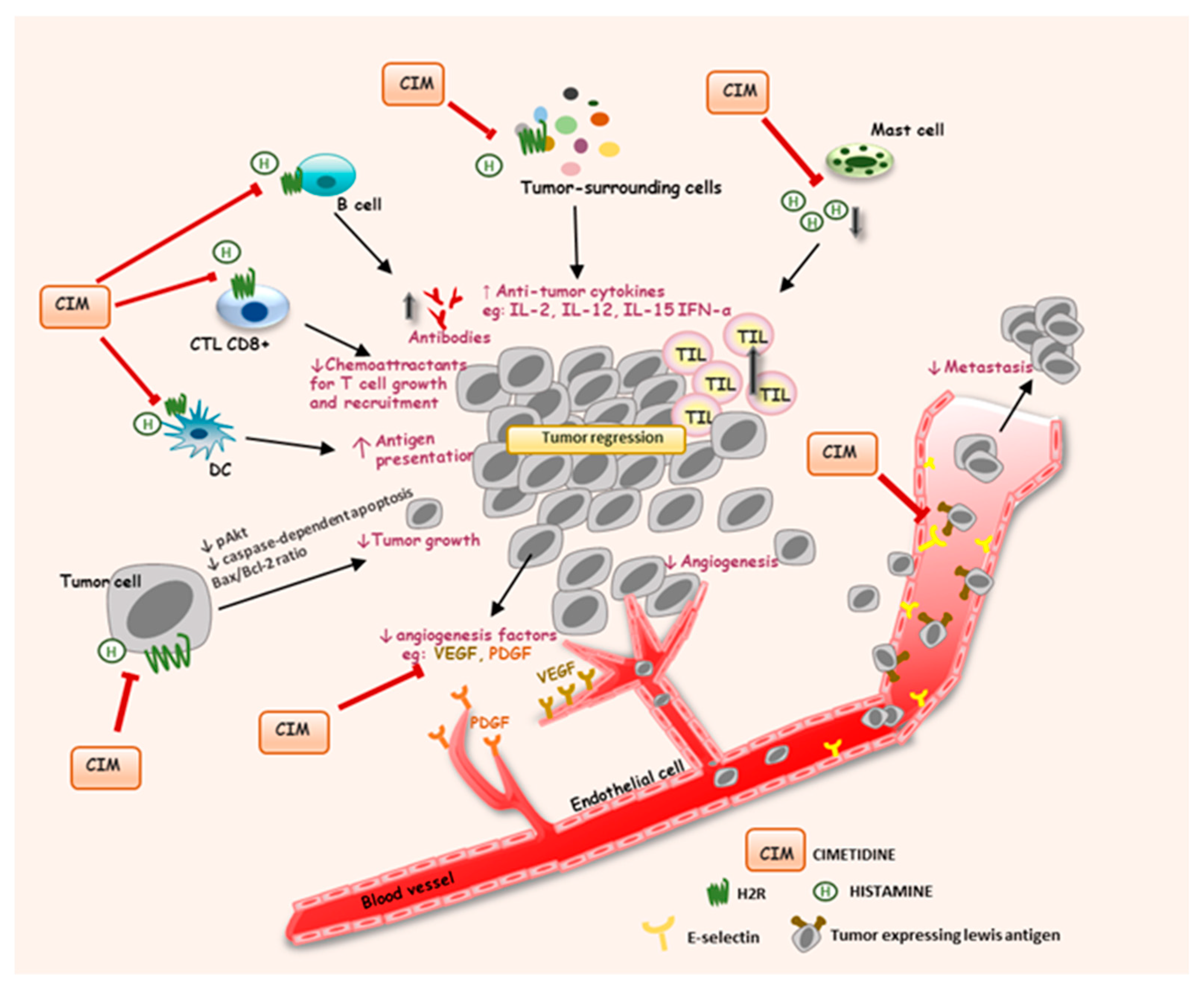
4. Pathophysiological Roles of Histamine and Histamine Receptors in Cancer and Their Implications as Potential Targets in Cancer Treatment
4.1. Histamine in Cancer Progression
4.2. Histamine Receptors in the Regulation of Cancer Progression and Their Implications as Potential Therapeutic Targets
4.2.1. Histamine H1 Receptor
4.2.2. Histamine H2 Receptor
4.2.3. Histamine H3 Receptor
4.2.4. Histamine H4 Receptor
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Kamińska, K.; Szczylik, C.; Bielecka, Z.F.; Bartnik, E.; Porta, C.; Lian, F.; Czarnecka, A.M. The role of the cell-cell interactions in cancer progression. J. Cell. Mol. Med. 2015, 19, 283–296. [Google Scholar] [CrossRef]
- Sousa, B.; Pereira, J.; Paredes, J. The Crosstalk Between Cell Adhesion and Cancer Metabolism. Int. J. Mol. Sci. 2019, 20, 1933. [Google Scholar] [CrossRef] [Green Version]
- Schirrmacher, V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment (Review). Int. J. Oncol. 2019, 54, 407–419. [Google Scholar] [CrossRef]
- Hirata, E.; Sahai, E. Tumor Microenvironment and Differential Responses to Therapy. Cold Spring Harb. Perspect. Med. 2017, 7, a026781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Cho, J.; Lee, K. Tumour Regression via Integrative Regulation of Neurological, Inflammatory, and Hypoxic Tumour Microenvironment. Biomol. Ther. 2020, 28, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branco, A.; Yoshikawa, F.S.Y.; Pietrobon, A.J.; Sato, M.N. Role of Histamine in Modulating the Immune Response and Inflammation. Mediat. Inflamm. 2018, 2018, 9524075. [Google Scholar] [CrossRef] [PubMed]
- Borriello, F.; Iannone, R.; Marone, G. Histamine Release from Mast Cells and Basophils. Handb. Exp. Pharmacol. 2017, 241, 121–139. [Google Scholar] [CrossRef]
- Massari, N.A.; Nicoud, M.B.; Medina, V.A. Histamine receptors and cancer pharmacology: An update. Br. J. Pharmacol. 2020, 177, 516–538. [Google Scholar] [CrossRef] [Green Version]
- Medina, V.A.; Rivera, E.S. Histamine receptors and cancer pharmacology. Br. J. Pharmacol. 2010, 161, 755–767. [Google Scholar] [CrossRef] [Green Version]
- Medina, V.; Croci, M.; Crescenti, E.; Mohamad, N.; Sanchez-Jiménez, F.; Massari, N.; Nuñez, M.; Cricco, G.; Martin, G.; Bergoc, R.; et al. The role of histamine in human mammary carcinogenesis: H3 and H4 receptors as potential therapeutic targets for breast cancer treatment. Cancer Biol. Ther. 2008, 7, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Davenas, E.; Rouleau, A.; Morisset, S.; Arrang, J.M. Autoregulation of McA-RH7777 hepatoma cell proliferation by histamine H3 receptors. J. Pharmacol. Exp. Ther. 2008, 326, 406–413. [Google Scholar] [CrossRef]
- Cricco, G.; Martin, G.; Labombarda, F.; Cocca, C.; Bergoc, R.; Rivera, E. Human pancreatic carcinoma cell line Panc-I and the role of histamine in growth regulation. Inflamm. Res. 2000, 49 (Suppl. S1), S68–S69. [Google Scholar] [CrossRef]
- Cricco, G.P.; Mohamad, N.A.; Sambuco, L.A.; Genre, F.; Croci, M.; Gutiérrez, A.S.; Medina, V.A.; Bergoc, R.M.; Rivera, E.S.; Martín, G.A. Histamine regulates pancreatic carcinoma cell growth through H3 and H4 receptors. Inflamm. Res. 2008, 57 (Suppl. S1), S23–S24. [Google Scholar] [CrossRef]
- Falus, A.; Hegyesi, H.; Lázár-Molnár, E.; Pós, Z.; László, V.; Darvas, Z. Paracrine and autocrine interactions in melanoma: Histamine is a relevant player in local regulation. Trends Immunol. 2001, 22, 648–652. [Google Scholar] [CrossRef]
- Rivera, E.S.; Cricco, G.P.; Engel, N.I.; Fitzsimons, C.P.; Martín, G.A.; Bergoc, R.M. Histamine as an autocrine growth factor: An unusual role for a widespread mediator. Semin. Cancer Biol. 2000, 10, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Bartholeyns, J.; Bouclier, M. Involvement of histamine in growth of mouse and rat tumors: Antitumoral properties of monofluoromethylhistidine, an enzyme-activated irreversible inhibitor of histidine decarboxylase. Cancer Res. 1984, 44, 639–645. [Google Scholar] [PubMed]
- Reynolds, J.L.; Akhter, J.; Morris, D.L. In vitro effect of histamine and histamine H1 and H2 receptor antagonists on cellular proliferation of human malignant melanoma cell lines. Melanoma Res. 1996, 6, 95–99. [Google Scholar] [CrossRef]
- Kubecova, M.; Kolostova, K.; Pinterova, D.; Kacprzak, G.; Bobek, V. Cimetidine: An anticancer drug? Eur. J. Pharm. Sci. 2011, 42, 439–444. [Google Scholar] [CrossRef]
- Parsons, M.E.; Ganellin, C.R. Histamine and its receptors. Br. J. Pharmacol. 2006, 147 (Suppl. S1), S127–S135. [Google Scholar] [CrossRef] [Green Version]
- Lieberman, P. The basics of histamine biology. Ann. Allergy Asthma Immunol. 2011, 106, S2–S5. [Google Scholar] [CrossRef] [PubMed]
- Panula, P.; Chazot, P.L.; Cowart, M.; Gutzmer, R.; Leurs, R.; Liu, W.L.; Stark, H.; Thurmond, R.L.; Haas, H.L. International Union of Basic and Clinical Pharmacology. XCVIII. Histamine Receptors. Pharmacol. Rev. 2015, 67, 601–655. [Google Scholar] [CrossRef] [Green Version]
- Seifert, R.; Strasser, A.; Schneider, E.H.; Neumann, D.; Dove, S.; Buschauer, A. Molecular and cellular analysis of human histamine receptor subtypes. Trends Pharmacol. Sci. 2013, 34, 33–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashina, K.; Tsubosaka, Y.; Nakamura, T.; Omori, K.; Kobayashi, K.; Hori, M.; Ozaki, H.; Murata, T. Histamine Induces Vascular Hyperpermeability by Increasing Blood Flow and Endothelial Barrier Disruption In Vivo. PLoS ONE 2015, 10, e0132367. [Google Scholar] [CrossRef]
- Nieto-Alamilla, G.; Márquez-Gómez, R.; García-Gálvez, A.M.; Morales-Figueroa, G.E.; Arias-Montaño, J.A. The Histamine H3 Receptor: Structure, Pharmacology, and Function. Mol. Pharmacol. 2016, 90, 649–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiligada, E.; Zampeli, E.; Sander, K.; Stark, H. Histamine H3 and H4 receptors as novel drug targets. Expert Opin. Investig. Drugs 2009, 18, 1519–1531. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.W.; Chen, Z. Role of histamine and its receptors in cerebral ischemia. ACS Chem. Neurosci. 2012, 3, 238–247. [Google Scholar] [CrossRef] [Green Version]
- Zampeli, E.; Tiligada, E. The role of histamine H4 receptor in immune and inflammatory disorders. Br. J. Pharmacol. 2009, 157, 24–33. [Google Scholar] [CrossRef]
- Schneider, E.H.; Seifert, R. The histamine H4-receptor and the central and peripheral nervous system: A critical analysis of the literature. Neuropharmacology 2016, 106, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Blaya, B.; Nicolau-Galmés, F.; Jangi, S.M.; Ortega-Martínez, I.; Alonso-Tejerina, E.; Burgos-Bretones, J.; Pérez-Yarza, G.; Asumendi, A.; Boyano, M.D. Histamine and histamine receptor antagonists in cancer biology. Inflamm. Allergy Drug Targets 2010, 9, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Akdis, C.A.; Blaser, K. Histamine in the immune regulation of allergic inflammation. J. Allergy Clin. Immunol. 2003, 112, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Jutel, M.; Akdis, M.; Akdis, C.A. Histamine, histamine receptors and their role in immune pathology. Clin. Exp. Allergy 2009, 39, 1786–1800. [Google Scholar] [CrossRef]
