
Endogenously-Produced Hyaluronan and Its Potential to Regulate the Development of Peritoneal Adhesions


 and
and 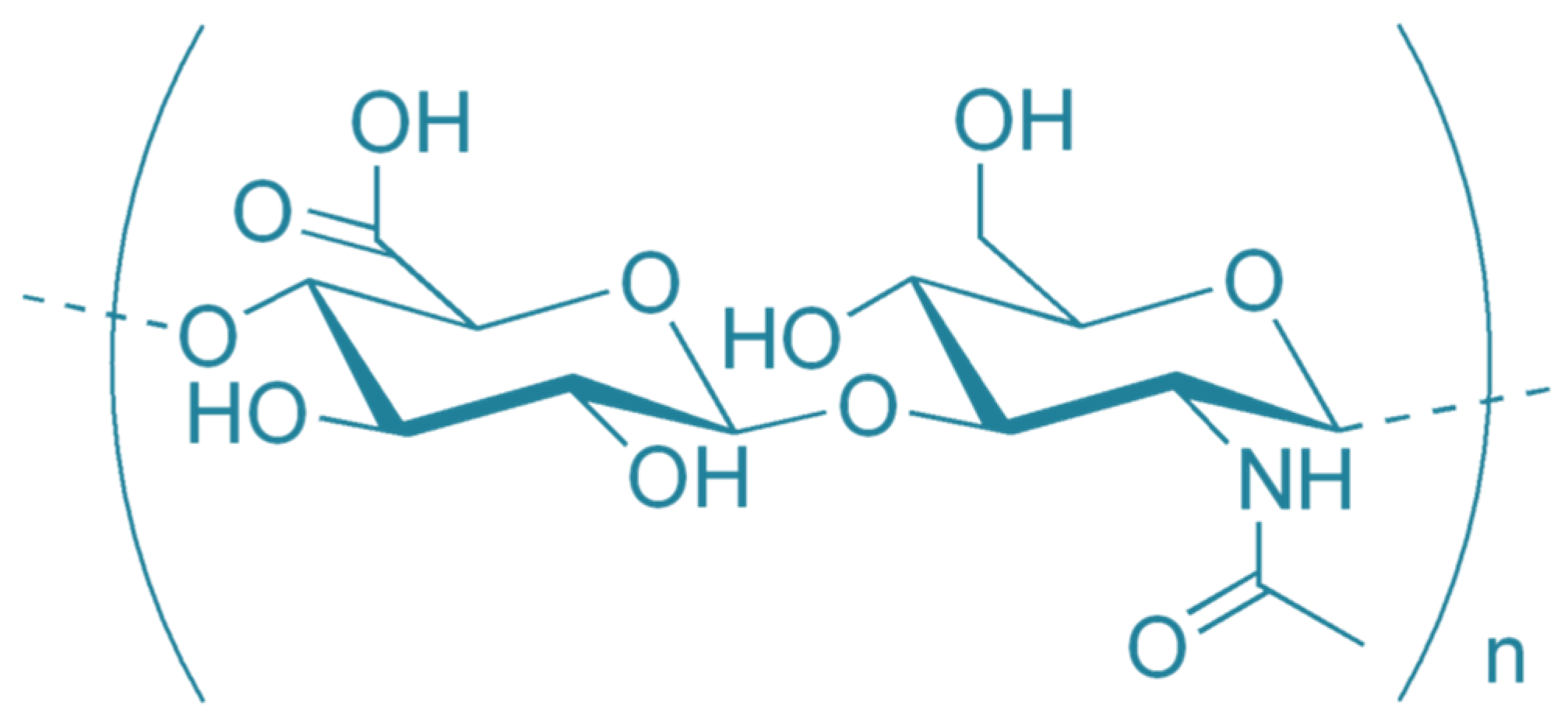{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Regulatory Role of ECM Components in Fibrosis
3. Hyaluronan (Hyaluronic Acid)
3.1. Metabolism of HA
3.1.1. HA Synthesis
3.1.2. HA Degradation
3.2. Role of HA in Its Physiological Functions
3.2.1. HA Fragmentation and Changes in Biological Properties
3.2.2. HA Supramolecular Complexes
3.3. HA and Energetic Metabolism
4. HA in Peritoneal Fibrosis
4.1. Role of Endogenous HA in Peritoneal Fibrosis
4.2. Cues from Other Tissues
4.3. Anti-Fibrotic Effects of Endogenous HA
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brochhausen, C.; Schmitt, V.H.; Planck, C.N.; Rajab, T.K.; Hollemann, D.; Tapprich, C.; Kramer, B.; Wallwiener, C.; Hierlemann, H.; Zehbe, R.; et al. Current strategies and future perspectives for intraperitoneal adhesion prevention. J. Gastrointest Surg. 2012, 16, 1256–1274. [Google Scholar] [CrossRef]
- Canis, M.; Botchorishvili, R.; Bourdel, N.; Gremeau, A.S.; Curinier, S.; Rabischong, B. Pelvic adhesions and fertility: Where are we in 2018? J. Visc. Surg. 2018, 155 (Suppl. 1), S11–S15. [Google Scholar] [CrossRef]
- Koninckx, P.R.; Gomel, V.; Ussia, A.; Adamyan, L. Role of the peritoneal cavity in the prevention of postoperative adhesions, pain, and fatigue. Fertil. Steril. 2016, 106, 998–1010. [Google Scholar] [CrossRef] [Green Version]
- Herrick, S.E.; Wilm, B. Post-Surgical Peritoneal Scarring and Key Molecular Mechanisms. Biomolecules 2021, 11, 692. [Google Scholar] [CrossRef]
- Zwicky, S.N.; Stroka, D.; Zindel, J. Sterile Injury Repair and Adhesion Formation at Serosal Surfaces. Front. Immunol. 2021, 12, 684967. [Google Scholar] [CrossRef]
- Capella-Monsonis, H.; Kearns, S.; Kelly, J.; Zeugolis, D.I. Battling adhesions: From understanding to prevention. BMC Biomed. Eng. 2019, 1, 5. [Google Scholar] [CrossRef] [Green Version]
- Manou, D.; Caon, I.; Bouris, P.; Triantaphyllidou, I.E.; Giaroni, C.; Passi, A.; Karamanos, N.K.; Vigetti, D.; Theocharis, A.D. The Complex Interplay Between Extracellular Matrix and Cells in Tissues. Methods Mol. Biol. 2019, 1952, 1–20. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Bulow, R.D.; Boor, P. Extracellular Matrix in Kidney Fibrosis: More Than Just a Scaffold. J. Histochem. Cytochem. 2019, 67, 643–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, M.W.; Rossi, D.; Peterson, M.; Smith, K.; Sikstrom, K.; White, E.S.; Connett, J.E.; Henke, C.A.; Larsson, O.; Bitterman, P.B. Fibrotic extracellular matrix activates a profibrotic positive feedback loop. J. Clin. Investig. 2014, 124, 1622–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricard-Blum, S. The collagen family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villesen, I.F.; Daniels, S.J.; Leeming, D.J.; Karsdal, M.A.; Nielsen, M.J. Review article: The signalling and functional role of the extracellular matrix in the development of liver fibrosis. Aliment Pharm. 2020, 52, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.B.; Chen, J.; Jin, X.L.; Huang, Y.; Su, Q.M.; Chen, L. Exosome-mediated peritoneal dissemination in gastric cancer and its clinical applications. Biomed. Rep. 2018, 8, 503–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, J.; Henke, C.A.; Bitterman, P.B. Extracellular matrix as a driver of progressive fibrosis. J. Clin. Investig. 2018, 128, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef] [PubMed]
