
Modulation of Canine Melanocortin-3 and -4 Receptors by Melanocortin-2 Receptor Accessory Protein 1 and 2



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ligands and Plasmids
2.2. Cell Culture and Transfection
2.3. Ligand Binding Assays
2.4. Ligand-Stimulated cAMP Assays
2.5. Flow Cytometry Assay
2.6. Statistical Analysis
3. Results
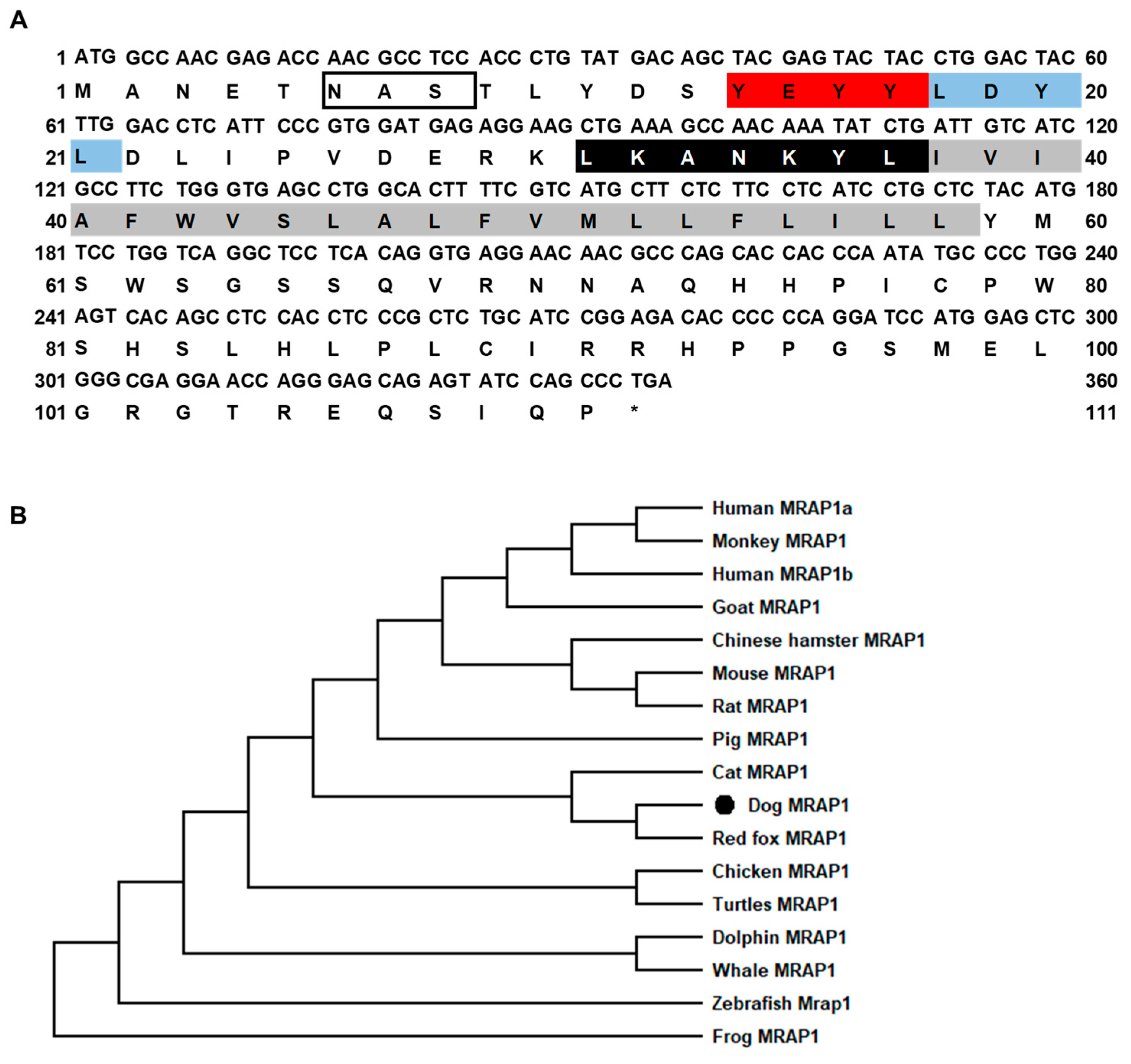
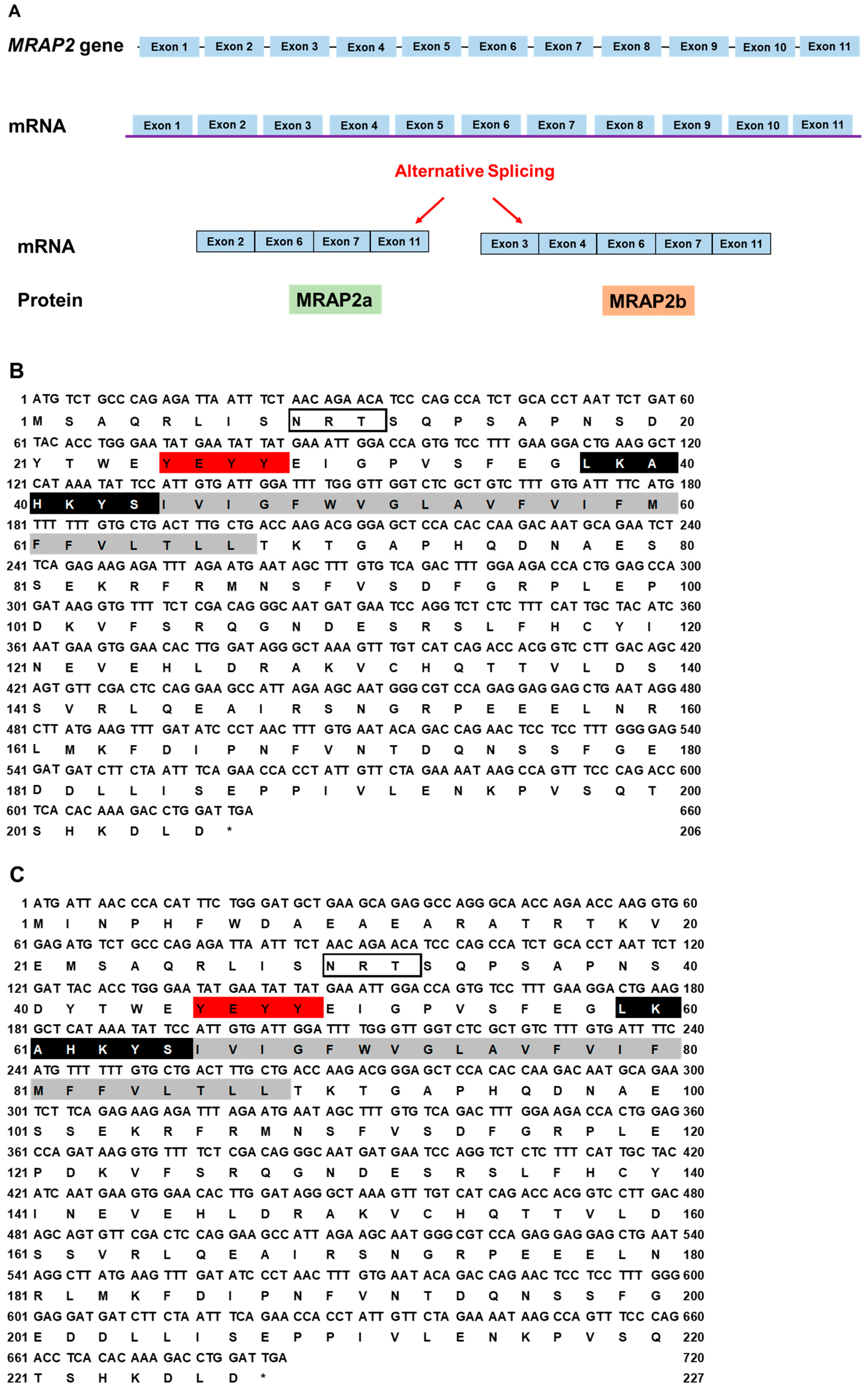
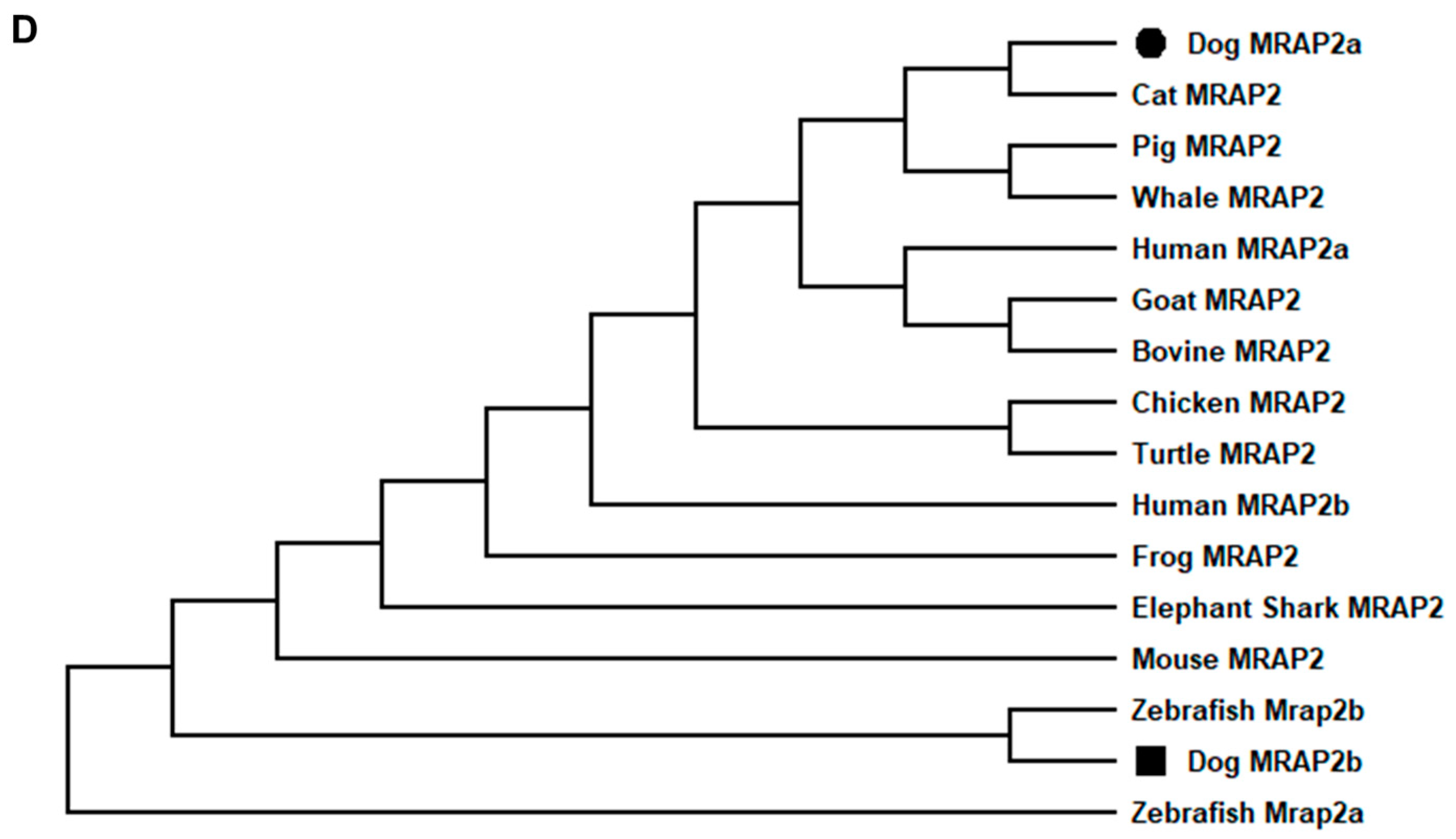
3.1. Nucleotide and Deduced Amino Acid Sequences of cMC3R, cMRAP1, and cMRAP2s
3.2. Ligand Binding Properties of cMC3R
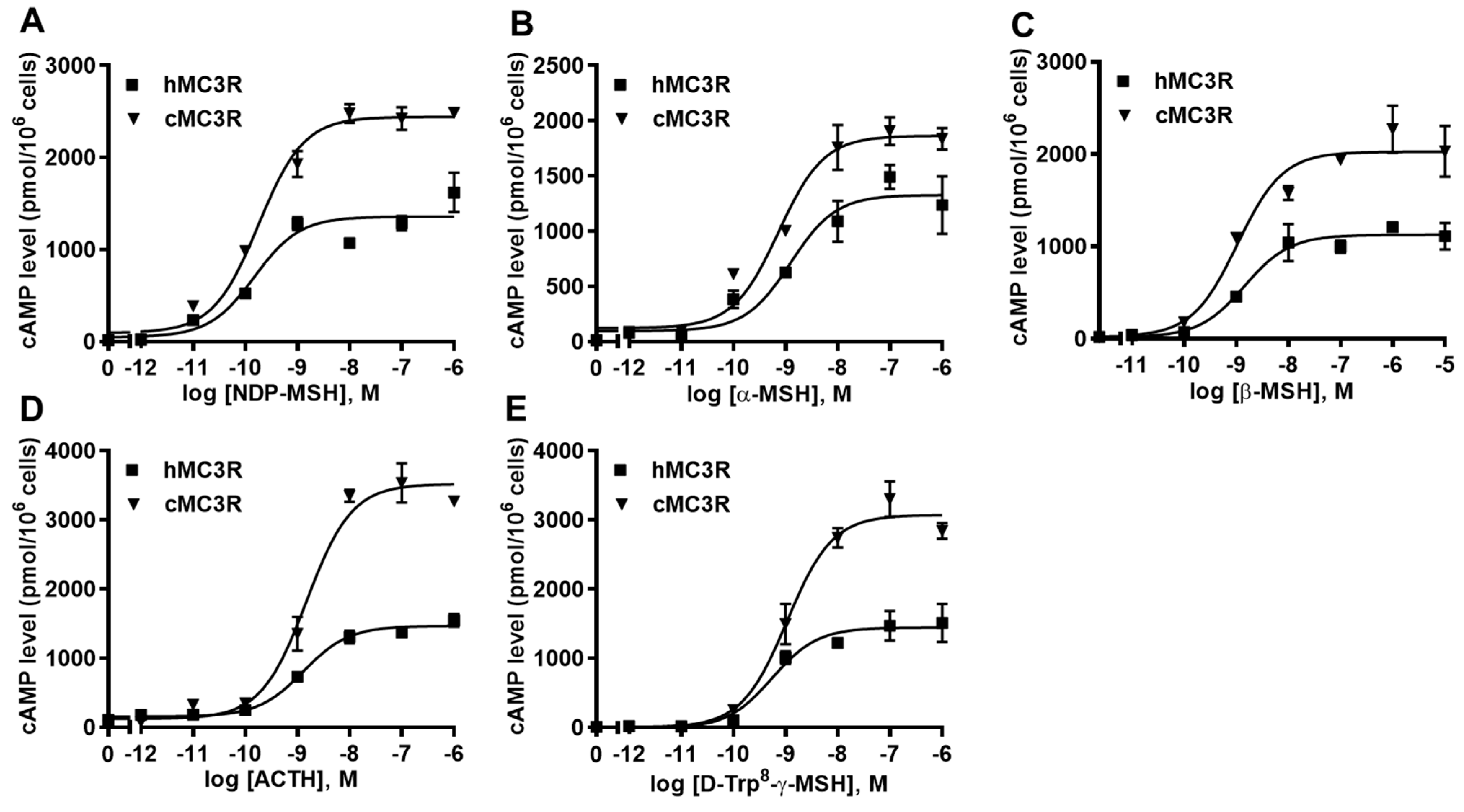
3.3. cAMP Signaling Properties of cMC3R
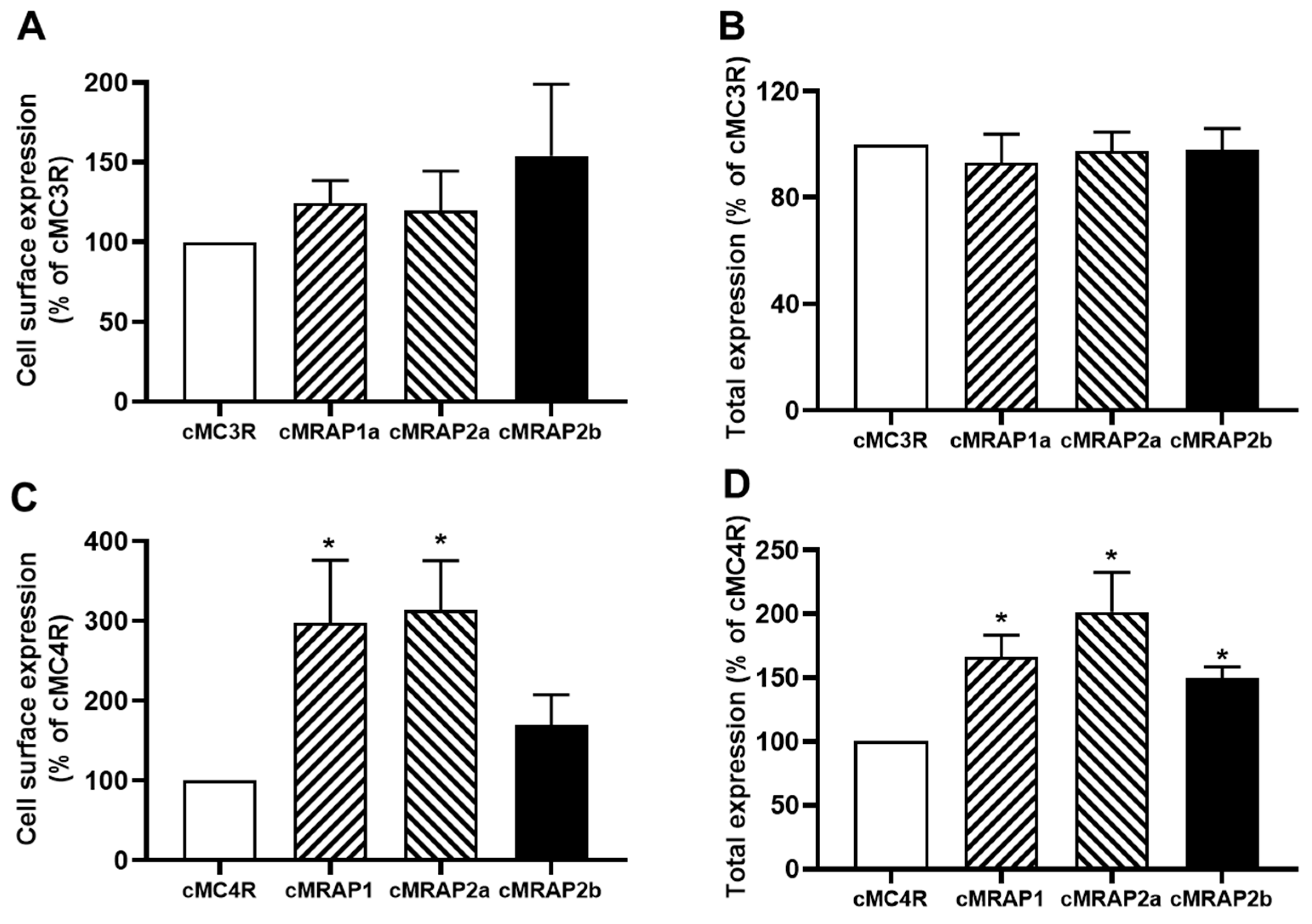
3.4. Modulation of cMC3R and cMC4R Expression by MRAPs
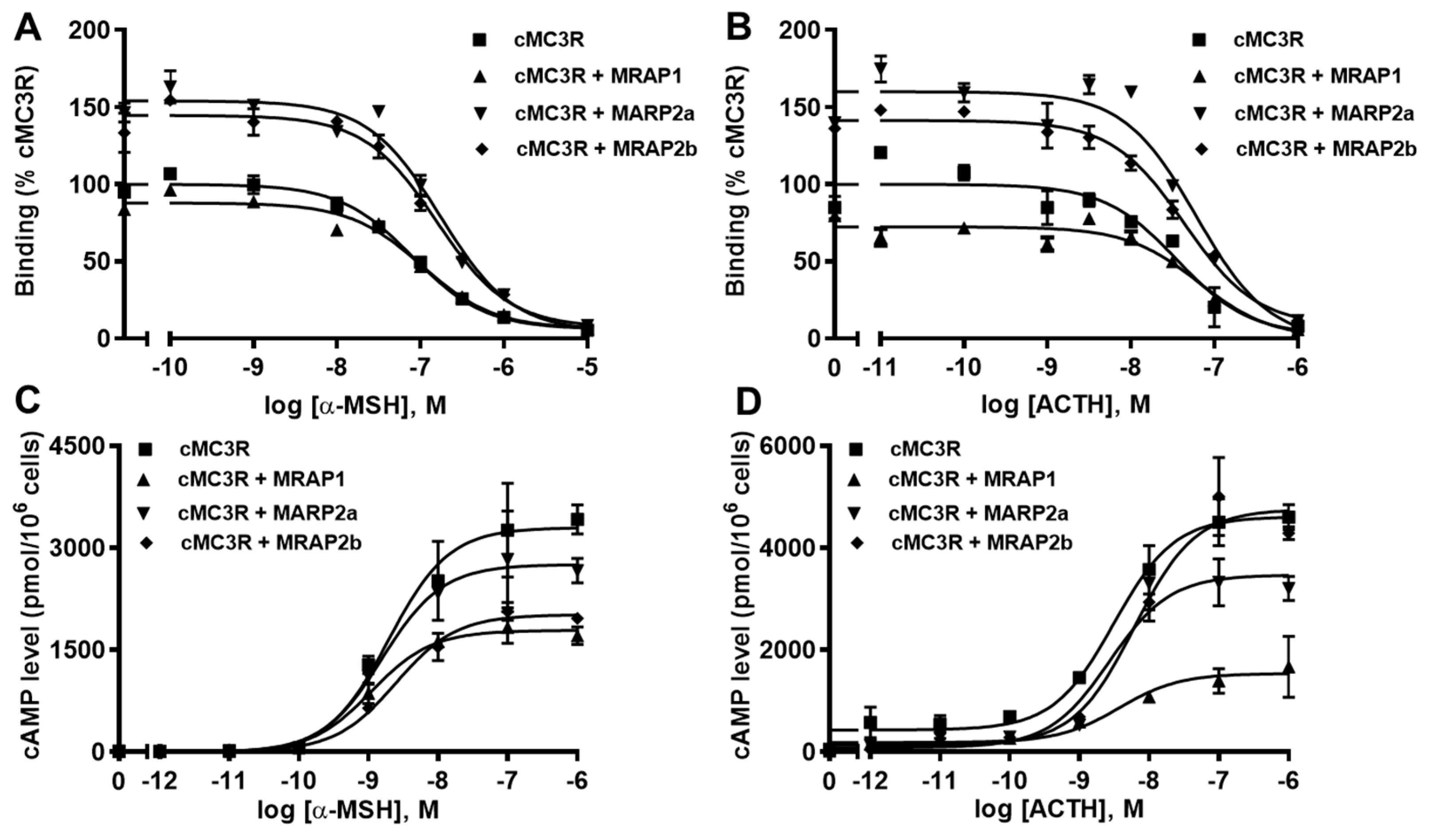
3.5. Modulation of cMC3R Pharmacology by MRAPs
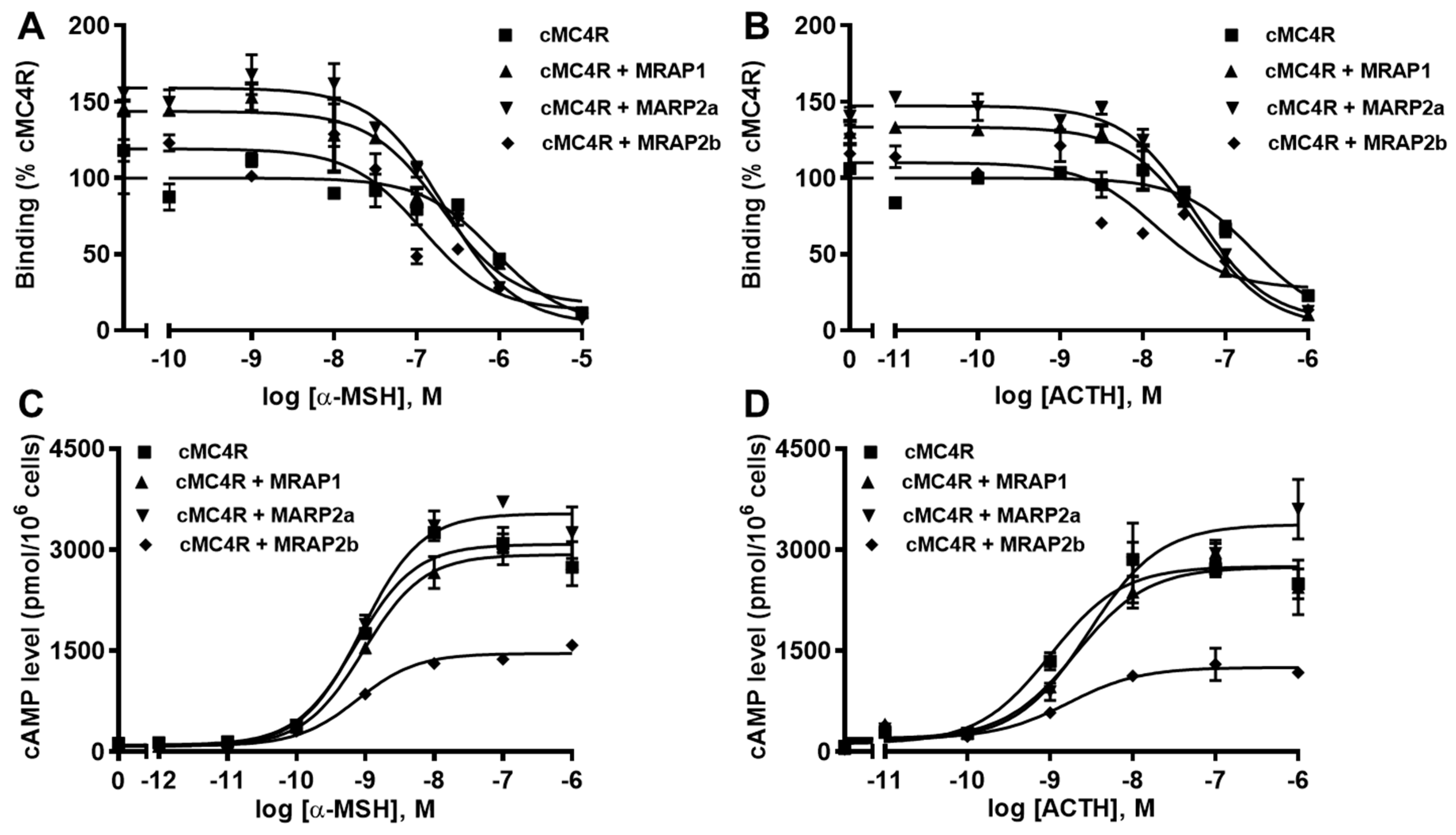
3.6. Modulation of cMC4R Pharmacology by MRAPs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gantz, I.; Miwa, H.; Konda, Y.; Shimoto, Y.; Tashiro, T.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular cloning, expression, and gene localization of a fourth melanocortin receptor. J. Biol. Chem. 1993, 268, 15174–15179. [Google Scholar] [CrossRef]
- Gantz, I.; Konda, Y.; Tashiro, T.; Shimoto, Y.; Miwa, H.; Munzert, G.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular cloning of a novel melanocortin receptor. J. Biol. Chem. 1993, 268, 8246–8250. [Google Scholar] [CrossRef]
- Roselli-Rehfuss, L.; Mountjoy, K.G.; Robbins, L.S.; Mortrud, M.T.; Low, M.J.; Tatro, J.B.; Entwistle, M.L.; Simerly, R.B.; Cone, R.D. Identification of a receptor for γ melanotropin and other proopiomelanocortin peptides in the hypothalamus and limbic system. Proc. Natl. Acad. Sci. USA 1993, 90, 8856–8860. [Google Scholar] [CrossRef] [Green Version]
- Mountjoy, K.G.; Mortrud, M.T.; Low, M.J.; Simerly, R.B.; Cone, R.D. Localization of the melanocortin-4 receptor (MC4-R) in neuroendocrine and autonomic control circuits in the brain. Mol. Endocrinol. 1994, 8, 1298–1308. [Google Scholar]
- Cone, R.D. Anatomy and regulation of the central melanocortin system. Nat. Neurosci. 2005, 8, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.X. Molecular mechanisms of the neural melanocortin receptor dysfunction in severe early onset obesity. Mol. Cell. Endocrinol. 2005, 239, 1–14. [Google Scholar] [CrossRef]
- Liu, T.; Ji, R.L.; Tao, Y.X. Naturally occurring mutations in G protein-coupled receptors associated with obesity and type 2 diabetes mellitus. Pharmacol. Ther. 2022, 234, 108044. [Google Scholar] [CrossRef]
- Chen, A.S.; Marsh, D.J.; Trumbauer, M.E.; Frazier, E.G.; Guan, X.M.; Yu, H.; Rosenblum, C.I.; Vongs, A.; Feng, Y.; Cao, L.; et al. Inactivation of the mouse melanocortin-3 receptor results in increased fat mass and reduced lean body mass. Nat. Genet. 2000, 26, 97–102. [Google Scholar] [CrossRef]
- Butler, A.A.; Kesterson, R.A.; Khong, K.; Cullen, M.J.; Pelleymounter, M.A.; Dekoning, J.; Baetscher, M.; Cone, R.D. A unique metabolic syndrome causes obesity in the melanocortin-3 receptor-deficient mouse. Endocrinology 2000, 141, 3518–3521. [Google Scholar] [CrossRef]