- Nicoud, M.B.; Formoso, K.; Medina, V.A. Pathophysiological Role of Histamine H4 Receptor in Cancer: Therapeutic Implications. Front. Pharmacol. 2019, 10, 556. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Sakaguchi, M.; Yoneyama, H.; Usami, Y.; Harusawa, S. Histamine H(3) receptor antagonist OUP-186 attenuates the proliferation of cultured human breast cancer cell lines. Biochem. Biophys. Res. Commun. 2016, 480, 479–485. [Google Scholar] [CrossRef]
- Martinel Lamas, D.J.; Croci, M.; Carabajal, E.; Crescenti, E.J.; Sambuco, L.; Massari, N.A.; Bergoc, R.M.; Rivera, E.S.; Medina, V.A. Therapeutic potential of histamine H4 receptor agonists in triple-negative human breast cancer experimental model. Br. J. Pharmacol. 2013, 170, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Gasser, S.; Lim, L.H.K.; Cheung, F.S.G. The role of the tumour microenvironment in immunotherapy. Endocr. Relat. Cancer 2017, 24, T283–T295. [Google Scholar] [CrossRef]
- O’Mahony, L.; Akdis, M.; Akdis, C.A. Regulation of the immune response and inflammation by histamine and histamine receptors. J. Allergy Clin. Immunol. 2011, 128, 1153–1162. [Google Scholar] [CrossRef]
- MacGlashan, D., Jr. Histamine: A mediator of inflammation. J. Allergy Clin. Immunol. 2003, 112, S53–S59. [Google Scholar] [CrossRef]
- Marichal, T.; Tsai, M.; Galli, S.J. Mast cells: Potential positive and negative roles in tumor biology. Cancer Immunol. Res. 2013, 1, 269–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurish, M.F.; Austen, K.F. The diverse roles of mast cells. J. Exp. Med. 2001, 194, F1–F5. [Google Scholar] [CrossRef] [PubMed]
- Derakhshani, A.; Vahidian, F.; Alihasanzadeh, M.; Mokhtarzadeh, A.; Lotfi Nezhad, P.; Baradaran, B. Mast cells: A double-edged sword in cancer. Immunol. Lett. 2019, 209, 28–35. [Google Scholar] [CrossRef]
- Galdiero, M.R.; Varricchi, G.; Seaf, M.; Marone, G.; Levi-Schaffer, F.; Marone, G. Bidirectional Mast Cell-Eosinophil Interactions in Inflammatory Disorders and Cancer. Front. Med. 2017, 4, 103. [Google Scholar] [CrossRef] [Green Version]
- Aller, M.A.; Arias, A.; Arias, J.I.; Arias, J. Carcinogenesis: The cancer cell-mast cell connection. Inflamm. Res. 2019, 68, 103–116. [Google Scholar] [CrossRef]
- Coussens, L.M.; Raymond, W.W.; Bergers, G.; Laig-Webster, M.; Behrendtsen, O.; Werb, Z.; Caughey, G.H.; Hanahan, D. Inflammatory mast cells up-regulate angiogenesis during squamous epithelial carcinogenesis. Genes Dev. 1999, 13, 1382–1397. [Google Scholar] [CrossRef]
- Conti, P.; Castellani, M.L.; Kempuraj, D.; Salini, V.; Vecchiet, J.; Tetè, S.; Mastrangelo, F.; Perrella, A.; De Lutiis, M.A.; Tagen, M.; et al. Role of mast cells in tumor growth. Ann. Clin. Lab. Sci. 2007, 37, 315–322. [Google Scholar] [PubMed]
- Galli, S.J.; Tsai, M. Mast cells: Versatile regulators of inflammation, tissue remodeling, host defense and homeostasis. J. Dermatol. Sci. 2008, 49, 7–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komi, D.E.A.; Khomtchouk, K.; Santa Maria, P.L. A Review of the Contribution of Mast Cells in Wound Healing: Involved Molecular and Cellular Mechanisms. Clin. Rev. Allergy Immunol. 2020, 58, 298–312. [Google Scholar] [CrossRef]
- Norrby, K. Mast cells and angiogenesis. APMIS 2002, 110, 355–371. [Google Scholar] [CrossRef]
- Saxena, S.; Singh, A.; Singh, P. Tumor associated mast cells: Biological roles and therapeutic applications. Anat. Cell Biol. 2020, 53, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Enamorado, M.; Iborra, S.; Priego, E.; Cueto, F.J.; Quintana, J.A.; Martínez-Cano, S.; Mejías-Pérez, E.; Esteban, M.; Melero, I.; Hidalgo, A.; et al. Enhanced anti-tumour immunity requires the interplay between resident and circulating memory CD8(+) T cells. Nat. Commun. 2017, 8, 16073. [Google Scholar] [CrossRef]
- Varricchi, G.; Galdiero, M.R.; Loffredo, S.; Marone, G.; Iannone, R.; Marone, G.; Granata, F. Are Mast Cells MASTers in Cancer? Front. Immunol. 2017, 8, 424. [Google Scholar] [CrossRef] [Green Version]
- Hofstra, C.L.; Desai, P.J.; Thurmond, R.L.; Fung-Leung, W.P. Histamine H4 receptor mediates chemotaxis and calcium mobilization of mast cells. J. Pharmacol. Exp. Ther. 2003, 305, 1212–1221. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.K.; Hirasawa, N.; Ohtsu, H.; Watanabe, T.; Ohuchi, K. Defective angiogenesis in the inflammatory granulation tissue in histidine decarboxylase-deficient mice but not in mast cell-deficient mice. J. Exp. Med. 2002, 195, 973–982. [Google Scholar] [CrossRef] [Green Version]
- Gaudino, S.J.; Kumar, P. Cross-Talk Between Antigen Presenting Cells and T Cells Impacts Intestinal Homeostasis, Bacterial Infections, and Tumorigenesis. Front. Immunol. 2019, 10, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellstrand, K.; Kylefjord, H.; Asea, A.; Hermodsson, S. Regulation of the natural killer cell response to interferon-alpha by biogenic amines. J. Interferon Res. 1992, 12, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Hellstrand, K.; Hermodsson, S. Synergistic activation of human natural killer cell cytotoxicity by histamine and interleukin-2. Int. Arch. Allergy Appl. Immunol. 1990, 92, 379–389. [Google Scholar] [CrossRef]
- Hellstrand, K.; Asea, A.; Dahlgren, C.; Hermodsson, S. Histaminergic regulation of NK cells. Role of monocyte-derived reactive oxygen metabolites. J. Immunol. 1994, 153, 4940–4947. [Google Scholar] [PubMed]
- Hellstrand, K.; Hermodsson, S. Histamine H2-receptor-mediated regulation of human natural killer cell activity. J. Immunol. 1986, 137, 656–660. [Google Scholar] [PubMed]
- Hansson, M.; Asea, A.; Ersson, U.; Hermodsson, S.; Hellstrand, K. Induction of apoptosis in NK cells by monocyte-derived reactive oxygen metabolites. J. Immunol. 1996, 156, 42–47. [Google Scholar] [PubMed]
- Medina, V.A.; Massari, N.A.; Cricco, G.P.; Martín, G.A.; Bergoc, R.M.; Rivera, E.S. Involvement of hydrogen peroxide in histamine-induced modulation of WM35 human malignant melanoma cell proliferation. Free. Radic. Biol. Med. 2009, 46, 1510–1515. [Google Scholar] [CrossRef] [PubMed]
- Perz, J.B.; Ho, A.D. Histamine dihydrochloride for the treatment of acute myeloid leukemia, malignant melanoma and renal cell carcinoma. Future Oncol. 2008, 4, 169–177. [Google Scholar] [CrossRef]
- Johansson, S.; Landström, M.; Hellstrand, K.; Henriksson, R. The response of Dunning R3327 prostatic adenocarcinoma to IL-2, histamine and radiation. Br. J. Cancer 1998, 77, 1213–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellstrand, K.; Hermodsson, S.; Naredi, P.; Mellqvist, U.H.; Brune, M. Histamine and cytokine therapy. Acta Oncol. 1998, 37, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Rydström, A.; Hallner, A.; Aurelius, J.; Sander, F.E.; Bernson, E.; Kiffin, R.; Thoren, F.B.; Hellstrand, K.; Martner, A. Dynamics of myeloid cell populations during relapse-preventive immunotherapy in acute myeloid leukemia. J. Leukoc. Biol. 2017, 102, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Palucka, K.; Banchereau, J. Cancer immunotherapy via dendritic cells. Nat. Rev. Cancer 2012, 12, 265–277. [Google Scholar] [CrossRef]