- Vigetti, D.; Karousou, E.; Viola, M.; Deleonibus, S.; De Luca, G.; Passi, A. Hyaluronan: Biosynthesis and signaling. Biochim. Biophys. Acta 2014, 1840, 2452–2459. [Google Scholar] [CrossRef] [Green Version]
- Litwiniuk, M.; Krejner, A.; Speyrer, M.S.; Gauto, A.R.; Grzela, T. Hyaluronic Acid in Inflammation and Tissue Regeneration. Wounds A Compend. Clin. Res. Pract. 2016, 28, 78–88. [Google Scholar]
- Itano, N.; Sawai, T.; Yoshida, M.; Lenas, P.; Yamada, Y.; Imagawa, M.; Shinomura, T.; Hamaguchi, M.; Yoshida, Y.; Ohnuki, Y.; et al. Three isoforms of mammalian hyaluronan synthases have distinct enzymatic properties. J. Biol. Chem. 1999, 274, 25085–25092. [Google Scholar] [CrossRef] [Green Version]
- Vigetti, D.; Viola, M.; Karousou, E.; De Luca, G.; Passi, A. Metabolic control of hyaluronan synthases. Matrix Biol. J. Int. Soc. Matrix Biol. 2014, 35, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Heldin, P.; Lin, C.Y.; Kolliopoulos, C.; Chen, Y.H.; Skandalis, S.S. Regulation of hyaluronan biosynthesis and clinical impact of excessive hyaluronan production. Matrix Biol. J. Int. Soc. Matrix Biol. 2019, 78-79, 100–117. [Google Scholar] [CrossRef]
- Garantziotis, S.; Savani, R.C. Hyaluronan biology: A complex balancing act of structure, function, location and context. Matrix Biol. J. Int. Soc. Matrix. Biol. 2019, 78-79, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kaul, A.; Singampalli, K.L.; Parikh, U.M.; Yu, L.; Keswani, S.G.; Wang, X. Hyaluronan, a double-edged sword in kidney diseases. Pediatric Nephrol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.; Stern, R. Chain gangs: New aspects of hyaluronan metabolism. Biochem. Res. Int. 2012, 2012, 893947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, R. Hyaluronan catabolism: A new metabolic pathway. Eur. J. Cell Biol. 2004, 83, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Yamamoto, H.; Tobisawa, Y.; Irie, F. TMEM2: A missing link in hyaluronan catabolism identified? Matrix Biol. J. Int. Soc. Matrix Biol. 2019, 78–79, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Tobisawa, Y.; Fujita, N.; Yamamoto, H.; Ohyama, C.; Irie, F.; Yamaguchi, Y. The cell surface hyaluronidase TMEM2 is essential for systemic hyaluronan catabolism and turnover. J. Biol. Chem. 2021, 297, 101281. [Google Scholar] [CrossRef]
- Yamazaki, K.; Fukuda, K.; Matsukawa, M.; Hara, F.; Yoshida, K.; Akagi, M.; Munakata, H.; Hamanishi, C. Reactive oxygen species depolymerize hyaluronan: Involvement of the hydroxyl radical. Pathophysiology 2003, 9, 215–220. [Google Scholar] [CrossRef]
- Avenoso, A.; Bruschetta, G.; D’Ascola, A.; Scuruchi, M.; Mandraffino, G.; Gullace, R.; Saitta, A.; Campo, S.; Campo, G.M. Hyaluronan fragments produced during tissue injury: A signal amplifying the inflammatory response. Arch. Biochem. Biophys. 2019, 663, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Albano, G.D.; Bonanno, A.; Cavalieri, L.; Ingrassia, E.; Di Sano, C.; Siena, L.; Riccobono, L.; Gagliardo, R.; Profita, M. Effect of High, Medium, and Low Molecular Weight Hyaluronan on Inflammation and Oxidative Stress in an In Vitro Model of Human Nasal Epithelial Cells. Mediat. Inflamm 2016, 2016, 8727289. [Google Scholar] [CrossRef] [Green Version]
- Yung, S.; Chan, T.M. Hyaluronan--regulator and initiator of peritoneal inflammation and remodeling. Int. J. Artif. Organs 2007, 30, 477–483. [Google Scholar] [CrossRef]
