
Growth Suppression in Lung Cancer Cells Harboring EGFR-C797S Mutation by Quercetin

 ,
,
Abstract
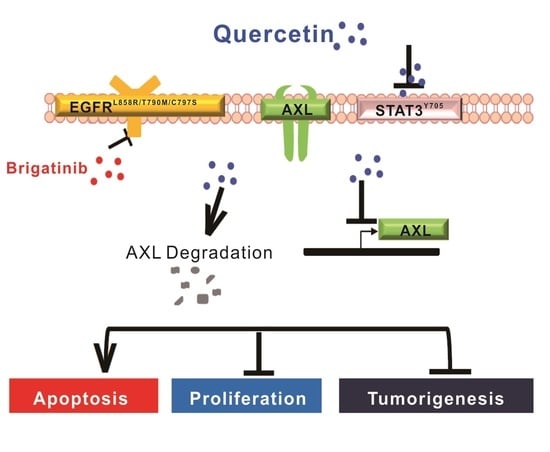
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Culture Media, Reagents, and Antibodies
2.2. Cell Line and Culture
2.3. Plasmids and Transfection
2.4. Cell Viability Assays
2.5. Colony Formation Assay
2.6. Apoptosis Assays
2.7. Western Blot Analysis
2.8. Real-Time Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
2.9. Xenograft Mouse Model
2.10. Immunohistochemistry (IHC)
2.11. Molecular Docking Analysis
2.12. Statistical Analysis
3. Results
3.1. Effects of Quercetin on the Viability and Growth of Human NSCLC Cells
3.2. Effects of Quercetin on the Induction of Apoptosis and Autophagy in NSCLC Cells
3.3. Quercetin Downregulates the Expression of AXL in EGFR-TKI-Resistant Cells
3.4. The Effects of Quercetin and Brigatinib on the Growth of H1975-MS35 Tumor Cells In Vitro and In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Sequist, L.V.; Yang, J.C.-H.; Yamamoto, N.; Obyrne, K.J.; Hirsh, V.; Mok, T.; Geater, S.L.; Orlov, S.; Tsai, C.-M.; Boyer, M.; et al. Phase III Study of Afatinib or Cisplatin Plus Pemetrexed in Patients With Metastatic Lung Adenocarcinoma With EGFR Mutations. J. Clin. Oncol. 2013, 31, 3327–3334. [Google Scholar] [CrossRef] [Green Version]
- Oxnard, G.R.; Binder, A.; Jänne, P.A. New Targetable Oncogenes in Non–Small-Cell Lung Cancer. J. Clin. Oncol. 2013, 31, 1097–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, C.R.; Jänne, P.A. The quest to overcome resistance to EGFR-targeted therapies in cancer. Nat. Med. 2013, 19, 1389–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gainor, J.F.; Shaw, A.T. Emerging Paradigms in the Development of Resistance to Tyrosine Kinase Inhibitors in Lung Cancer. J. Clin. Oncol. 2013, 31, 3987–3996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thress, K.S.; Paweletz, C.P.; Felip, E.; Cho, B.C.; Stetson, D.; Dougherty, B.; Lai, Z.; Markovets, A.; Vivancos, A.; Kuang, Y.; et al. Acquired EGFR C797S mutation mediates resistance to AZD9291 in non–small cell lung cancer harboring EGFR T790M. Nat. Med. 2015, 21, 560–562. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.-H.; Wu, C.-C.; Huang, K.-Y.; Leu, Y.-L.; Yang, S.-C.; Chen, C.-L.; Chen, C.-Y. Integrated Omics Analysis of Non-Small-Cell Lung Cancer Cells Harboring the EGFR C797S Mutation Reveals the Potential of AXL as a Novel Therapeutic Target in TKI-Resistant Lung Cancer. Cancers 2020, 13, 111. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Garg, V.K.; Tuli, H.S.; Yerer, M.B.; Sak, K.; Sharma, A.K.; Kumar, M.; Aggarwal, V.; Sandhu, S.S. Fisetin and Quercetin: Promising Flavonoids with Chemopreventive Potential. Biomolecules 2019, 9, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Gao, L.; Zhang, Y.; Simpson, B.K. Preparation of Quercetin Loaded Microparticles and their Antitumor Activity against Human Lung Cancer Cells (A549) in vitro. Curr. Pharm. Biotechnol. 2019, 20, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yuan, L.; Meng, L.; Qiu, J.; Wang, C. Quercetin-loaded mixed micelles exhibit enhanced cytotoxic efficacy in non-small cell lung cancer in vitro. Exp. Ther. Med. 2017, 14, 5503–5508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimaszewska-Wiśniewska, A.; Hałas-Wiśniewska, M.; Izdebska, M.; Gagat, M.; Grzanka, A.; Grzanka, D. Antiproliferative and antimetastatic action of quercetin on A549 non-small cell lung cancer cells through its effect on the cytoskeleton. Acta Histochem. 2017, 119, 99–112. [Google Scholar] [CrossRef]