- Reid, C.D. The biology and clinical applications of dendritic cells. Transfus. Med. 1998, 8, 77–86. [Google Scholar] [CrossRef]
- Jutel, M.; Akdis, C. T-cell regulatory mechanisms in specific immunotherapy. Chem. Immunol. Allergy 2008, 94, 158–177. [Google Scholar] [CrossRef] [Green Version]
- Szeberényi, J.B.; Pállinger, E.; Zsinkó, M.; Pós, Z.; Rothe, G.; Orsó, E.; Szeberényi, S.; Schmitz, G.; Falus, A.; László, V. Inhibition of effects of endogenously synthesized histamine disturbs in vitro human dendritic cell differentiation. Immunol. Lett. 2001, 76, 175–182. [Google Scholar] [CrossRef]
- Mazzoni, A.; Young, H.A.; Spitzer, J.H.; Visintin, A.; Segal, D.M. Histamine regulates cytokine production in maturing dendritic cells, resulting in altered T cell polarization. J. Clin. Investig. 2001, 108, 1865–1873. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, A.; Leifer, C.A.; Mullen, G.E.; Kennedy, M.N.; Klinman, D.M.; Segal, D.M. Cutting edge: Histamine inhibits IFN-alpha release from plasmacytoid dendritic cells. J. Immunol. 2003, 170, 2269–2273. [Google Scholar] [CrossRef] [Green Version]
- van der Pouw Kraan, T.C.; Snijders, A.; Boeije, L.C.; de Groot, E.R.; Alewijnse, A.E.; Leurs, R.; Aarden, L.A. Histamine inhibits the production of interleukin-12 through interaction with H2 receptors. J. Clin. Investig. 1998, 102, 1866–1873. [Google Scholar] [CrossRef] [Green Version]
- Dijkstra, D.; Stark, H.; Chazot, P.L.; Shenton, F.C.; Leurs, R.; Werfel, T.; Gutzmer, R. Human inflammatory dendritic epidermal cells express a functional histamine H4 receptor. J. Investig. Dermatol. 2008, 128, 1696–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutzmer, R.; Diestel, C.; Mommert, S.; Köther, B.; Stark, H.; Wittmann, M.; Werfel, T. Histamine H4 receptor stimulation suppresses IL-12p70 production and mediates chemotaxis in human monocyte-derived dendritic cells. J. Immunol. 2005, 174, 5224–5232. [Google Scholar] [CrossRef] [PubMed]
- Colluru, V.T.; McNeel, D.G. B lymphocytes as direct antigen-presenting cells for anti-tumor DNA vaccines. Oncotarget 2016, 7, 67901–67918. [Google Scholar] [CrossRef] [Green Version]
- Rossetti, R.A.M.; Lorenzi, N.P.C.; Yokochi, K.; Rosa, M.; Benevides, L.; Margarido, P.F.R.; Baracat, E.C.; Carvalho, J.P.; Villa, L.L.; Lepique, A.P. B lymphocytes can be activated to act as antigen presenting cells to promote anti-tumor responses. PLoS ONE 2018, 13, e0199034. [Google Scholar] [CrossRef]
- Sarvaria, A.; Madrigal, J.A.; Saudemont, A. B cell regulation in cancer and anti-tumor immunity. Cell. Mol. Immunol. 2017, 14, 662–674. [Google Scholar] [CrossRef] [Green Version]
- Banu, Y.; Watanabe, T. Augmentation of antigen receptor-mediated responses by histamine H1 receptor signaling. J. Exp. Med. 1999, 189, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, M.; Kimata, H. Histamine inhibits immunoglobulin production via histamine H2 receptors without affecting cell growth in human B cells. Clin. Immunol. Immunopathol. 1994, 73, 96–102. [Google Scholar] [CrossRef]
- Okitsu-Negishi, S.; Furusawa, S.; Yoshino, K.; Abe, T. The suppressive effect of histamine on anti-DNP antibody production in mice. Int. Arch. Allergy Appl. Immunol. 1988, 86, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Rogers, D.; Vila-Leahey, A.; Pessôa, A.C.; Oldford, S.; Marignani, P.A.; Marshall, J.S. Ranitidine Inhibition of Breast Tumor Growth Is B Cell Dependent and Associated With an Enhanced Antitumor Antibody Response. Front. Immunol. 2018, 9, 1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okeke, E.B.; Uzonna, J.E. The Pivotal Role of Regulatory T Cells in the Regulation of Innate Immune Cells. Front. Immunol. 2019, 10, 680. [Google Scholar] [CrossRef] [Green Version]
- Chraa, D.; Naim, A.; Olive, D.; Badou, A. T lymphocyte subsets in cancer immunity: Friends or foes. J. Leukoc. Biol. 2019, 105, 243–255. [Google Scholar] [CrossRef]
- Sheikhi, A.; Jafarzadeh, A.; Kokhaei, P.; Hojjat-Farsangi, M. Whole Tumor Cell Vaccine Adjuvants: Comparing IL-12 to IL-2 and IL-15. Iran. J. Immunol. 2016, 13, 148–166. [Google Scholar]
- Jafarzadeh, A.; Minaee, K.; Farsinejad, A.R.; Nemati, M.; Khosravimashizi, A.; Daneshvar, H.; Mohammadi, M.M.; Sheikhi, A.; Ghaderi, A. Evaluation of the circulating levels of IL-12 and IL-33 in patients with breast cancer: Influences of the tumor stages and cytokine gene polymorphisms. Iran. J. Basic Med. Sci. 2015, 18, 1189–1198. [Google Scholar]
- Hansson, M.; Hermodsson, S.; Brune, M.; Mellqvist, U.H.; Naredi, P.; Betten, A.; Gehlsen, K.R.; Hellstrand, K. Histamine protects T cells and natural killer cells against oxidative stress. J. Interferon Cytokine Res. 1999, 19, 1135–1144. [Google Scholar] [CrossRef] [PubMed]
- Trinchieri, G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat. Rev. Immunol. 2003, 3, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-gamma: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef]
- Mocellin, S.; Panelli, M.C.; Wang, E.; Nagorsen, D.; Marincola, F.M. The dual role of IL-10. Trends Immunol. 2003, 24, 36–43. [Google Scholar] [CrossRef]
- Tomita, K.; Okabe, S. Exogenous histamine stimulates colorectal cancer implant growth via immunosuppression in mice. J. Pharmacol. Sci. 2005, 97, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Kmiecik, T.; Otocka-Kmiecik, A.; Górska-Ciebiada, M.; Ciebiada, M. T lymphocytes as a target of histamine action. Arch. Med. Sci. 2012, 8, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Khalife, E.; Khodadadi, A.; Talaeizadeh, A.; Rahimian, L.; Nemati, M.; Jafarzadeh, A. Overexpression of Regulatory T Cell-Related Markers (FOXP3, CTLA-4 and GITR) by Peripheral Blood Mononuclear Cells from Patients with Breast Cancer. Asian Pac. J. Cancer Prev. 2018, 19, 3019–3025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jutel, M.; Blaser, K.; Akdis, C.A. Histamine in allergic inflammation and immune modulation. Int. Arch. Allergy Immunol. 2005, 137, 82–92. [Google Scholar] [CrossRef]
- Kumar, A.; Cleveland, R.P. “Immunoregulatory effects of cimetidine: Inhibition of suppressor cell effector function in vivo”. Immunopharmacol. Immunotoxicol. 1988, 10, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Griswold, D.E.; Alessi, S.; Badger, A.M.; Poste, G.; Hanna, N. Inhibition of T suppressor cell expression by histamine type 2 (H2) receptor antagonists. J. Immunol. 1984, 132, 3054–3057. [Google Scholar]
- Zhang, Y.; Chen, Z.; Luo, X.; Wu, B.; Li, B.; Wang, B. Cimetidine down-regulates stability of Foxp3 protein via Stub1 in Treg cells. Hum. Vaccines Immunother. 2016, 12, 2512–2518. [Google Scholar] [CrossRef] [Green Version]