- Dong, Y.; Arif, A.; Olsson, M.; Cali, V.; Hardman, B.; Dosanjh, M.; Lauer, M.; Midura, R.J.; Hascall, V.C.; Brown, K.L.; et al. Endotoxin free hyaluronan and hyaluronan fragments do not stimulate TNF-alpha, interleukin-12 or upregulate co-stimulatory molecules in dendritic cells or macrophages. Sci. Rep. 2016, 6, 36928. [Google Scholar] [CrossRef] [PubMed]
- Safrankova, B.; Hermannova, M.; Nesporova, K.; Velebny, V.; Kubala, L. Absence of differences among low, middle, and high molecular weight hyaluronan in activating murine immune cells in vitro. Int. J. Biol. Macromol. 2018, 107, 1–8. [Google Scholar] [CrossRef]
- Scott, J.E.; Cummings, C.; Brass, A.; Chen, Y. Secondary and tertiary structures of hyaluronan in aqueous solution, investigated by rotary shadowing-electron microscopy and computer simulation. Hyaluronan is a very efficient network-forming polymer. Biochem. J. 1991, 274 Pt 3, 699–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, E.N.; Baker, E. Role of the Hyaluronan Receptor, Stabilin-2/HARE, in Health and Disease. Int. J. Mol. Sci. 2020, 21, 3504. [Google Scholar] [CrossRef] [PubMed]
- de la Motte, C.A. Hyaluronan in intestinal homeostasis and inflammation: Implications for fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G945–G949. [Google Scholar] [CrossRef]
- Wilson, R.B. Hypoxia, cytokines and stromal recruitment: Parallels between pathophysiology of encapsulating peritoneal sclerosis, endometriosis and peritoneal metastasis. Pleura Peritoneum 2018, 3, 20180103. [Google Scholar] [CrossRef]
- Banerji, S.; Wright, A.J.; Noble, M.; Mahoney, D.J.; Campbell, I.D.; Day, A.J.; Jackson, D.G. Structures of the Cd44-hyaluronan complex provide insight into a fundamental carbohydrate-protein interaction. Nat. Struct. Mol. Biol. 2007, 14, 234–239. [Google Scholar] [CrossRef]
- Wolny, P.M.; Banerji, S.; Gounou, C.; Brisson, A.R.; Day, A.J.; Jackson, D.G.; Richter, R.P. Analysis of CD44-hyaluronan interactions in an artificial membrane system: Insights into the distinct binding properties of high and low molecular weight hyaluronan. J. Biol. Chem. 2010, 285, 30170–30180. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Cao, M.; Liu, H.; He, Y.; Xu, J.; Du, Y.; Liu, Y.; Wang, W.; Cui, L.; Hu, J.; et al. The high and low molecular weight forms of hyaluronan have distinct effects on CD44 clustering. J. Biol. Chem. 2012, 287, 43094–43107. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Owen, S.C. Hyaluronic acid binding to CD44S is indiscriminate of molecular weight. Biochim. Biophys. Acta-Biomembr. 2020, 1862, 183348. [Google Scholar] [CrossRef]
- Sunabori, T.; Koike, M.; Asari, A.; Oonuki, Y.; Uchiyama, Y. Suppression of Ischemia-Induced Hippocampal Pyramidal Neuron Death by Hyaluronan Tetrasaccharide through Inhibition of Toll-Like Receptor 2 Signaling Pathway. Am. J. Pathol. 2016, 186, 2143–2151. [Google Scholar] [CrossRef] [Green Version]
- Rugg, M.S.; Willis, A.C.; Mukhopadhyay, D.; Hascall, V.C.; Fries, E.; Fulop, C.; Milner, C.M.; Day, A.J. Characterization of complexes formed between TSG-6 and inter-alpha-inhibitor that act as intermediates in the covalent transfer of heavy chains onto hyaluronan. J. Biol. Chem. 2005, 280, 25674–25686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesley, J.; Gal, I.; Mahoney, D.J.; Cordell, M.R.; Rugg, M.S.; Hyman, R.; Day, A.J.; Mikecz, K. TSG-6 modulates the interaction between hyaluronan and cell surface CD44. J. Biol. Chem. 2004, 279, 