- Chang, J.-H.; Lai, S.-L.; Chen, W.-S.; Hung, W.-Y.; Chow, J.-M.; Hsiao, M.; Lee, W.-J.; Chien, M.-H. Quercetin suppresses the metastatic ability of lung cancer through inhibiting Snail-dependent Akt activation and Snail-independent ADAM9 expression pathways. Biochim. Biophys. Acta Mol. Cell Research. 2017, 1864, 1746–1758. [Google Scholar] [CrossRef]
- Chuang, C.H.; Yeh, C.L.; Yeh, S.L.; Lin, E.S.; Wang, L.Y.; Wang, Y.H. Quercetin metabolites inhibit mmp-2 expression in a549 lung cancer cells by ppar-gamma associated mechanisms. J. Nutr. Biochem. 2016, 33, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Khuda-Bukhsh, A.R. Quercetin Down-regulates IL-6/STAT-3 Signals to Induce Mitochondrial-mediated Apoptosis in a Nonsmall- cell Lung-cancer Cell Line, A549. J. Pharmacopuncture 2015, 18, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.-Y.; Li, Y.; Jiang, N.; Zhao, J.; Ge, J.-F. Anticancer effect and apoptosis induction by quercetin in the human lung cancer cell line A-549. Mol. Med. Rep. 2011, 5, 822–826. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.I.; Lee, S.J.; Choi, Y.-J.; Kim, M.J.; Kim, T.Y.; Ko, S.-G. Quercetin Induces Apoptosis in Glioblastoma Cells by Suppressing Axl/IL-6/STAT3 Signaling Pathway. Am. J. Chin. Med. 2021, 49, 767–784. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.-H.; Lin, Y.-H.; Yang, S.-C.; Chang, P.-C.; Wang, T.-C.; Chen, C.-Y. Tid1-S regulates the mitochondrial localization of EGFR in non-small cell lung carcinoma. Oncogenesis 2017, 6, e361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-Y.; Jan, C.-I.; Lo, J.-F.; Yang, S.-C.; Chang, Y.-L.; Pan, S.-H.; Wang, W.-L.; Hong, T.-M.; Yang, P.-C. Tid1-L Inhibits EGFR Signaling in Lung Adenocarcinoma by Enhancing EGFR Ubiquitinylation and Degradation. Cancer Res. 2013, 73, 4009–4019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-Y.; Chen, C.-C.; Chuang, W.-Y.; Leu, Y.-L.; Ueng, S.-H.; Hsueh, C.; Yeh, C.-T.; Wang, T.-H. Hydroxygenkwanin Inhibits Class I HDAC Expression and Synergistically Enhances the Antitumor Activity of Sorafenib in Liver Cancer Cells. Front. Oncol. 2020, 10, 216. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chang, P.C.; Wang, T.H.; Wang, T.V. The in vivo anti-leukemia activity of n-(1-pyrenlyl) maleimide in a bioluminescent mouse model. Leuk. Res. 2017, 62, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.-M.; Chen, C.-Y.; Huang, P.-R.; Hsu, C.-W.; Wu, C.-C.; Wang, T.-C.V. Proteomic analyses of genes regulated by heterogeneous nuclear ribonucleoproteins A/B in Jurkat cells. Proteomics 2014, 14, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Leu, Y.-L.; Wang, T.-H.; Wu, C.-C.; Huang, K.-Y.; Jiang, Y.-W.; Hsu, Y.-C.; Chen, C.-Y. Hydroxygenkwanin Suppresses Non-Small Cell Lung Cancer Progression by Enhancing EGFR Degradation. Molecules 2020, 25, 941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-Y.; Jan, C.-I.; Pi, W.-C.; Wang, W.-L.; Yang, P.-C.; Wang, T.-H.; Karni, R.; Wang, T.-C.V. Heterogeneous nuclear ribonucleoproteins A1 and A2 modulate expression of Tid1 isoforms and EGFR signaling in non-small cell lung cancer. Oncotarget 2016, 7, 16760–16772. [Google Scholar] [CrossRef] [Green Version]
- Jan, C.-I.; Yu, C.-C.; Hung, M.-C.; Harn, H.-J.; Nieh, S.; Lee, H.-S.; Lou, M.A.; Wu, Y.-C.; Chen, C.-Y.; Huang, C.-Y.; et al. Tid1, CHIP and ErbB2 interactions and their prognostic implications for breast cancer patients. J. Pathol. 2011, 225, 424–437. [Google Scholar] [CrossRef] [PubMed]