- Stagg, J.; Allard, B. Immunotherapeutic approaches in triple-negative breast cancer: Latest research and clinical prospects. Ther. Adv. Med. Oncol. 2013, 5, 169–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gantner, F.; Sakai, K.; Tusche, M.W.; Cruikshank, W.W.; Center, D.M.; Bacon, K.B. Histamine h(4) and h(2) receptors control histamine-induced interleukin-16 release from human CD8(+) T cells. J. Pharmacol. Exp. Ther. 2002, 303, 300–307. [Google Scholar] [CrossRef]
- Compérat, E.; Rouprêt, M.; Drouin, S.J.; Camparo, P.; Bitker, M.O.; Houlgatte, A.; Cancel-Tassin, G.; Cussenot, O. Tissue expression of IL16 in prostate cancer and its association with recurrence after radical prostatectomy. Prostate 2010, 70, 1622–1627. [Google Scholar] [CrossRef]
- Poschke, I.; Mao, Y.; Adamson, L.; Salazar-Onfray, F.; Masucci, G.; Kiessling, R. Myeloid-derived suppressor cells impair the quality of dendritic cell vaccines. Cancer Immunol. Immunother. 2012, 61, 827–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoechst, B.; Voigtlaender, T.; Ormandy, L.; Gamrekelashvili, J.; Zhao, F.; Wedemeyer, H.; Lehner, F.; Manns, M.P.; Greten, T.F.; Korangy, F. Myeloid derived suppressor cells inhibit natural killer cells in patients with hepatocellular carcinoma via the NKp30 receptor. Hepatology 2009, 50, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Han, Y.; Guo, Q.; Zhang, M.; Cao, X. Cancer-expanded myeloid-derived suppressor cells induce anergy of NK cells through membrane-bound TGF-beta 1. J. Immunol. 2009, 182, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Sinha, P.; Clements, V.K.; Ostrand-Rosenberg, S. Reduction of myeloid-derived suppressor cells and induction of M1 macrophages facilitate the rejection of established metastatic disease. J. Immunol. 2005, 174, 636–645. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Pan, P.Y.; Li, Q.; Sato, A.I.; Levy, D.E.; Bromberg, J.; Divino, C.M.; Chen, S.H. Gr-1+CD115+ immature myeloid suppressor cells mediate the development of tumor-induced T regulatory cells and T-cell anergy in tumor-bearing host. Cancer Res. 2006, 66, 1123–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009, 9, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.K.; Saleem, S.J.; Folgosa, L.; Zellner, H.B.; Damle, S.R.; Nguyen, G.K.; Ryan, J.J.; Bear, H.D.; Irani, A.M.; Conrad, D.H. Mast cell histamine promotes the immunoregulatory activity of myeloid-derived suppressor cells. J. Leukoc. Biol. 2014, 96, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.D.; Ai, W.; Asfaha, S.; Bhagat, G.; Friedman, R.A.; Jin, G.; Park, H.; Shykind, B.; Diacovo, T.G.; Falus, A.; et al. Histamine deficiency promotes inflammation-associated carcinogenesis through reduced myeloid maturation and accumulation of CD11b+Ly6G+ immature myeloid cells. Nat. Med. 2011, 17, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Xu, M.; Li, X.; Jia, J.; Fan, K.; Lai, G. Cimetidine suppresses lung tumor growth in mice through proapoptosis of myeloid-derived suppressor cells. Mol. Immunol. 2013, 54, 74–83. [Google Scholar] [CrossRef]
- Vila-Leahey, A.; Oldford, S.A.; Marignani, P.A.; Wang, J.; Haidl, I.D.; Marshall, J.S. Ranitidine modifies myeloid cell populations and inhibits breast tumor development and spread in mice. Oncoimmunology 2016, 5, e1151591. [Google Scholar] [CrossRef]
- Reynolds, J.L.; Akhter, J.; Adams, W.J.; Morris, D.L. Histamine content in colorectal cancer. Are there sufficient levels of histamine to affect lymphocyte function? Eur. J. Surg. Oncol. 1997, 23, 224–227. [Google Scholar] [CrossRef]
- von Mach-Szczypiński, J.; Stanosz, S.; Sieja, K.; Stanosz, M. Metabolism of histamine in tissues of primary ductal breast cancer. Metabolism 2009, 58, 867–870. [Google Scholar] [CrossRef] [PubMed]
- Hegyesi, H.; Somlai, B.; Varga, V.L.; Toth, G.; Kovacs, P.; Molnar, E.L.; Laszlo, V.; Karpati, S.; Rivera, E.; Falus, A.; et al. Suppression of melanoma cell proliferation by histidine decarboxylase specific antisense oligonucleotides. J. Investig. Dermatol. 2001, 117, 151–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graff, L.; Frungieri, M.; Zanner, R.; Pohlinger, A.; Prinz, C.; Gratzl, M. Expression of histidine decarboxylase and synthesis of histamine by human small cell lung carcinoma. Am. J. Pathol. 2002, 160, 1561–1565. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Caballero, M.; Neugebauer, E.; Campos, R.; Nunez de Castro, I.; Vara-Thorbeck, C. Increased histidine decarboxylase (HDC) activity in human colorectal cancer: Results of a study on ten patients. Agents Actions 1988, 23, 357–360. [Google Scholar] [CrossRef]
- Moriarty, C.M.; Stucky, J.L.; Hamburger, K.W.; Patil, K.D.; Foley, J.F.; Koefoot, R.R. Blood histamine and solid malignant tumors. J. Cancer Res. Clin. Oncol. 1988, 114, 588–592. [Google Scholar] [CrossRef]
- Cui, J.; Xu, G.; Liu, J.; Pang, Z.; Florholmen, J.; Cui, G. The expression of non-mast histamine in tumor associated microvessels in human colorectal cancers. Pathol. Oncol. Res. 2013, 19, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Pós, Z.; Sáfrány, G.; Müller, K.; Tóth, S.; Falus, A.; Hegyesi, H. Phenotypic profiling of engineered mouse melanomas with manipulated histamine production identifies histamine H2 receptor and rho-C as histamine-regulated melanoma progression markers. Cancer Res. 2005, 65, 4458–4466. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Ganesh, B.P.; Shi, Z.; Shah, R.R.; Fultz, R.; Major, A.; Venable, S.; Lugo, M.; Hoch, K.; Chen, X.; et al. Gut Microbe-Mediated Suppression of Inflammation-Associated Colon Carcinogenesis by Luminal Histamine Production. Am. J. Pathol. 2017, 187, 2323–2336. [Google Scholar] [CrossRef] [Green Version]
- Parihar, A.; Dube, A.; Gupta, P.K. Photodynamic treatment of oral squamous cell carcinoma in hamster cheek pouch model using chlorin p6-histamine conjugate. Photodiagnosis Photodyn. Ther. 2013, 10, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Monti Hughes, A.; Pozzi, E.; Thorp, S.I.; Curotto, P.; Medina, V.A.; Martinel Lamas, D.J.; Rivera, E.S.; Garabalino, M.A.; Farías, R.O.; Gonzalez, S.J.; et al. Histamine reduces boron neutron capture therapy-induced mucositis in an oral precancer model. Oral Dis. 2015, 21, 770–777. [Google Scholar] [CrossRef]
- Faustino-Rocha, A.I.; Ferreira, R.; Gama, A.; Oliveira, P.A.; Ginja, M. Antihistamines as promising drugs in cancer therapy. Life Sci. 2017, 172, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, H.; Kitada, K.; Nakai, K.; Sarai, A. PrognoScan: A new database for meta-analysis of the prognostic value of genes. BMC Med. Genom. 2009, 2, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Wei, X.; Shi, L.; Chen, B.; Zhao, G.; Yang, H. Integrative genomic analyses of the histamine H1 receptor and its role in cancer prediction. Int. J. Mol. Med. 2014, 33, 1019–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Hou, Y.; Yin, C.; Hu, J.; Gao, T.; Huang, X.; Zhang, X.; Xing, J.; An, J.; Wan, S.; et al. Upregulation of histamine receptor H1 promotes tumor progression and contributes to poor prognosis in hepatocellular carcinoma. Oncogene 2020, 39, 1724–1738. [Google Scholar] [CrossRef]