- Zhang, Y.; Kilroy, G.E.; Henagan, T.M.; Prpic-Uhing, V.; Richards, W.G.; Bannon, A.W.; Mynatt, R.L.; Gettys, T.W. Targeted deletion of melanocortin receptor subtypes 3 and 4, but not CART, alters nutrient partitioning and compromises behavioral and metabolic responses to leptin. FASEB J. 2005, 19, 1482–1491. [Google Scholar] [CrossRef]
- Huszar, D.; Lynch, C.A.; Fairchild-Huntress, V.; Dunmore, J.H.; Fang, Q.; Berkemeier, L.R.; Gu, W.; Kesterson, R.A.; Boston, B.A.; Cone, R.D.; et al. Targeted disruption of the melanocortin-4 receptor results in obesity in mice. Cell 1997, 88, 131–141. [Google Scholar] [CrossRef]
- Balthasar, N.; Dalgaard, L.T.; Lee, C.E.; Yu, J.; Funahashi, H.; Williams, T.; Ferreira, M.; Tang, V.; McGovern, R.A.; Kenny, C.D.; et al. Divergence of melanocortin pathways in the control of food intake and energy expenditure. Cell 2005, 123, 493–505. [Google Scholar] [CrossRef]
- Tao, Y.X. Mutations in melanocortin-4 receptor and human obesity. Prog. Mol. Biol. Transl. Sci. 2009, 88, 173–204. [Google Scholar]
- Tao, Y.X. Mutations in the melanocortin-3 receptor (MC3R) gene: Impact on human obesity or adiposity. Curr. Opin. Investig. Drugs 2010, 11, 1092–1096. [Google Scholar] [PubMed]
- Tao, Y.X. The melanocortin-4 receptor: Physiology, pharmacology, and pathophysiology. Endocr. Rev. 2010, 31, 506–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Tao, Y.X. Mutations in melanocortin-3 receptor gene and human obesity. Prog. Mol. Biol. Transl. Sci. 2016, 140, 97–129. [Google Scholar] [CrossRef]
- Chandramohan, G.; Durham, N.; Sinha, S.; Norris, K.; Vaziri, N.D. Role of γ melanocyte-stimulating hormone-renal melanocortin 3 receptor system in blood pressure regulation in salt-resistant and salt-sensitive rats. Metabolism 2009, 58, 1424–1429. [Google Scholar] [CrossRef]
- Versteeg, D.H.; Van Bergen, P.; Adan, R.A.; De Wildt, D.J. Melanocortins and cardiovascular regulation. Eur. J. Pharmacol. 1998, 360, 1–14. [Google Scholar] [CrossRef]
- Mioni, C.; Giuliani, D.; Cainazzo, M.M.; Leone, S.; Bazzani, C.; Grieco, P.; Novellino, E.; Tomasi, A.; Bertolini, A.; Guarini, S. Further evidence that melanocortins prevent myocardial reperfusion injury by activating melanocortin MC3 receptors. Eur. J. Pharmacol. 2003, 477, 227–234. [Google Scholar] [CrossRef]
- Getting, S.J.; Christian, H.C.; Lam, C.W.; Gavins, F.N.; Flower, R.J.; Schioth, H.B.; Perretti, M. Redundancy of a functional melanocortin 1 receptor in the anti-inflammatory actions of melanocortin peptides: Studies in the recessive yellow (e/e) mouse suggest an important role for melanocortin 3 receptor. J. Immunol. 2003, 170, 3323–3330. [Google Scholar] [CrossRef] [Green Version]
- Catania, A.; Gatti, S.; Colombo, G.; Lipton, J.M. Targeting melanocortin receptors as a novel strategy to control inflammation. Pharmacol. Rev. 2004, 56, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Getting, S.J.; Riffo-Vasquez, Y.; Pitchford, S.; Kaneva, M.; Grieco, P.; Page, C.P.; Perretti, M.; Spina, D. A role for MC3R in modulating lung inflammation. Pulm. Pharmacol. Ther. 2008, 21, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.B.; Montero-Melendez, T.; Greco, K.V.; Perretti, M. Melanocortin receptors as novel effectors of macrophage responses in inflammation. Front. Immunol. 2011, 2, 41. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Guo, D.Y.; Lin, Y.J.; Tao, Y.X. Melanocortin regulation of inflammation. Front. Endocrinol. 2019, 10, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, B.Y.H.; Williamson, A.; Finer, S.; Day, F.R.; Tadross, J.A.; Goncalves Soares, A.; Wade, K.; Sweeney, P.; Bedenbaugh, M.N.; Porter, D.T.; et al. MC3R links nutritional state to childhood growth and the timing of puberty. Nature 2021, 599, 436–441. [Google Scholar] [CrossRef]