25745–25754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulop, C.; Szanto, S.; Mukhopadhyay, D.; Bardos, T.; Kamath, R.V.; Rugg, M.S.; Day, A.J.; Salustri, A.; Hascall, V.C.; Glant, T.T.; et al. Impaired cumulus mucification and female sterility in tumor necrosis factor-induced protein-6 deficient mice. Development 2003, 130, 2253–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasanello, D.C.; Su, J.; Deng, S.; Yin, R.; Colville, M.J.; Berenson, J.M.; Kelly, C.M.; Freer, H.; Rollins, A.; Wagner, B.; et al. Hyaluronic acid synthesis, degradation, and crosslinking in equine osteoarthritis: TNF-alpha-TSG-6-mediated HC-HA formation. Arthritis Res. 2021, 23, 218. [Google Scholar] [CrossRef]
- Ni, K.; Gill, A.; Tseng, V.; Mikosz, A.M.; Koike, K.; Beatman, E.L.; Xu, C.Y.; Cao, D.; Gally, F.; Mould, K.J.; et al. Rapid clearance of heavy chain-modified hyaluronan during resolving acute lung injury. Respir. Res. 2018, 19, 107. [Google Scholar] [CrossRef]
- Bogdani, M.; Johnson, P.Y.; Potter-Perigo, S.; Nagy, N.; Day, A.J.; Bollyky, P.L.; Wight, T.N. Hyaluronan and hyaluronan-binding proteins accumulate in both human type 1 diabetic islets and lymphoid tissues and associate with inflammatory cells in insulitis. Diabetes 2014, 63, 2727–2743. [Google Scholar] [CrossRef] [Green Version]
- Mutoji, K.N.; Sun, M.; Nash, A.; Puri, S.; Hascall, V.; Coulson-Thomas, V.J. Anti-inflammatory protein TNFalpha-stimulated gene-6 (TSG-6) reduces inflammatory response after brain injury in mice. BMC Immunol. 2021, 22, 52. [Google Scholar] [CrossRef]
- Kang, I.; Chang, M.Y.; Wight, T.N.; Frevert, C.W. Proteoglycans as Immunomodulators of the Innate Immune Response to Lung Infection. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2018, 66, 241–259. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.; Wight, T.N.; Frevert, C.W. Proteoglycans: Key regulators of pulmonary inflammation and the innate immune response to lung infection. Anat. Rec. 2010, 293, 968–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, M.; Diao, J.; Gu, H.; Khatri, I.; Zhao, J.; Cattral, M.S. Toll-like Receptor 2 Activation Promotes Tumor Dendritic Cell Dysfunction by Regulating IL-6 and IL-10 Receptor Signaling. Cell Rep. 2015, 13, 2851–2864. [Google Scholar] [CrossRef] [Green Version]
- Albeiroti, S.; Soroosh, A.; de la Motte, C.A. Hyaluronan’s Role in Fibrosis: A Pathogenic Factor or a Passive Player? Biomed. Res. Int. 2015, 2015, 790203. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, W.J.; Mullen, P.J.; Schmid, E.W.; Flores, A.; Momcilovic, M.; Sharpley, M.S.; Jelinek, D.; Whiteley, A.E.; Maxwell, M.B.; Wilde, B.R.; et al. Extracellular Matrix Remodeling Regulates Glucose Metabolism through TXNIP Destabilization. Cell 2018, 175, 117–132.e121. [Google Scholar] [CrossRef] [Green Version]
- Si, M.; Wang, Q.; Li, Y.; Lin, H.; Luo, D.; Zhao, W.; Dou, X.; Liu, J.; Zhang, H.; Huang, Y.; et al. Inhibition of hyperglycolysis in mesothelial cells prevents peritoneal fibrosis. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef]
- Zhao, X.; Psarianos, P.; Ghoraie, L.S.; Yip, K.; Goldstein, D.; Gilbert, R.; Witterick, I.; Pang, H.; Hussain, A.; Lee, J.H.; et al. Metabolic regulation of dermal fibroblasts contributes to skin extracellular matrix homeostasis and fibrosis. Nat. Metab. 2019, 1, 147–157. [Google Scholar] [CrossRef]