- Grimm, M.; Krimmel, M.; Alexander, D.; Munz, A.; Kluba, S.; Keutel, C.; Hoffmann, J.; Polligkeit, J.; Reinert, S.; Hoefert, S. Prognostic value of histamine H1 receptor expression in oral squamous cell carcinoma. Clin. Oral Investig. 2013, 17, 949–955. [Google Scholar] [CrossRef]
- Fernández-Nogueira, P.; Bragado, P.; Almendro, V.; Ametller, E.; Rios, J.; Choudhury, S.; Mancino, M.; Gascón, P. Differential expression of neurogenes among breast cancer subtypes identifies high risk patients. Oncotarget 2016, 7, 5313–5326. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Nogueira, P.; Noguera-Castells, A.; Fuster, G.; Recalde-Percaz, L.; Moragas, N.; López-Plana, A.; Enreig, E.; Jauregui, P.; Carbó, N.; Almendro, V.; et al. Histamine receptor 1 inhibition enhances antitumor therapeutic responses through extracellular signal-regulated kinase (ERK) activation in breast cancer. Cancer Lett. 2018, 424, 70–83. [Google Scholar] [CrossRef]
- Gómez-Fabre, P.M.; de Pedro, E.; Medina, M.A.; Núñez de Castro, I.; Márquez, J. Polyamine contents of human breast cancer cells treated with the cytotoxic agents chlorpheniramine and dehydrodidemnin B. Cancer Lett. 1997, 113, 141–144. [Google Scholar] [CrossRef]
- Jangi, S.M.; Díaz-Pérez, J.L.; Ochoa-Lizarralde, B.; Martín-Ruiz, I.; Asumendi, A.; Pérez-Yarza, G.; Gardeazabal, J.; Díaz-Ramón, J.L.; Boyano, M.D. H1 histamine receptor antagonists induce genotoxic and caspase-2-dependent apoptosis in human melanoma cells. Carcinogenesis 2006, 27, 1787–1796. [Google Scholar] [CrossRef]
- Jangi, S.M.; Ruiz-Larrea, M.B.; Nicolau-Galmés, F.; Andollo, N.; Arroyo-Berdugo, Y.; Ortega-Martínez, I.; Díaz-Pérez, J.L.; Boyano, M.D. Terfenadine-induced apoptosis in human melanoma cells is mediated through Ca2+ homeostasis modulation and tyrosine kinase activity, independently of H1 histamine receptors. Carcinogenesis 2008, 29, 500–509. [Google Scholar] [CrossRef] [Green Version]
- Or, C.R.; Su, H.L.; Lee, W.C.; Yang, S.Y.; Ho, C.; Chang, C.C. Diphenhydramine induces melanoma cell apoptosis by suppressing STAT3/MCL-1 survival signaling and retards B16-F10 melanoma growth in vivo. Oncol. Rep. 2016, 36, 3465–3471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jangi, S.M.; Asumendi, A.; Arlucea, J.; Nieto, N.; Perez-Yarza, G.; Morales, M.C.; de la Fuente-Pinedo, M.; Boyano, M.D. Apoptosis of human T-cell acute lymphoblastic leukemia cells by diphenhydramine, an H1 histamine receptor antagonist. Oncol. Res. 2004, 14, 363–372. [Google Scholar] [CrossRef]
- Verdoodt, F.; Dehlendorff, C.; Jäättelä, M.; Strauss, R.; Pottegård, A.; Hallas, J.; Friis, S.; Kjaer, S.K. Antihistamines and Ovarian Cancer Survival: Nationwide Cohort Study and in Vitro Cell Viability Assay. J. Natl. Cancer Inst. 2020, 112, 964–967. [Google Scholar] [CrossRef]
- Wang, W.T.; Chen, Y.H.; Hsu, J.L.; Leu, W.J.; Yu, C.C.; Chan, S.H.; Ho, Y.F.; Hsu, L.C.; Guh, J.H. Terfenadine induces anti-proliferative and apoptotic activities in human hormone-refractory prostate cancer through histamine receptor-independent Mcl-1 cleavage and Bak up-regulation. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2014, 387, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug resistance in cancer: An overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef] [Green Version]
- An, L.; Li, D.D.; Chu, H.X.; Zhang, Q.; Wang, C.L.; Fan, Y.H.; Song, Q.; Ma, H.D.; Feng, F.; Zhao, Q.C. Terfenadine combined with epirubicin impedes the chemo-resistant human non-small cell lung cancer both in vitro and in vivo through EMT and Notch reversal. Pharmacol. Res. 2017, 124, 105–115. [Google Scholar] [CrossRef]
- Fritz, I.; Wagner, P.; Bottai, M.; Eriksson, H.; Ingvar, C.; Krakowski, I.; Nielsen, K.; Olsson, H. Desloratadine and loratadine use associated with improved melanoma survival. Allergy 2020, 75, 2096–2099. [Google Scholar] [CrossRef]
- Fritz, I.; Wagner, P.; Broberg, P.; Einefors, R.; Olsson, H. Desloratadine and loratadine stand out among common H(1)-antihistamines for association with improved breast cancer survival. Acta Oncol. 2020, 59, 1103–1109. [Google Scholar] [CrossRef] [PubMed]
- Ellegaard, A.M.; Dehlendorff, C.; Vind, A.C.; Anand, A.; Cederkvist, L.; Petersen, N.H.T.; Nylandsted, J.; Stenvang, J.; Mellemgaard, A.; Østerlind, K.; et al. Repurposing Cationic Amphiphilic Antihistamines for Cancer Treatment. EBioMedicine 2016, 9, 130–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafarzadeh, A.; Nemati, M.; Khorramdelazad, H.; Hassan, Z.M. Immunomodulatory properties of cimetidine: Its therapeutic potentials for treatment of immune-related diseases. Int. Immunopharmacol. 2019, 70, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Richter, J.M.; Colditz, G.A.; Huse, D.M.; Delea, T.E.; Oster, G. Cimetidine and adverse reactions: A meta-analysis of randomized clinical trials of short-term therapy. Am. J. Med. 1989, 87, 278–284. [Google Scholar] [CrossRef]
- Taylor, T.V.; Boom, S.J.; Blower, A.L.; McMahon, R.F.; Lawler, W. Healing of a malignant gastric ulcer with cimetidine. J. R. Coll. Surg. Edinb. 1988, 33, 339–340. [Google Scholar]
- Bolton, E.; King, J.; Morris, D.L. H2-antagonists in the treatment of colon and breast cancer. Semin. Cancer Biol. 2000, 10, 3–10. [Google Scholar] [CrossRef]
- Losurdo, G.; Principi, M.; Girardi, B.; Pricci, M.; Barone, M.; Ierardi, E.; Di Leo, A. Histamine and Histaminergic Receptors in Colorectal Cancer: From Basic Science to Evidence-based Medicine. Anti-Cancer Agents Med. Chem. 2018, 18, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Fujiwara, H.; Ueda, Y.; Itoh, T.; Yamashita, T.; Yoshimura, T.; Okugawa, K.; Yamamoto, Y.; Yano, Y.; Yamagishi, H. Cimetidine modulates the antigen presenting capacity of dendritic cells from colorectal cancer patients. Br. J. Cancer 2002, 86, 1257–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanaka, S.; Ichikawa, A. Effect of cimetidine on intratumoral cytokine expression in an experimental tumor. Biochem. Biophys. Res. Commun. 2001, 281, 1113–1119. [Google Scholar] [CrossRef]
- Kotake, T.; Kinouchi, T.; Saiki, S.; Kuroda, M.; Miki, T.; Kiyohara, H.; Usami, M. Treatment of metastatic renal cell carcinoma with a combination of human lymphoblastoid interferon-alpha and cimetidine. Jpn. J. Clin. Oncol. 1991, 21, 46–51. [Google Scholar]
- Walker, P.R.; Khuder, S.A.; Quan, W.D., Jr. Continuous infusion interleukin-2 and antihistamines in metastatic kidney cancer. Cancer Biother. Radiopharm. 2005, 20, 487–490. [Google Scholar] [CrossRef]
- Wu, X.X.; Mizutani, Y.; Kakehi, Y.; Nakamura, E.; Mitsumori, K.; Takahashi, T.; Terachi, T.; Okada, Y.; Yoshida, O. Expression of major histocompatibility complex antigens and adhesion molecules on renal cell carcinoma cells, and effect of interferon-alpha and/or cimetidine on the expression. Hinyokika Kiyo. Acta Urol. Jpn. 1998, 44, 621–626. [Google Scholar]