- Smith, A.I.; Funder, J.W. Proopiomelanocortin processing in the pituitary, central nervous system, and peripheral tissues. Endocr. Rev. 1988, 9, 159–179. [Google Scholar] [CrossRef]
- Dores, R.M.; Lecaude, S. Trends in the evolution of the proopiomelanocortin gene. Gen. Comp. Endocrinol. 2005, 142, 81–93. [Google Scholar] [CrossRef]
- Metherell, L.A.; Chapple, J.P.; Cooray, S.; David, A.; Becker, C.; Ruschendorf, F.; Naville, D.; Begeot, M.; Khoo, B.; Nurnberg, P.; et al. Mutations in MRAP, encoding a new interacting partner of the ACTH receptor, cause familial glucocorticoid deficiency type 2. Nat. Genet. 2005, 37, 166–170. [Google Scholar] [CrossRef]
- Sebag, J.A.; Hinkle, P.M. Melanocortin-2 receptor accessory protein MRAP forms antiparallel homodimers. Proc. Natl. Acad. Sci. USA 2007, 104, 20244–20249. [Google Scholar] [CrossRef] [Green Version]
- Chan, L.F.; Webb, T.R.; Chung, T.T.; Meimaridou, E.; Cooray, S.N.; Guasti, L.; Chapple, J.P.; Egertova, M.; Elphick, M.R.; Cheetham, M.E.; et al. MRAP and MRAP2 are bidirectional regulators of the melanocortin receptor family. Proc. Natl. Acad. Sci. USA 2009, 106, 6146–6151. [Google Scholar] [CrossRef] [Green Version]
- Sebag, J.A.; Zhang, C.; Hinkle, P.M.; Bradshaw, A.M.; Cone, R.D. Developmental control of the melanocortin-4 receptor by MRAP2 proteins in zebrafish. Science 2013, 341, 278–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouault, A.A.J.; Srinivasan, D.K.; Yin, T.C.; Lee, A.A.; Sebag, J.A. Melanocortin receptor accessory proteins (MRAPs): Functions in the melanocortin system and beyond. Biochim. Biophys. Acta 2017, 1864, 2322–2329. [Google Scholar] [CrossRef]
- Tao, Y.X. Molecular chaperones and G protein-coupled receptor maturation and pharmacology. Mol. Cell. Endocrinol. 2020, 511, 110862. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Rached, M.; Gallo-Payet, N. Differential regulation of the human adrenocorticotropin receptor [melanocortin-2 receptor (MC2R)] by human MC2R accessory protein isoforms alpha and beta in isogenic human embryonic kidney 293 cells. Mol. Endocrinol. 2007, 21, 1656–1669. [Google Scholar] [CrossRef] [Green Version]
- Novoselova, T.; King, P.; Guasti, L.; Metherell, L.A.; Clark, A.J.L.; Chan, L.F. ACTH signalling and adrenal development: Lessons from mouse models. Endocr. Connect. 2019, 8, R122–R130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinkle, P.M.; Serasinghe, M.N.; Jakabowski, A.; Sebag, J.A.; Wilson, K.R.; Haskell-Luevano, C. Use of chimeric melanocortin-2 and -4 receptors to identify regions responsible for ligand specificity and dependence on melanocortin 2 receptor accessory protein. Eur. J. Pharmacol. 2011, 660, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Kay, E.I.; Botha, R.; Montgomery, J.M.; Mountjoy, K.G. hMRAPa increases αMSH-induced hMC1R and hMC3R functional coupling and hMC4R constitutive activity. J. Mol. Endocrinol. 2013, 50, 203–215. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, X.; Zhou, Y.; Cui, L.; Li, J.; Wu, C.; Wan, Y.; Li, J.; Wang, Y. The interaction of MC3R and MC4R with MRAP2, ACTH, α-MSH and AgRP in chickens. J. Endocrinol. 2017, 234, 155–174. [Google Scholar] [CrossRef]