- Chanmee, T.; Ontong, P.; Izumikawa, T.; Higashide, M.; Mochizuki, N.; Chokchaitaweesuk, C.; Khansai, M.; Nakajima, K.; Kakizaki, I.; Kongtawelert, P.; et al. Hyaluronan Production Regulates Metabolic and Cancer Stem-like Properties of Breast Cancer Cells via Hexosamine Biosynthetic Pathway-coupled HIF-1 Signaling. J. Biol. Chem. 2016, 291, 24105–24120. [Google Scholar] [CrossRef] [Green Version]
- Grandoch, M.; Flogel, U.; Virtue, S.; Maier, J.K.; Jelenik, T.; Kohlmorgen, C.; Feldmann, K.; Ostendorf, Y.; Castaneda, T.R.; Zhou, Z.; et al. 4-Methylumbelliferone improves the thermogenic capacity of brown adipose tissue. Nat. Metab. 2019, 1, 546–559. [Google Scholar] [CrossRef]
- Tammi, M.I.; Oikari, S.; Pasonen-Seppanen, S.; Rilla, K.; Auvinen, P.; Tammi, R.H. Activated hyaluronan metabolism in the tumor matrix—Causes and consequences. Matrix Biol. J. Int. Soc. Matrix Biol. 2019, 78-79, 147–164. [Google Scholar] [CrossRef]
- Tsitrina, A.A.; Krasylov, I.V.; Maltsev, D.I.; Andreichenko, I.N.; Moskvina, V.S.; Ivankov, D.N.; Bulgakova, E.V.; Nesterchuk, M.; Shashkovskaya, V.; Dashenkova, N.O.; et al. Inhibition of hyaluronan secretion by novel coumarin compounds and chitin synthesis inhibitors. Glycobiology 2021, 31, 959–974. [Google Scholar] [CrossRef]
- Tsuchiya, S.; Ohashi, Y.; Ishizuka, S.; Ishiguro, N.; O’Rourke, D.P.; Knudson, C.B.; Knudson, W. Suppression of murine osteoarthritis by 4-methylumbelliferone. J. Orthop. Res. 2020, 38, 1122–1131. [Google Scholar] [CrossRef]
- Terabe, K.; Ohashi, Y.; Tsuchiya, S.; Ishizuka, S.; Knudson, C.B.; Knudson, W. Chondroprotective effects of 4-methylumbelliferone and hyaluronan synthase-2 overexpression involve changes in chondrocyte energy metabolism. J. Biol. Chem. 2019, 294, 17799–17817. [Google Scholar] [CrossRef] [PubMed]
- Andreichenko, I.N.; Tsitrina, A.A.; Fokin, A.V.; Gabdulkhakova, A.I.; Maltsev, D.I.; Perelman, G.S.; Bulgakova, E.V.; Kulikov, A.M.; Mikaelyan, A.S.; Kotelevtsev, Y.V. 4-methylumbelliferone Prevents Liver Fibrosis by Affecting Hyaluronan Deposition, FSTL1 Expression and Cell Localization. Int. J. Mol. Sci. 2019, 20, 6301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, J.M.; Sinha, R.; Seita, J.; Fernhoff, N.; Christ, S.; Koopmans, T.; Krampitz, G.W.; McKenna, K.M.; Xing, L.; Sandholzer, M.; et al. Surgical adhesions in mice are derived from mesothelial cells and can be targeted by antibodies against mesothelial markers. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koopmans, T.; Rinkevich, Y. Mesothelial to mesenchyme transition as a major developmental and pathological player in trunk organs and their cavities. Commun. Biol. 2018, 1, 170. [Google Scholar] [CrossRef] [Green Version]
- Mutsaers, S.E.; Prele, C.M.; Pengelly, S.; Herrick, S.E. Mesothelial cells and peritoneal homeostasis. Fertil. Steril. 2016, 106, 1018–1024. [Google Scholar] [CrossRef] [Green Version]
- Witowski, J.; Jorres, A. Peritoneal cell culture: Fibroblasts. Perit. Dial. Int. 2006, 26, 292–299. [Google Scholar] [CrossRef]
- Kawanishi, K. Diverse properties of the mesothelial cells in health and disease. Pleura Peritoneum 2016, 1, 79–89. [Google Scholar] [CrossRef]
- Fortin, C.N.; Saed, G.M.; Diamond, M.P. Predisposing factors to post-operative adhesion development. Hum. Reprod. Update 2015, 21, 536–551. [Google Scholar] [CrossRef] [Green Version]
- Yung, S.; Coles, G.A.; Williams, J.D.; Davies, M. The source and possible significance of hyaluronan in the peritoneal cavity. Kidney Int. 1994, 46, 527–533. [Google Scholar] [CrossRef] [Green Version]