- Takahashi, H.K.; Watanabe, T.; Yokoyama, A.; Iwagaki, H.; Yoshino, T.; Tanaka, N.; Nishibori, M. Cimetidine induces interleukin-18 production through H2-agonist activity in monocytes. Mol. Pharmacol. 2006, 70, 450–453. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, I.; Chu, T.M. Synergistic antitumor activity of interleukin-2 and cimetidine against syngeneic murine tumor. Cancer Immunol. Immunother. 1991, 33, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.L.; Adams, W.J. Cimetidine and colorectal cancer--old drug, new use? Nat. Med. 1995, 1, 1243–1244. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.L.; Miatkowski, K.; Sizing, I.; Griffiths, D.; Zafari, M.; Benjamin, C.D.; Meier, W.; Mackay, F. Signaling through the lymphotoxin beta receptor induces the death of some adenocarcinoma tumor lines. J. Exp. Med. 1996, 183, 867–878. [Google Scholar] [CrossRef] [Green Version]
- Hazama, S.; Noma, T.; Wang, F.; Iizuka, N.; Ogura, Y.; Yoshimura, K.; Inoguchi, E.; Hakozaki, M.; Hirose, K.; Suzuki, T.; et al. Tumour cells engineered to secrete interleukin-15 augment anti-tumour immune responses in vivo. Br. J. Cancer 1999, 80, 1420–1426. [Google Scholar] [CrossRef] [Green Version]
- Hock, H.; Dorsch, M.; Kunzendorf, U.; Qin, Z.; Diamantstein, T.; Blankenstein, T. Mechanisms of rejection induced by tumor cell-targeted gene transfer of interleukin 2, interleukin 4, interleukin 7, tumor necrosis factor, or interferon gamma. Proc. Natl. Acad. Sci. USA 1993, 90, 2774–2778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parshad, R.; Kapoor, S.; Gupta, S.D.; Kumar, A.; Chattopadhyaya, T.K. Does famotidine enhance tumor infiltrating lymphocytes in breast cancer? Results of a randomized prospective pilot study. Acta Oncol. 2002, 41, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Deva, S.; Jameson, M. Histamine type 2 receptor antagonists as adjuvant treatment for resected colorectal cancer. Cochrane Database Syst. Rev. 2012, 8, Cd007814. [Google Scholar] [CrossRef]
- Ravelli, A.; Roviello, G.; Cretella, D.; Cavazzoni, A.; Biondi, A.; Cappelletti, M.R.; Zanotti, L.; Ferrero, G.; Ungari, M.; Zanconati, F.; et al. Tumor-infiltrating lymphocytes and breast cancer: Beyond the prognostic and predictive utility. Tumour Biol. 2017, 39, 1010428317695023. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, S.; Pal, S.; Sahni, P.; Dattagupta, S.; Kanti Chattopadhyay, T. Effect of pre-operative short course famotidine on tumor infiltrating lymphocytes in colorectal cancer: A double blind, placebo controlled, prospective randomized study. J. Surg. Res. 2005, 129, 172–175. [Google Scholar] [CrossRef]
- Adams, W.J.; Morris, D.L. Pilot study--cimetidine enhances lymphocyte infiltration of human colorectal carcinoma: Results of a small randomized control trial. Cancer 1997, 80, 15–21. [Google Scholar] [CrossRef]
- Nielsen, H.J.; Moesgaard, F.; Hammer, J.H. Effect of ranitidine and low-dose interleukin-2 in vitro on NK-cell activity in peripheral blood from patients with liver metastases from colorectal cancer. Eur. J. Surg. Oncol. 1995, 21, 526–530. [Google Scholar] [CrossRef]
- Lin, C.Y.; Bai, D.J.; Yuan, H.Y.; Wang, K.; Yang, G.L.; Hu, M.B.; Wu, Z.Q.; Li, Y. Perioperative cimetidine administration promotes peripheral blood lymphocytes and tumor infiltrating lymphocytes in patients with gastrointestinal cancer: Results of a randomized controlled clinical trial. World J. Gastroenterol. 2004, 10, 136–142. [Google Scholar] [CrossRef]
- Parshad, R.; Hazrah, P.; Kumar, S.; Gupta, S.D.; Ray, R.; Bal, S. Effect of preoperative short course famotidine on TILs and survival in breast cancer. Indian J. Cancer 2005, 42, 185–190. [Google Scholar]
- Kennedy, L.; Hodges, K.; Meng, F.; Alpini, G.; Francis, H. Histamine and histamine receptor regulation of gastrointestinal cancers. Transl. Gastrointest. Cancer 2012, 1, 215–227. [Google Scholar]
- Adams, W.J.; Lawson, J.A.; Morris, D.L. Cimetidine inhibits in vivo growth of human colon cancer and reverses histamine stimulated in vitro and in vivo growth. Gut 1994, 35, 1632–1636. [Google Scholar] [CrossRef]
- Jiang, C.G.; Liu, F.R.; Yu, M.; Li, J.B.; Xu, H.M. Cimetidine induces apoptosis in gastric cancer cells in vitro and inhibits tumor growth in vivo. Oncol. Rep. 2010, 23, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Dana, P.; Vaeteewoottacharn, K.; Kariya, R.; Matsuda, K.; Wongkham, S.; Okada, S. Repurposing cimetidine for cholangiocarcinoma: Antitumor effects in vitro and in vivo. Oncol. Lett. 2017, 13, 1432–1436. [Google Scholar] [CrossRef] [Green Version]
- Fujikawa, T.; Shiraha, H.; Nakanishi, Y.; Takaoka, N.; Ueda, N.; Suzuki, M.; Shiratori, Y. Cimetidine inhibits epidermal growth factor-induced cell signaling. J. Gastroenterol. Hepatol. 2007, 22, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Hirst, G.; Arumugam, P.J.; Beynon, J. Ranitidine as adjuvant treatment in colorectal cancer (Br J Surg 2002; 89: 1416-1422). Br. J. Surg. 2003, 90, 249. [Google Scholar] [CrossRef]
- Papagerakis, S.; Bellile, E.; Peterson, L.A.; Pliakas, M.; Balaskas, K.; Selman, S.; Hanauer, D.; Taylor, J.M.; Duffy, S.; Wolf, G. Proton pump inhibitors and histamine 2 blockers are associated with improved overall survival in patients with head and neck squamous carcinoma. Cancer Prev. Res. 2014, 7, 1258–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vila-Leahey, A.; Rogers, D.; Marshall, J.S. The impact of ranitidine on monocyte responses in the context of solid tumors. Oncotarget 2016, 7, 10891–10904. [Google Scholar] [CrossRef]
- Mathes, R.W.; Malone, K.E.; Daling, J.R.; Porter, P.L.; Li, C.I. Relationship between histamine2-receptor antagonist medications and risk of invasive breast cancer. Cancer Epidemiol. Biomark. Prev. 2008, 17, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Okegawa, T.; Pong, R.C.; Li, Y.; Hsieh, J.T. The role of cell adhesion molecule in cancer progression and its application in cancer therapy. Acta Biochim. Pol. 2004, 51, 445–457. [Google Scholar] [CrossRef] [Green Version]
- Borentain, P.; Carmona, S.; Mathieu, S.; Jouve, E.; El-Battari, A.; Gérolami, R. Inhibition of E-selectin expression on the surface of endothelial cells inhibits hepatocellular carcinoma growth by preventing tumor angiogenesis. Cancer Chemother. Pharmacol. 2016, 77, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Matsumoto, S.; Morishima, T.; Kawabe, T.; Okamoto, T. Cimetidine inhibits cancer cell adhesion to endothelial cells and prevents metastasis by blocking E-selectin expression. Cancer Res. 2000, 60, 3978–3984. [Google Scholar]
- Liu, F.R.; Jiang, C.G.; Li, Y.S.; Li, J.B.; Li, F. Cimetidine inhibits the adhesion of gastric cancer cells expressing high levels of sialyl Lewis x in human vascular endothelial cells by blocking E-selectin expression. Int. J. Mol. Med. 2011, 27, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Izumi, K.; Okabe, S. Roxatidine- and cimetidine-induced angiogenesis inhibition suppresses growth of colon cancer implants in syngeneic mice. J. Pharmacol. Sci. 2003, 93, 321–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cianchi, F.; Cortesini, C.; Schiavone, N.; Perna, F.; Magnelli, L.; Fanti, E.; Bani, D.; Messerini, L.; Fabbroni, V.; Perigli, G.; et al. The role of cyclooxygenase-2 in mediating the effects of histamine on cell proliferation and vascular endothelial growth factor production in colorectal cancer. Clin. Cancer Res. 2005, 11, 6807–6815. [Google Scholar] [CrossRef] [Green Version]