- Tai, X.; Xue, S.; Zhang, C.; Liu, Y.; Chen, J.; Han, Y.; Lin, G.; Zhang, C. Pharmacological evaluation of MRAP proteins on Xenopus neural melanocortin signaling. J. Cell. Physiol. 2021, 236, 6344–6361. [Google Scholar] [CrossRef]
- Kay, E.I.; Botha, R.; Montgomery, J.M.; Mountjoy, K.G. hMRAPa specifically alters hMC4R molecular mass and N-linked complex glycosylation in HEK293 cells. J. Mol. Endocrinol. 2013, 50, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Kay, E.I.; Botha, R.; Montgomery, J.M.; Mountjoy, K.G. hMRAPα, but not hMRAP2, enhances hMC4R constitutive activity in HEK293 cells and this is not dependent on hMRAPα induced changes in hMC4R complex N-linked glycosylation. PLoS ONE 2015, 10, e0140320. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xu, Y.; Zheng, J.; Kuang, Z.; Zhang, C.; Li, N.; Lin, G.; Zhang, C. Pharmacological modulation of dual melanocortin-4 receptor signaling by melanocortin receptor accessory proteins in the Xenopus laevis. J. Cell. Physiol. 2021, 236, 5980–5993. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.L.; Tao, Y.X. Regulation of melanocortin-3 and -4 receptors by isoforms of melanocortin-2 receptor accessory protein 1 and 2. Biomolecules 2022, 12, 244. [Google Scholar] [CrossRef]
- Asai, M.; Ramachandrappa, S.; Joachim, M.; Shen, Y.; Zhang, R.; Nuthalapati, N.; Ramanathan, V.; Strochlic, D.E.; Ferket, P.; Linhart, K.; et al. Loss of function of the melanocortin 2 receptor accessory protein 2 is associated with mammalian obesity. Science 2013, 341, 275–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novoselova, T.V.; Larder, R.; Rimmington, D.; Lelliott, C.; Wynn, E.H.; Gorrigan, R.J.; Tate, P.H.; Guasti, L.; O’Rahilly, S.; Clark, A.J.L. Loss of Mrap2 is associated with Sim1 deficiency and increased circulating cholesterol. J. Endocrinol. 2016, 230, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Geets, E.; Zegers, D.; Beckers, S.; Verrijken, A.; Massa, G.; Van Hoorenbeeck, K.; Verhulst, S.; Van Gaal, L.; Van Hul, W. Copy number variation (CNV) analysis and mutation analysis of the 6q14.1-6q16.3 genes SIM1 and MRAP2 in Prader Willi like patients. Mol. Genet. Metab. 2016, 117, 383–388. [Google Scholar] [CrossRef]
- Baron, M.; Maillet, J.; Huyvaert, M.; Dechaume, A.; Boutry, R.; Loiselle, H.; Durand, E.; Toussaint, B.; Vaillant, E.; Philippe, J.; et al. Loss-of-function mutations in MRAP2 are pathogenic in hyperphagic obesity with hyperglycemia and hypertension. Nat. Med. 2019, 25, 1733–1738. [Google Scholar] [CrossRef]
- da Fonseca, A.C.P.; Abreu, G.M.; Zembrzuski, V.M.; Campos Junior, M.; Carneiro, J.R.I.; Nogueira Neto, J.F.; Magno, F.; Rosado, E.L.; Bozza, P.T.; de Cabello, G.M.K.; et al. Study of LEP, MRAP2 and POMC genes as potential causes of severe obesity in Brazilian patients. Eat. Weight Disord. 2021, 26, 1399–1408. [Google Scholar] [CrossRef]