- Yung, S.; Thomas, G.J.; Davies, M. Induction of hyaluronan metabolism after mechanical injury of human peritoneal mesothelial cells in vitro. Kidney Int. 2000, 58, 1953–1962. [Google Scholar] [CrossRef]
- Tzuman, Y.C.; Sapoznik, S.; Granot, D.; Nevo, N.; Neeman, M. Peritoneal adhesion and angiogenesis in ovarian carcinoma are inversely regulated by hyaluronan: The role of gonadotropins. Neoplasia 2010, 12, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koistinen, V.; Jokela, T.; Oikari, S.; Karna, R.; Tammi, M.; Rilla, K. Hyaluronan-positive plasma membrane protrusions exist on mesothelial cells in vivo. Histochem. Cell Biol. 2016, 145, 531–544. [Google Scholar] [CrossRef]
- Haslinger, B.; Mandl-Weber, S.; Sellmayer, A.; Sitter, T. Hyaluronan fragments induce the synthesis of MCP-1 and IL-8 in cultured human peritoneal mesothelial cells. Cell Tissue Res. 2001, 305, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Yung, S.; Coles, G.A.; Davies, M. IL-1 beta, a major stimulator of hyaluronan synthesis in vitro of human peritoneal mesothelial cells: Relevance to peritonitis in CAPD. Kidney Int. 1996, 50, 1337–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raby, A.C.; Gonzalez-Mateo, G.T.; Williams, A.; Topley, N.; Fraser, D.; Lopez-Cabrera, M.; Labeta, M.O. Targeting Toll-like receptors with soluble Toll-like receptor 2 prevents peritoneal dialysis solution-induced fibrosis. Kidney Int. 2018, 94, 346–362. [Google Scholar] [CrossRef]
- Shi, J.; Li, Q.; Sheng, M.; Zheng, M.; Yu, M.; Zhang, L. The Role of TLR4 in M1 Macrophage-Induced Epithelial-Mesenchymal Transition of Peritoneal Mesothelial Cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2016, 40, 1538–1548. [Google Scholar] [CrossRef]
- Tsai, J.M.; Shoham, M.; Fernhoff, N.B.; George, B.M.; Marjon, K.D.; McCracken, M.N.; Kao, K.S.; Sinha, R.; Volkmer, A.K.; Miyanishi, M.; et al. Neutrophil and monocyte kinetics play critical roles in mouse peritoneal adhesion formation. Blood Adv. 2019, 3, 2713–2721. [Google Scholar] [CrossRef] [Green Version]
- Dechaud, H.; Witz, C.A.; Montoya-Rodriguez, I.A.; Degraffenreid, L.A.; Schenken, R.S. Mesothelial cell-associated hyaluronic acid promotes adhesion of endometrial cells to mesothelium. Fertil. Steril. 2001, 76, 1012–1018. [Google Scholar] [CrossRef]
- Rodgers, A.K.; Nair, A.; Binkley, P.A.; Tekmal, R.; Schenken, R.S. Inhibition of CD44 N- and O-linked glycosylation decreases endometrial cell lines attachment to peritoneal mesothelial cells. Fertil. Steril. 2011, 95, 823–825. [Google Scholar] [CrossRef] [Green Version]
- Krediet, R.T.; Struijk, D.G. Peritoneal changes in patients on long-term peritoneal dialysis. Nat. Rev. Nephrol. 2013, 9, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Li, Q.; Zhang, L.; Lin, H.; Hu, J.; Li, D.; Shi, S.; Cui, S.; Zhou, J.; Ji, J.; et al. Mesenchymal stem cells attenuate peritoneal injury through secretion of TSG-6. PLoS ONE 2012, 7, e43768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanda, R.; Hamada, C.; Kaneko, K.; Nakano, T.; Wakabayashi, K.; Io, H.; Horikoshi, S.; Tomino, Y. Pentraxin 3 as a new biomarker of peritoneal injury in peritoneal dialysis patients. J. Artif. Organs 2013, 16, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.S.; Jou, I.M.; Jung, Y.C.; Wu, C.L.; Shiau, A.L.; Chen, C.Y. In vivo expression of thrombospondin-1 suppresses the formation of peritoneal adhesion in rats. World J. Gastrointest Surg. 2019, 11, 85–92. [Google Scholar] [CrossRef]