- Chihara, Y.; Fujimoto, K.; Miyake, M.; Hiasa, Y.; Hirao, Y. Anti-tumor effect of cimetidine via inhibiting angiogenesis factors in N-butyl-N-(4-hydroxybutyl) nitrosamine-induced mouse and rat bladder carcinogenesis. Oncol. Rep. 2009, 22, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlicker, E.; Kathmann, M. Role of the Histamine H(3) Receptor in the Central Nervous System. In Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2017; Volume 241, pp. 277–299. [Google Scholar] [CrossRef]
- Yu, D.; Zhao, J.; Wang, Y.; Hu, J.; Zhao, Q.; Li, J.; Zhu, J. Upregulated histamine receptor H3 promotes tumor growth and metastasis in hepatocellular carcinoma. Oncol. Rep. 2019, 41, 3347–3354. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Jia, J.; Zhang, J.J.; Xun, Y.P.; Xie, S.J.; Liang, J.F.; Guo, H.G.; Zhu, J.Z.; Ma, S.L.; Zhang, S.R. Inhibition of histamine receptor H3 suppresses the growth and metastasis of human non-small cell lung cancer cells via inhibiting PI3K/Akt/mTOR and MEK/ERK signaling pathways and blocking EMT. Acta Pharmacol. Sin. 2020, 42, 1288–1297. [Google Scholar] [CrossRef]
- Tanaka, T.; Kochi, T.; Shirakami, Y.; Mori, T.; Kurata, A.; Watanabe, N.; Moriwaki, H.; Shimizu, M. Cimetidine and Clobenpropit Attenuate Inflammation-Associated Colorectal Carcinogenesis in Male ICR Mice. Cancers 2016, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Francis, H.; Onori, P.; Gaudio, E.; Franchitto, A.; DeMorrow, S.; Venter, J.; Kopriva, S.; Carpino, G.; Mancinelli, R.; White, M.; et al. H3 histamine receptor-mediated activation of protein kinase Calpha inhibits the growth of cholangiocarcinoma in vitro and in vivo. Mol. Cancer Res. 2009, 7, 1704–1713. [Google Scholar] [CrossRef] [Green Version]
- Thurmond, R.L.; Venable, J.; Savall, B.; La, D.; Snook, S.; Dunford, P.J.; Edwards, J.P. Clinical Development of Histamine H(4) Receptor Antagonists. In Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2017; Volume 241, pp. 301–320. [Google Scholar] [CrossRef]
- Huang, J.F.; Thurmond, R.L. The new biology of histamine receptors. Curr. Allergy Asthma Rep. 2008, 8, 21–27. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massari, N.A.; Medina, V.A.; Martinel Lamas, D.J.; Cricco, G.P.; Croci, M.; Sambuco, L.; Bergoc, R.M.; Rivera, E.S. Role of H4 receptor in histamine-mediated responses in human melanoma. Melanoma Res. 2011, 21, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Coruzzi, G.; Adami, M.; Pozzoli, C. Role of histamine H4 receptors in the gastrointestinal tract. Front. Biosci. 2012, 4, 226–239. [Google Scholar] [CrossRef]
- Martinel Lamas, D.J.; Rivera, E.S.; Medina, V.A. Histamine H4 receptor: Insights into a potential therapeutic target in breast cancer. Front. Biosci. 2015, 7, 1–9. [Google Scholar]
- Sterle, H.A.; Nicoud, M.B.; Massari, N.A.; Táquez Delgado, M.A.; Herrero Ducloux, M.V.; Cremaschi, G.A.; Medina, V.A. Immunomodulatory role of histamine H4 receptor in breast cancer. Br. J. Cancer 2019, 120, 128–138. [Google Scholar] [CrossRef]
- Deiteren, A.; De Man, J.G.; Pelckmans, P.A.; De Winter, B.Y. Histamine H4 receptors in the gastrointestinal tract. Br. J. Pharmacol. 2015, 172, 1165–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, V.A.; Brenzoni, P.G.; Lamas, D.J.; Massari, N.; Mondillo, C.; Nunez, M.A.; Pignataro, O.; Rivera, E.S. Role of histamine H4 receptor in breast cancer cell proliferation. Front. Biosci. 2011, 3, 1042–1060. [Google Scholar] [CrossRef]
- He, G.H.; Ding, J.Q.; Zhang, X.; Xu, W.M.; Lin, X.Q.; Huang, M.J.; Feng, J.; Wang, P.; Cai, W.K. Activation of histamine H(4) receptor suppresses the proliferation and invasion of esophageal squamous cell carcinoma via both metabolism and non-metabolism signaling pathways. J. Mol. Med. 2018, 96, 951–964. [Google Scholar] [CrossRef]
- Fang, Z.; Yao, W.; Xiong, Y.; Li, J.; Liu, L.; Shi, L.; Zhang, W.; Zhang, C.; Nie, L.; Wan, J. Attenuated expression of HRH4 in colorectal carcinomas: A potential influence on tumor growth and progression. BMC Cancer 2011, 11, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Xiong, Y.; Li, J.; Yang, Y.; Liu, L.; Wang, W.; Wang, L.; Li, M.; Fang, Z. Deletion and down-regulation of HRH4 gene in gastric carcinomas: A potential correlation with tumor progression. PLoS ONE 2012, 7, e31207. [Google Scholar] [CrossRef]
- Paik, W.H.; Ryu, J.K.; Jeong, K.S.; Park, J.M.; Song, B.J.; Lee, S.H.; Kim, Y.T.; Yoon, Y.B. Clobenpropit enhances anti-tumor effect of gemcitabine in pancreatic cancer. World J. Gastroenterol. 2014, 20, 8545–8557. [Google Scholar] [CrossRef] [PubMed]
- Massari, N.A.; Medina, V.A.; Cricco, G.P.; Martinel Lamas, D.J.; Sambuco, L.; Pagotto, R.; Ventura, C.; Ciraolo, P.J.; Pignataro, O.; Bergoc, R.M.; et al. Antitumor activity of histamine and clozapine in a mouse experimental model of human melanoma. J. Dermatol. Sci. 2013, 72, 252–262. [Google Scholar] [CrossRef]
- Massari, N.A.; Nicoud, M.B.; Sambuco, L.; Cricco, G.P.; Martinel Lamas, D.J.; Herrero Ducloux, M.V.; Blanco, H.; Rivera, E.S.; Medina, V.A. Histamine therapeutic efficacy in metastatic melanoma: Role of histamine H4 receptor agonists and opportunity for combination with radiation. Oncotarget 2017, 8, 26471–26491. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Han, Y.; Staloch, D.; Francis, T.; Stokes, A.; Francis, H. The H4 histamine receptor agonist, clobenpropit, suppresses human cholangiocarcinoma progression by disruption of epithelial mesenchymal transition and tumor metastasis. Hepatology 2011, 54, 1718–1728. [Google Scholar] [CrossRef]
- Cai, W.K.; Hu, J.; Li, T.; Meng, J.R.; Ma, X.; Yin, S.J.; Zhao, C.H.; He, G.H.; Xu, G.L. Activation of histamine H4 receptors decreases epithelial-to-mesenchymal transition progress by inhibiting transforming growth factor-β1 signalling pathway in non-small cell lung cancer. Eur. J. Cancer 2014, 50, 1195–1206. [Google Scholar] [CrossRef]
- Bakir, B.; Chiarella, A.M.; Pitarresi, J.R.; Rustgi, A.K. EMT, MET, Plasticity, and Tumor Metastasis. Trends Cell Biol. 2020, 30, 764–776. [Google Scholar] [CrossRef]
- Gonzalez-Avila, G.; Sommer, B.; García-Hernández, A.A.; Ramos, C. Matrix Metalloproteinases’ Role in Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1245, 97–131. [Google Scholar] [CrossRef] [PubMed]
- Rosethorne, E.M.; Charlton, S.J. Agonist-biased signaling at the histamine H4 receptor: JNJ7777120 recruits β-arrestin without activating G proteins. Mol. Pharmacol. 2011, 79, 749–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoud, M.B.; Táquez Delgado, M.A.; Sarasola, M.P.; Vidal, A.; Speisky, D.; Cremaschi, G.A.; Sterle, H.A.; Medina, V.A. Impact of histamine H4 receptor deficiency on the modulation of T cells in a murine breast cancer model. Cancer Immunol. Immunother. 2020, 70, 233–244. [Google Scholar] [CrossRef]