- Yang, L.K.; Zhang, Z.R.; Wen, H.S.; Tao, Y.X. Characterization of channel catfish (Ictalurus punctatus) melanocortin-3 receptor reveals a potential network in regulation of energy homeostasis. Gen. Comp. Endocrinol. 2019, 277, 90–103. [Google Scholar] [CrossRef]
- Zhang, J.; Li, J.; Wu, C.; Hu, Z.; An, L.; Wan, Y.; Fang, C.; Zhang, X.; Li, J.; Wang, Y. The Asp298Asn polymorphism of melanocortin-4 receptor (MC4R) in pigs: Evidence for its potential effects on MC4R constitutive activity and cell surface expression. Anim. Genet. 2020, 51, 694–706. [Google Scholar] [CrossRef]
- Tao, M.; Ji, R.L.; Huang, L.; Fan, S.Y.; Liu, T.; Liu, S.J.; Tao, Y.X. Regulation of melanocortin-4 receptor pharmacology by two isoforms of melanocortin receptor accessory protein 2 in topmouth culter (Culter alburnus). Front. Endocrinol. 2020, 11, 538. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.L.; Huang, L.; Wang, Y.; Liu, T.; Fan, S.Y.; Tao, M.; Tao, Y.X. Topmouth culter melanocortin-3 receptor: Regulation by two isoforms of melanocortin-2 receptor accessory protein 2. Endocr. Connect. 2021, 10, 1489–1501. [Google Scholar] [CrossRef]
- Wen, Z.Y.; Liu, T.; Qin, C.J.; Zou, Y.C.; Wang, J.; Li, R.; Tao, Y.X. MRAP2 interaction with melanocortin-4 receptor in snakehead (Channa argus). Biomolecules 2021, 11, 481. [Google Scholar] [CrossRef]
- Yan, J.; Tao, Y.X. Pharmacological characterization of canine melancortin-4 receptor and its natural variant V213F. Domest. Anim. Endocrinol. 2011, 41, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Steiner, A.L.; Kipnis, D.M.; Utiger, R.; Parker, C. Radioimmunoassay for the measurement of adenosine 3′,5′-cyclic phosphate. Proc. Natl. Acad. Sci. USA 1969, 64, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Mo, X.L.; Yang, R.; Tao, Y.X. Functions of transmembrane domain 3 of human melanocortin-4 receptor. J. Mol. Endocrinol. 2012, 49, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.X.; Segaloff, D.L. Functional characterization of melanocortin-4 receptor mutations associated with childhood obesity. Endocrinology 2003, 144, 4544–4551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Okayama, H. High-efficiency transformation of mammalian cells by plasmid DNA. Mol. Cell Biol. 1987, 7, 2745–2752. [Google Scholar] [PubMed] [Green Version]
- Wang, S.X.; Fan, Z.C.; Tao, Y.X. Functions of acidic transmembrane residues in human melanocortin-3 receptor binding and activation. Biochem Pharmacol 2008, 76, 520–530. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.Q.; Hou, Z.S.; Wen, H.S.; Li, Y.; Qi, X.; Li, W.J.; Tao, Y.X. Melanocortin-4 receptor in spotted sea bass, Lateolabrax maculatus: Cloning, tissue distribution, physiology, and pharmacology. Front. Endocrinol. 2019, 10, 705. [Google Scholar] [CrossRef] [Green Version]
- Gruber, K.A.; Ji, R.L.; Gallazzi, F.; Jiang, S.; Van Doren, S.R.; Tao, Y.X.; Newton Northup, J. Development of a therapeutic peptide for cachexia suggests a platform approach for drug-like peptides. ACS Pharmacol. Transl. Sci. 2022, 5, 344–361. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Xie, H.J.; Wang, W.; Wang, Z.Q.; Tao, Y.X. Pharmacology of the giant panda (Ailuropoda melanoleuca) melanocortin-3 receptor. Gen. Comp. Endocrinol. 