- Osada, S.; Hamada, C.; Shimaoka, T.; Kaneko, K.; Horikoshi, S.; Tomino, Y. Alterations in proteoglycan components and histopathology of the peritoneum in uraemic and peritoneal dialysis (PD) patients. Nephrol. Dial. Transpl. 2009, 24, 3504–3512. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Balaji, S.; Steen, E.H.; Blum, A.J.; Li, H.; Chan, C.K.; Manson, S.R.; Lu, T.C.; Rae, M.M.; Austin, P.F.; et al. High-molecular weight hyaluronan attenuates tubulointerstitial scarring in kidney injury. JCI Insight 2020, 5, e136345. [Google Scholar] [CrossRef]
- Chang, S.H.; Yeh, Y.H.; Lee, J.L.; Hsu, Y.J.; Kuo, C.T.; Chen, W.J. Transforming growth factor-beta-mediated CD44/STAT3 signaling contributes to the development of atrial fibrosis and fibrillation. Basic Res. Cardiol. 2017, 112, 58. [Google Scholar] [CrossRef]
- Decleves, A.E.; Caron, N.; Voisin, V.; Legrand, A.; Bouby, N.; Kultti, A.; Tammi, M.I.; Flamion, B. Synthesis and fragmentation of hyaluronan in renal ischaemia. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc.-Eur. Ren. Assoc. 2012, 27, 3771–3781. [Google Scholar] [CrossRef] [Green Version]
- Frangogiannis, N.G. Cardiac fibrosis. Cardiovasc. Res. 2021, 117, 1450–1488. [Google Scholar] [CrossRef]
- Muller, J.; Gorressen, S.; Grandoch, M.; Feldmann, K.; Kretschmer, I.; Lehr, S.; Ding, Z.; Schmitt, J.P.; Schrader, J.; Garbers, C.; et al. Interleukin-6-dependent phenotypic modulation of cardiac fibroblasts after acute myocardial infarction. Basic Res. Cardiol. 2014, 109, 440. [Google Scholar] [CrossRef]
- Colombaro, V.; Decleves, A.E.; Jadot, I.; Voisin, V.; Giordano, L.; Habsch, I.; Nonclercq, D.; Flamion, B.; Caron, N. Inhibition of hyaluronan is protective against renal ischaemia-reperfusion injury. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc.-Eur. Ren. Assoc. 2013, 28, 2484–2493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meran, S.; Thomas, D.; Stephens, P.; Martin, J.; Bowen, T.; Phillips, A.; Steadman, R. Involvement of hyaluronan in regulation of fibroblast phenotype. J. Biol. Chem. 2007, 282, 25687–25697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meran, S.; Thomas, D.W.; Stephens, P.; Enoch, S.; Martin, J.; Steadman, R.; Phillips, A.O. Hyaluronan facilitates transforming growth factor-beta1-mediated fibroblast proliferation. J. Biol. Chem. 2008, 283, 6530–6545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, R.H.; Thomas, G.J.; Williams, J.D.; Steadman, R. Myofibroblastic differentiation leads to hyaluronan accumulation through reduced hyaluronan turnover. J. Biol. Chem. 2004, 279, 41453–41460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ontong, P.; Prachayasittikul, V. Unraveled roles of hyaluronan in severe COVID-19. EXCLI J. 2021, 20, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Kulshrestha, R.; Bansal, S.K. Dynamic role of LMW-hyaluronan fragments and Toll-like receptors 2,4 in progression of bleomycin induced lung parenchymal injury to fibrosis. Egypt. J. Bronchol. 2021, 15, 1–13. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, D.; Liang, J.; Meltzer, E.B.; Gray, A.; Miura, R.; Wogensen, L.; Yamaguchi, Y.; Noble, P.W. Severe lung fibrosis requires an invasive fibroblast phenotype regulated by hyaluronan and CD44. J. Exp. Med. 2011, 208, 1459–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huebener, P.; Abou-Khamis, T.; Zymek, P.; Bujak, M.; Ying, X.; Chatila, K.; Haudek, S.; Thakker, G.; Frangogiannis, N.G. CD44 is critically involved in infarct healing by regulating the inflammatory and fibrotic response. J. Immunol. 2008, 180, 2625–2633. [Google Scholar] [CrossRef]