- Nicoud, M.B.; Sterle, H.A.; Massari, N.A.; Táquez Delgado, M.A.; Formoso, K.; Herrero Ducloux, M.V.; Martinel Lamas, D.; Cremaschi, G.A.; Medina, V.A. Study of the antitumour effects and the modulation of immune response by histamine in breast cancer. Br. J. Cancer 2020, 122, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Shao, Y.; Zhang, J.; Dong, X.L.; Liu, W.L.; Yang, H.; Liu, L.; Li, M.H.; Yue, C.F.; Fang, Z.Y.; et al. Polymorphisms in human histamine receptor H4 gene are associated with atopic dermatitis. Br. J. Dermatol. 2010, 162, 1038–1043. [Google Scholar] [CrossRef]
- Simon, T.; Semsei, A.F.; Ungvári, I.; Hadadi, E.; Virág, V.; Nagy, A.; Vangor, M.S.; László, V.; Szalai, C.; Falus, A. Asthma endophenotypes and polymorphisms in the histamine receptor HRH4 gene. Int. Arch. Allergy Immunol. 2012, 159, 109–120. [Google Scholar] [CrossRef]
- He, G.H.; Lu, J.; Shi, P.P.; Xia, W.; Yin, S.J.; Jin, T.B.; Chen, D.D.; Xu, G.L. Polymorphisms of human histamine receptor H4 gene are associated with breast cancer in Chinese Han population. Gene 2013, 519, 260–265. [Google Scholar] [CrossRef]
- Cai, W.K.; Zhang, J.B.; Chen, J.H.; Meng, J.R.; Ma, X.; Zhang, J.; Zhou, Y.L.; Xu, G.L.; He, G.H. The HRH4 rs11662595 mutation is associated with histamine H(4) receptor dysfunction and with increased epithelial-to-mesenchymal transition progress in non-small cell lung cancer. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2954–2963. [Google Scholar] [CrossRef]
- van Rijn, R.M.; van Marle, A.; Chazot, P.L.; Langemeijer, E.; Qin, Y.; Shenton, F.C.; Lim, H.D.; Zuiderveld, O.P.; Sansuk, K.; Dy, M.; et al. Cloning and characterization of dominant negative splice variants of the human histamine H4 receptor. Biochem. J. 2008, 414, 121–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| HR Subtype | Distribution | Agonist | Antagonist/Inverse Agonist |
|---|---|---|---|
| H1R | B cells, cancer cells, chondrocyte, dendritic cells, endothelial cells, eosinophils, hepatocytes, mast cells, monocytes, neutrophils, nerve cells, smooth muscle, and T cells | Histaprofiden, methylhistaprodifen, and suprahistaprodifen | Astemizole, cetirizine, chlorpheniramine, cyproheptadine, diphenhydramine, fexofenadine, loratadine, mepyramine, pheniramine, pyrilamine, terfenadine, and triprolidine |
| H2R | B cells, cancer cells, chondrocyte, dendritic cells, endothelial cells, eosinophils, epithelial cells, gastric parietal cells, hepatocytes, mast cells, monocytes, neutrophils, nerve cells, smooth muscle, and T cells | Amthamine, dimaprit, and impromidine | Burimamide, cimetidine, famotidine, lafutidine, ranitidine, and tiotidine |
| H3R | Cancer cells, eosinophils, histaminergic neurons, and monocytes | Imetit, immepip, immethridine, and R-α-(-)methylhistamine | Clobenpropit, ciproxifan, JNJ5207852, JNJ10181457, OUP-186, pitolisant, and thioperamide |
| H4R | Basophils, cancer cells, dendritic cells, hematopoietic cells, hepatocytes, leukocytes, mast cells, monocytes, neutrophils, and T cells | Clobenpropit, JNJ28610244, 4-Methylhistamine, ST-1006, VUF 6884, and VUF 8430 | A-940894, A-987306, JNJ7777120, JNJ10191584, JNJ39758979, thioperamide, VUF 6002, and ZPL-3893787 |
| Cancer Type | Experimental Models | H1R Antagonists | Effects | References |
|---|---|---|---|---|
| HCC | SNU-368 | Terfenadine | ↓ proliferation ↓ migration ↓ invasion | [126] |
| SNU-368 xenograft mice | ↓ tumor growth ↓ metastasis | |||
| BC | Basal and HER2-targeted therapy-resistant cells | Terfenadine | ↓ proliferation ↑ apoptosis | [129] |
| MDA-MB-231 MCF-7 | Chlorpheniramine | ↓ cell number | [130] | |
| MDA-MB-231 T-47D xenograft mice | Terfenadine | ↓ tumor growth | [129] | |
| Human | Desloratadine Loratadine | ↑ survival rate | [140] | |
| NSCLC | A549 | Terfinadine | ↓ migration ↓ invasion | [138] |
| A549 xenograft mice | ↓ tumor growth ↓ metastasis | |||
| NSCLC | CAD | ↓ mortality | [141] | |
| MM | A375 HT144 HSs294T | Terfenadine | ↑ apoptosis | [131] |
| A2058 A375 | Diphenhydramine | ↑ apoptosis | [133] | |
| B16F10 xenograft mice | ↓ tumor growth ↑ survival time | |||
| B16F10 A375 syngeneic mice | Terfenadine | ↓ tumor growth | [32] | |
| Human | Desloratadine Loratadine | ↑ survival rate | [139] | |
| Leukemia | CCRF-CEM Jurkat | Diphenhydramine | ↑ apoptosis | [134] |
| OC | OVCAR-3 UWB1-289 OCV-316 | CAD | ↑ cell death | [135] |
| Human | CAD | ↓ cancer mortality | ||
| Prostate cancer | PC-3 DU-145 | Terfenadine | ↓ proliferation ↑ apoptosis | [136] |
| Cancer Type | Experimental Models | H4R Agonists | Effects | References |
|---|---|---|---|---|
| BC | MDA-MB-231 | Histamine Clozapine JNJ28610244 | ↓ proliferation ↑ apoptosis | [37] |
| MDA-MB-231 MCF-7 | Clobenpropit VUF 8430 | ↓ proliferation ↑ apoptosis ↑ senescence | [195] | |
| MDA-MB-231 xenograft mice | Histamine Clozapine JNJ28610244 | ↓ tumor growth ↓ angiogenesis ↓ metastasis | [37] | |
| 4T1 syngeneic mice | Histamine | ↓ tumor growth | [208] | |
| MM | WM35 M1/15 | Clobenpropit VUF 8430 | ↓ proliferation ↑ senescence | [190] |
| M1/15 xenograft mice | Clozapine | ↓ tumor growth ↑ survival time | [200] | |
| 1205Lu xenograft mice | Histamine | ↓ metastasis | [201] | |
| 1205Lu | Histamine plus ionizing radiation | ↓ proliferation ↑ apoptosis | ||
| ESCC | TE-2 | 4-Methylhistamine | ↓ proliferation ↓ invasion ↓ metastasis | [196] |
| TE-2 xenograft mice | 4-Methylhistamine | ↓ tumor growth ↑ survival time | ||
| CRC | Colo-320 Lovo | Clozapine | ↓proliferation ↑ cell cycle arrest ↑ apoptosis | [197] |
| GC | AGS | Clobenpropit 4-Methylhistamine | ↓ proliferation ↑ cell cycle arrest | [198] |
| PC | Panc-1 | Clobenpropit | ↑ apoptosis ↓ migration | [199] |
| Panc-1 xenograft mice | Clobenpropit | ↓ tumor growth | ||
| CCA | Mz-ChA-1 SG231 HuH-28 TFK-1 HuCCT-1 CCLP1 | Clobenpropit | ↓ proliferation ↑ apoptosis | [202] |
| Mz-ChA-1 | Clobenpropit | ↓ proliferation ↓ invasion, ↓ migration | ||
| NSCLC | H157 H460 A549 H322 | 4-Methylhistamine | ↓ invasion ↓ metastasis | [203] |
| A549 xenograft mice | 4-Methylhistamine | ↓ tumor growth ↑ survival time |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, P.L.; Cho, J. Pathophysiological Roles of Histamine Receptors in Cancer Progression: Implications and Perspectives as Potential Molecular Targets. Biomolecules 2021, 11, 1232. https://doi.org/10.3390/biom11081232
Nguyen PL, Cho J. Pathophysiological Roles of Histamine Receptors in Cancer Progression: Implications and Perspectives as Potential Molecular Targets. Biomolecules. 2021; 11(8):1232. https://doi.org/10.3390/biom11081232
Chicago/Turabian StyleNguyen, Phuong Linh, and Jungsook Cho. 2021. "Pathophysiological Roles of Histamine Receptors in Cancer Progression: Implications and Perspectives as Potential Molecular Targets" Biomolecules 11, no. 8: 1232. https://doi.org/10.3390/biom11081232
APA StyleNguyen, P. L., & Cho, J. (2021). Pathophysiological Roles of Histamine Receptors in Cancer Progression: Implications and Perspectives as Potential Molecular Targets. Biomolecules, 11(8), 1232. https://doi.org/10.3390/biom11081232