2019, 277, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.C.; Sartin, J.L.; Tao, Y.X. Molecular cloning and pharmacological characterization of porcine melanocortin-3 receptor. J. Endocrinol. 2008, 196, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.X. Functional characterization of novel melanocortin-3 receptor mutations identified from obese subjects. Biochim. Biophys. Acta 2007, 1772, 1167–1174. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.X.; Huang, H.; Wang, Z.Q.; Yang, F.; Williams, J.N.; Nikiforovich, G.V. Constitutive activity of neural melanocortin receptors. Methods Enzymol. 2010, 484, 267–279. [Google Scholar]
- Renquist, B.J.; Zhang, C.; Williams, S.Y.; Cone, R.D. Development of an assay for high-throughput energy expenditure monitoring in the zebrafish. Zebrafish 2013, 10, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xue, S.; Lei, X.; Song, W.; Li, L.; Li, X.; Fu, Y.; Zhang, C.; Zhang, H.; Luo, Y.; et al. Pharmacological evaluation of melanocortin 2 receptor accessory protein 2 on axolotl neural melanocortin signaling. Front. Endocrinol. 2022, 13, 820896. [Google Scholar] [CrossRef]
- Zhang, H.J.; Cui, Z.H.; Liu, M.; Min, T.Q.; Xiao, X.; Wang, Z.Q.; Tao, Y.X. Pharmacological characterization of three chicken melanocortin-3 receptor mutants. Domest. Anim. Endocrinol. 2021, 74, 106507. [Google Scholar] [CrossRef]
- Srinivasan, S.; Lubrano-Berthelier, C.; Govaerts, C.; Picard, F.; Santiago, P.; Conklin, B.R.; Vaisse, C. Constitutive activity of the melanocortin-4 receptor is maintained by its N-terminal domain and plays a role in energy homeostasis in humans. J. Clin. Investig. 2004, 114, 1158–1164. [Google Scholar] [CrossRef] [Green Version]
- Ersoy, B.A.; Pardo, L.; Zhang, S.; Thompson, D.A.; Millhauser, G.; Govaerts, C.; Vaisse, C. Mechanism of N-terminal modulation of activity at the melanocortin-4 receptor GPCR. Nat. Chem. Biol. 2012, 8, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Nishi, S.; Nakabayashi, K.; Kobilka, B.; Hsueh, A.J.W. The ectodomain of the luteinizing hormone receptor interacts with exoloop 2 to constrain the transmembrane region. Studies using chimeric human and fly receptors. J. Biol. Chem. 2002, 277, 3958–3964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parma, J.; Van Sande, J.; Swillens, S.; Tonacchera, M.; Dumont, J.; Vassart, G. Somatic mutations causing constitutive activity of the thyrotropin receptor are the major cause of hyperfunctioning thyroid adenomas: Identification of additional mutations activating both the cyclic adenosine 3′,5′-monophosphate and inositol phosphate-Ca2+ cascades. Mol. Endocrinol. 1995, 9, 725–733. [Google Scholar] [PubMed] [Green Version]
- Zhang, M.; Tong, K.P.; Fremont, V.; Chen, J.; Narayan, P.; Puett, D.; Weintraub, B.D.; Szkudlinski, M.W. The extracellular domain suppresses constitutive activity of the transmembrane domain of the human TSH receptor: Implications for hormone-receptor interaction and antagonist design. Endocrinology 2000, 141, 3514–3517. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Li, L.; Jin, X.; Xu, B.; Pi, L.; Liu, S.; Zhu, W.; Zhang, C.; Luan, B.; Gong, L.; et al. Pharmacological effect of human melanocortin-2 receptor accessory protein 2 variants on hypothalamic melanocortin receptors. Endocrine 2018, 61, 94–104. [Google Scholar] [CrossRef]