- Midgley, A.C.; Oltean, S.; Hascall, V.; Woods, E.L.; Steadman, R.; Phillips, A.O.; Meran, S. Nuclear hyaluronidase 2 drives alternative splicing of CD44 pre-mRNA to determine profibrotic or antifibrotic cell phenotype. Sci. Signal. 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Patouraux, S.; Rousseau, D.; Bonnafous, S.; Lebeaupin, C.; Luci, C.; Canivet, C.M.; Schneck, A.S.; Bertola, A.; Saint-Paul, M.C.; Iannelli, A.; et al. CD44 is a key player in non-alcoholic steatohepatitis. J. Hepatol. 2017, 67, 328–338. [Google Scholar] [CrossRef]
- Yang, Y.M.; Noureddin, M.; Liu, C.; Ohashi, K.; Kim, S.Y.; Ramnath, D.; Powell, E.E.; Sweet, M.J.; Roh, Y.S.; Hsin, I.F.; et al. Hyaluronan synthase 2-mediated hyaluronan production mediates Notch1 activation and liver fibrosis. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Zaman, A.; Cui, Z.; Foley, J.P.; Zhao, H.; Grimm, P.C.; Delisser, H.M.; Savani, R.C. Expression and role of the hyaluronan receptor RHAMM in inflammation after bleomycin injury. Am. J. Respir. Cell Mol. Biol. 2005, 33, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Cui, Z.; Liao, J.; Cheong, N.; Longoria, C.; Cao, G.; DeLisser, H.M.; Savani, R.C. The Receptor for Hyaluronan-Mediated Motility (CD168) promotes inflammation and fibrosis after acute lung injury. Matrix Biol. J. Int. Soc. Matrix Biol. 2019, 78-79, 255–271. [Google Scholar] [CrossRef] [PubMed]
- Colombaro, V.; Jadot, I.; Decleves, A.E.; Voisin, V.; Giordano, L.; Habsch, I.; Malaisse, J.; Flamion, B.; Caron, N. Lack of hyaluronidases exacerbates renal post-ischemic injury, inflammation, and fibrosis. Kidney Int. 2015, 88, 61–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evanko, S.P.; Potter-Perigo, S.; Petty, L.J.; Workman, G.A.; Wight, T.N. Hyaluronan Controls the Deposition of Fibronectin and Collagen and Modulates TGF-beta1 Induction of Lung Myofibroblasts. Matrix Biol. 2015, 42, 74–92. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Sue, Y.M.; Cheng, C.Y.; Chen, Y.C.; Liu, C.T.; Hsu, Y.H.; Hwang, P.A.; Huang, N.J.; Chen, T.H. Oligo-fucoidan prevents renal tubulointerstitial fibrosis by inhibiting the CD44 signal pathway. Sci. Rep. 2017, 7, 40183. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocurkova, A.; Nesporova, K.; Sandanusova, M.; Kerberova, M.; Lehka, K.; Velebny, V.; Kubala, L.; Ambrozova, G. Endogenously-Produced Hyaluronan and Its Potential to Regulate the Development of Peritoneal Adhesions. Biomolecules 2022, 12, 45. https://doi.org/10.3390/biom12010045
Kocurkova A, Nesporova K, Sandanusova M, Kerberova M, Lehka K, Velebny V, Kubala L, Ambrozova G. Endogenously-Produced Hyaluronan and Its Potential to Regulate the Development of Peritoneal Adhesions. Biomolecules. 2022; 12(1):45. https://doi.org/10.3390/biom12010045
Chicago/Turabian StyleKocurkova, Anna, Kristina Nesporova, Miriam Sandanusova, Michaela Kerberova, Katerina Lehka, Vladimir Velebny, Lukas Kubala, and Gabriela Ambrozova. 2022. "Endogenously-Produced Hyaluronan and Its Potential to Regulate the Development of Peritoneal Adhesions" Biomolecules 12, no. 1: 45. https://doi.org/10.3390/biom12010045
APA StyleKocurkova, A., Nesporova, K., Sandanusova, M., Kerberova, M., Lehka, K., Velebny, V., Kubala, L., & Ambrozova, G. (2022). Endogenously-Produced Hyaluronan and Its Potential to Regulate the Development of Peritoneal Adhesions. Biomolecules, 12(1), 45. https://doi.org/10.3390/biom12010045