- Tseng, W.-C.; Chen, C.-Y.; Chern, C.-Y.; Wang, C.-A.; Lee, W.-C.; Chi, Y.-C.; Cheng, S.-F.; Kuo, Y.-T.; Chiu, Y.-C.; Tseng, S.-T.; et al. Targeting HR Repair as a Synthetic Lethal Approach to Increase DNA Damage Sensitivity by a RAD52 Inhibitor in BRCA2-Deficient Cancer Cells. Int. J. Mol. Sci. 2021, 22, 4422. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.K.; DeRyckere, D.; Davies, K.D.; Earp, H.S. The TAM family: Phosphatidylserine-sensing receptor tyrosine kinases gone awry in cancer. Nat. Rev. Cancer 2014, 14, 769–785. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, L.; Yin, J.C.; Wu, X.; Shao, Y.W.; Gao, B. Lung Adenocarcinoma Harboring EGFR 19del/C797S/T790M Triple Mutations Responds to Brigatinib and Anti-EGFR Antibody Combination Therapy. J. Thorac. Oncol. 2019, 14, e85–e88. [Google Scholar] [CrossRef] [PubMed]
- Uchibori, K.; Inase, N.; Araki, M.; Kamada, M.; Sato, S.; Okuno, Y.; Fujita, N.; Katayama, R. Brigatinib combined with anti-EGFR antibody overcomes osimertinib resistance in EGFR-mutated non-small-cell lung cancer. Nat. Commun. 2017, 8, 14768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Ding, H.; Tang, X.; Liang, M.; Li, S.; Zhang, J.; Cao, J. Quercetin induces pro-apoptotic autophagy via SIRT1 / AMPK signaling pathway in human lung cancer cell lines A549 and H1299 in vitro. Thorac. Cancer 2021, 12, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Rankin, E.; Fuh, K.; Castellini, L.; Viswanathan, K.; Finger, E.C.; Diep, A.N.; LaGory, E.L.; Kariolis, M.S.; Chan, A.; Lindgren, D.; et al. Direct regulation of GAS6/AXL signaling by HIF promotes renal metastasis through SRC and MET. Proc. Natl. Acad. Sci. USA 2014, 111, 13373–13378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S. Polyphenol Compound as a Transcription Factor Inhibitor. Nutrients 2015, 7, 8987–9004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.W.; Hou, W.C.; Shen, S.C.; Juan, S.H.; Ko, C.H.; Wang, L.M.; Chen, Y.C. Quercetin inhibition of tumor invasion via suppressing pkc delta/erk/ap-1-dependent matrix metalloproteinase-9 activation in breast carcinoma cells. Carcinogenesis 2008, 29, 1807–1815. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Wannatung, T.; Lee, S.; Yang, W.K.; Chung, S.H.; Lim, J.-S.; Choe, W.; Kang, I.; Kim, S.-S.; Ha, J. Quercetin enhances hypoxia-mediated apoptosis via direct inhibition of AMPK activity in HCT116 colon cancer. Apoptosis 2012, 17, 938–949. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, K.-Y.; Wang, T.-H.; Chen, C.-C.; Leu, Y.-L.; Li, H.-J.; Jhong, C.-L.; Chen, C.-Y. Growth Suppression in Lung Cancer Cells Harboring EGFR-C797S Mutation by Quercetin. Biomolecules 2021, 11, 1271. https://doi.org/10.3390/biom11091271
Huang K-Y, Wang T-H, Chen C-C, Leu Y-L, Li H-J, Jhong C-L, Chen C-Y. Growth Suppression in Lung Cancer Cells Harboring EGFR-C797S Mutation by Quercetin. Biomolecules. 2021; 11(9):1271. https://doi.org/10.3390/biom11091271
Chicago/Turabian StyleHuang, Kuo-Yen, Tong-Hong Wang, Chin-Chuan Chen, Yann-Lii Leu, Hsin-Jung Li, Cai-Ling Jhong, and Chi-Yuan Chen. 2021. "Growth Suppression in Lung Cancer Cells Harboring EGFR-C797S Mutation by Quercetin" Biomolecules 11, no. 9: 1271. https://doi.org/10.3390/biom11091271
APA StyleHuang, K. -Y., Wang, T. -H., Chen, C. -C., Leu, Y. -L., Li, H. -J., Jhong, C. -L., & Chen, C. -Y. (2021). Growth Suppression in Lung Cancer Cells Harboring EGFR-C797S Mutation by Quercetin. Biomolecules, 11(9), 1271. https://doi.org/10.3390/biom11091271