- Wang, M.; Chen, Y.J.; Zhu, M.; Xu, B.X.; Guo, W.X.; Lyu, Y.S.; Zhang, C. Pharmacological modulation of melanocortin-4 receptor by melanocortin receptor accessory protein 2 in Nile tilapia. Gen. Comp. Endocrinol. 2019, 282, 113219. [Google Scholar] [CrossRef]
- Soletto, L.; Hernandez-Balfago, S.; Rocha, A.; Scheerer, P.; Kleinau, G.; Cerdá-Reverter, J.M. Melanocortin receptor accessory protein 2-induced adrenocorticotropic hormone response of human melanocortin 4 receptor. J. Endocr. Soc. 2019, 3, 314–323. [Google Scholar] [CrossRef] [Green Version]
- Rao, Y.Z.; Chen, R.; Zhang, Y.; Tao, Y.X. Orange-spotted grouper melanocortin-4 receptor: Modulation of signaling by MRAP2. Gen. Comp. Endocrinol. 2019, 284, 113234. [Google Scholar] [CrossRef]
- Tao, Y.X. Constitutive activation of G protein-coupled receptors and diseases: Insights into mechanism of activation and therapeutics. Pharmacol. Ther. 2008, 120, 129–148. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Forlano, P.M.; Cone, R.D. AgRP and POMC neurons are hypophysiotropic and coordinately regulate multiple endocrine axes in a larval teleost. Cell Metab. 2012, 15, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.X. Constitutive activity in melanocortin-4 receptor: Biased signaling of inverse agonists. Adv. Pharmacol. 2014, 70, 135–154. [Google Scholar] [CrossRef]
- Gillyard, T.; Fowler, K.; Williams, S.Y.; Cone, R.D. Obesity-associated mutant melanocortin-4 receptors with normal Gαs coupling frequently exhibit other discoverable pharmacological and biochemical defects. J. Neuroendocrinol. 2019, 31, e12795. [Google Scholar] [CrossRef] [PubMed]
- Black, D.L. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem. 2003, 72, 291–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef]
- Wahl, M.C.; Will, C.L.; Luhrmann, R. The spliceosome: Design principles of a dynamic RNP machine. Cell 2009, 136, 701–718. [Google Scholar] [CrossRef] [Green Version]
- Liang, Q.; Wu, N.; Zaneveld, S.; Liu, H.; Fu, S.; Wang, K.; Bertrand, R.; Wang, J.; Li, Y.; Chen, R. Transcript isoforms of Reep6 have distinct functions in the retina. Hum. Mol. Genet. 2021, 30, 1907–1918. [Google Scholar] [CrossRef] [PubMed]
- Webb, T.R.; Chan, L.; Cooray, S.N.; Cheetham, M.E.; Chapple, J.P.; Clark, A.J. Distinct melanocortin 2 receptor accessory protein domains are required for melanocortin 2 receptor interaction and promotion of receptor trafficking. Endocrinology 2009, 150, 720–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouault, A.A.J.; Lee, A.A.; Sebag, J.A. Regions of MRAP2 required for the inhibition of orexin and prokineticin receptor signaling. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 2322–2329. [Google Scholar] [CrossRef]
- Rouault, A.A.J.; Rosselli-Murai, L.K.; Hernandez, C.C.; Gimenez, L.E.; Tall, G.G.; Sebag, J.A. The GPCR accessory protein MRAP2 regulates both biased signaling and constitutive activity of the ghrelin receptor GHSR1a. Sci. Signal. 2020, 13, eaax4569. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MC3R | IC50 (nM) | cMC3R | hMC3R |
|---|---|---|---|
| Bmax (%) | 247.97 ± 13.44 b | 100 | |
| NDP-MSH | IC50 (nM) | 4.37 ± 0.72 a | 1.98 ± 0.43 |
| α-MSH | IC50 (nM) | 240.42 ± 37.34 | 221.57 ± 30.23 |
| β-MSH | IC50 (nM) | 140.18 ± 31.37 | 161.56 ± 31.45 |
| ACTH | IC50 (nM) | 58.52 ± 8.67 | 45.71 ± 5.23 |
| D-Trp8-γ-MSH | IC50 (nM) | 39.30 ± 0.30 b | 24.62 ± 1.25 |
| MC3R | EC50/Rmax | cMC3R | hMC3R |
|---|---|---|---|
| Basal (%) | 100.72 ± 7.69 | 100 | |
| NDP-MSH | EC50 (nM) | 0.40 ± 0.16 | 0.24 ± 0.11 |
| Rmax (%) | 199.38 ± 28.06 a | 100 | |
| α-MSH | EC50 (nM) | 0.97 ± 0.35 | 1.49 ± 0.22 |
| Rmax (%) | 145.69 ± 14.30 | 100 | |
| β-MSH | EC50 (nM) | 1.04 ± 0.23 | 1.46 ± 0.55 |
| Rmax (%) | 229.77 ± 46.45 a | 100 | |
| ACTH | EC50 (nM) | 1.46 ± 0.30 | 2.12 ± 0.69 |
| Rmax (%) | 182.83 ± 25.21 a | 100 | |
| D-Trp8-γ-MSH | EC50 (nM) | 1.08 ± 0.15 | 0.71 ± 0.19 |
| Rmax (%) | 188.25 ± 19.56 a | 100 |
| cMC3R/cMRAPs | Bmax (%) | α-MSH | ACTH |
|---|---|---|---|
| IC50 (nM) | IC50 (nM) | ||
| cMC3R | 100 | 106.68 ± 13.80 | 55.56 ± 11.64 |
| cMC3R/cMRAP1 | 81.81 ± 5.01 a | 127.76 ± 30.47 | 68.59 ± 9.31 |
| cMC3R/cMRAP2a | 148.43 ± 14.94 a | 118.61 ± 22.25 | 57.58 ± 4.09 |
| cMC3R/cMRAP2b | 128.42 ± 9.06 a | 148.03 ± 30.16 | 49.59 ± 6.77 |
| cMC3R/cMRAPs | Basal (%) | α-MSH | ACTH | ||
|---|---|---|---|---|---|
| EC50 (nM) | Rmax (%) | EC50 (nM) | Rmax (%) | ||
| cMC3R | 100 | 3.08 ± 1.26 | 100 | 2.02 ± 0.39 | 100 |
| cMC3R/cMRAP1 | 100.91 ± 16.74 | 0.88 ± 0.18 | 46.67 ± 8.88 b | 2.11 ± 0.56 | 46.61 ± 12.37 a |
| cMC3R/cMRAP2a | 84.71 ± 19.03 | 1.37 ± 0.26 | 73.02 ± 5.81 a | 2.18 ± 0.39 | 73.79 ± 4.94 a |
| cMC3R/cMRAP2b | 68.88 ± 9.35 a | 3.64 ± 1.11 | 66.93 ± 9.64 a | 3.41 ± 0.99 | 85.02 ± 11.04 |
| cMC4R/cMRAPs | Bmax (%) | α-MSH | ACTH |
|---|---|---|---|
| IC50 (nM) | IC50 (nM) | ||
| cMC4R | 100 | 504.66 ± 124.33 | 348.29 ± 94.51 |
| cMC4R/cMRAP1 | 151.36 ± 10.67 a | 186.62 ± 13.60 a | 37.63 ± 6.84 b |
| cMC4R/cMRAP2a | 167.03 ± 11.83 c | 228.40 ± 40.74 a | 35.63 ± 8.10 b |
| cMC4R/cMRAP2b | 121.60 ± 10.06 | 120.19 ± 22.41 b | 16.93 ± 5.69 c |
| cMC4R/cMRAPs | Basal (%) | α-MSH | ACTH | ||
|---|---|---|---|---|---|
| EC50 (nM) | Rmax (%) | EC50 (nM) | Rmax (%) | ||
| cMC4R | 100 | 1.18 ± 0.21 | 100 | 0.92 ± 0.41 | 100 |
| cMC4R/cMRAP1 | 53.89 ± 4.06 b | 1.37 ± 0.24 | 99.80 ± 3.80 | 1.92 ± 0.40 | 76.21 ± 13.29 |
| cMC4R/cMRAP2a | 74.70 ± 4.28 b | 0.90 ± 0.10 | 87.00 ± 10.28 | 3.82 ± 1.25 | 144.41 ± 13.36 a |
| cMC4R/cMRAP2b | 55.82 ± 3.43 b | 0.63 ± 0.10 | 52.62 ± 7.93 b | 1.20 ± 0.44 | 55.87 ± 9.82 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, R.-L.; Jiang, S.-S.; Tao, Y.-X. Modulation of Canine Melanocortin-3 and -4 Receptors by Melanocortin-2 Receptor Accessory Protein 1 and 2. Biomolecules 2022, 12, 1608. https://doi.org/10.3390/biom12111608
Ji R-L, Jiang S-S, Tao Y-X. Modulation of Canine Melanocortin-3 and -4 Receptors by Melanocortin-2 Receptor Accessory Protein 1 and 2. Biomolecules. 2022; 12(11):1608. https://doi.org/10.3390/biom12111608
Chicago/Turabian StyleJi, Ren-Lei, Shan-Shan Jiang, and Ya-Xiong Tao. 2022. "Modulation of Canine Melanocortin-3 and -4 Receptors by Melanocortin-2 Receptor Accessory Protein 1 and 2" Biomolecules 12, no. 11: 1608. https://doi.org/10.3390/biom12111608
APA StyleJi, R. -L., Jiang, S. -S., & Tao, Y. -X. (2022). Modulation of Canine Melanocortin-3 and -4 Receptors by Melanocortin-2 Receptor Accessory Protein 1 and 2. Biomolecules, 12(11), 1608. https://doi.org/10.3390/biom12111608