
Construction of Multiple Logic Circuits Based on Allosteric DNAzymes




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemical Reagents
2.2. DNA Assembly
2.3. Native PAGE
2.4. Measurement of Fluorescence Spectroscopy
3. Results
3.1. Mechanism of the Allosteric Strategy and DNAzyme Assemblying
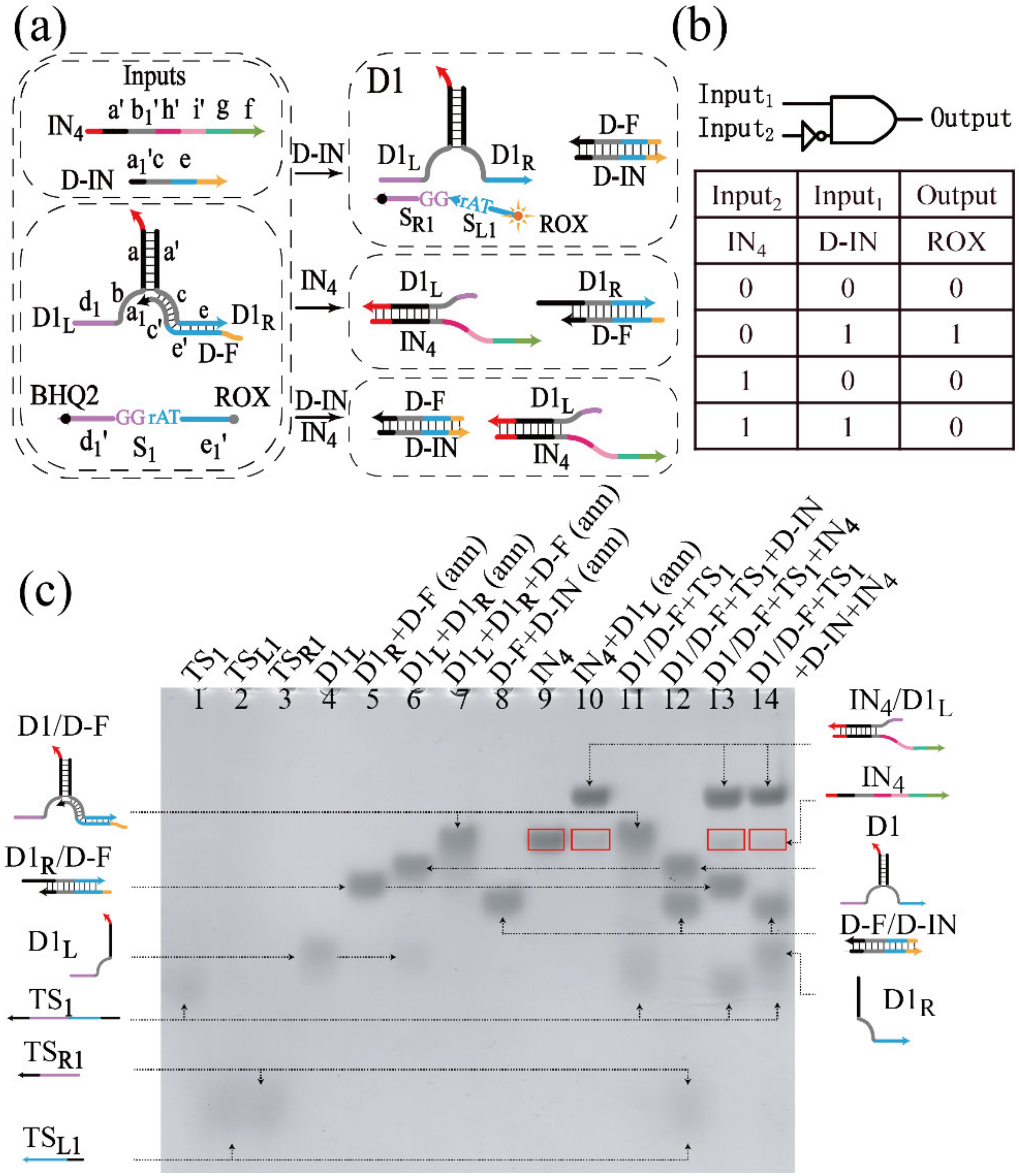
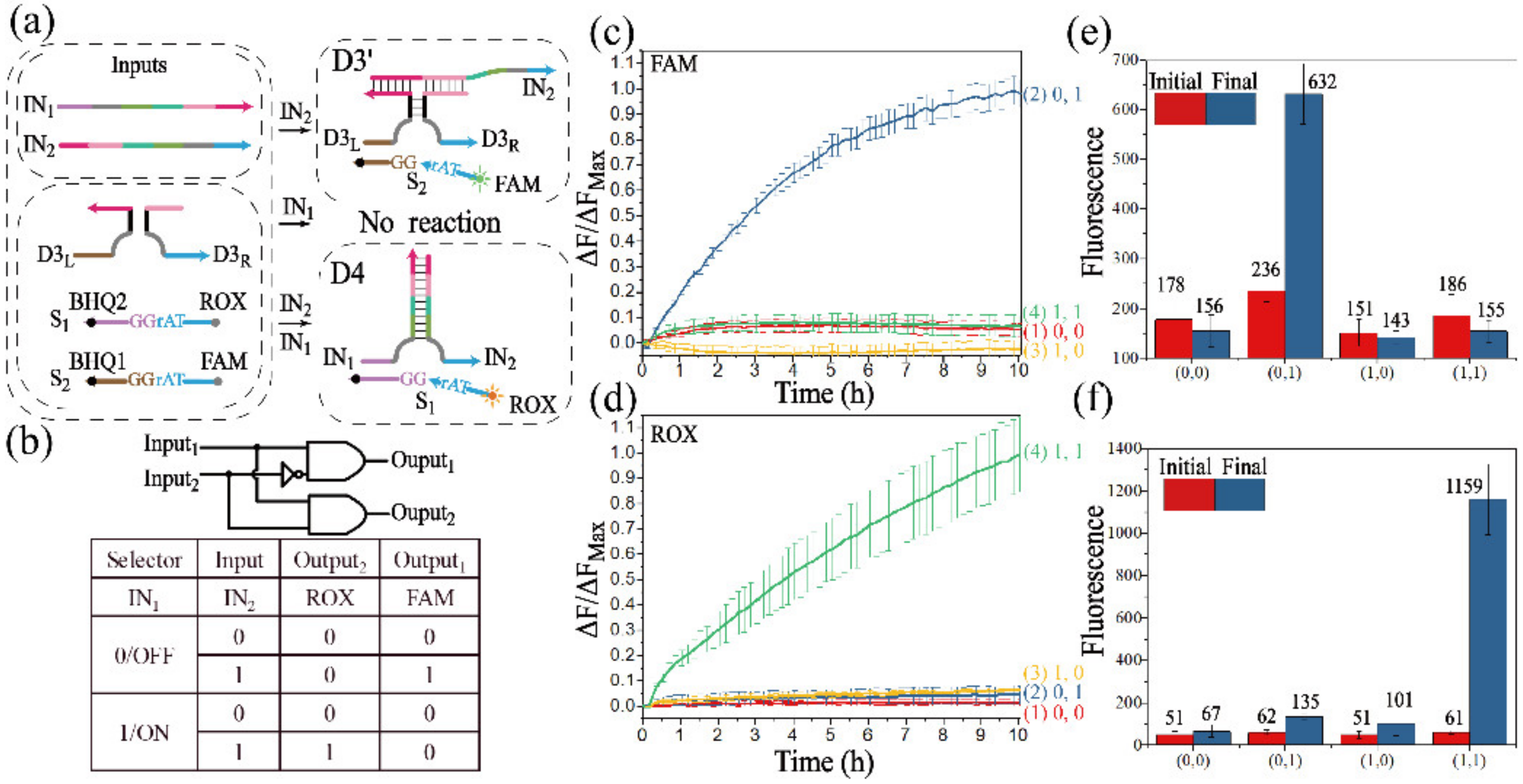
3.1.1. Realization of the INH Gate
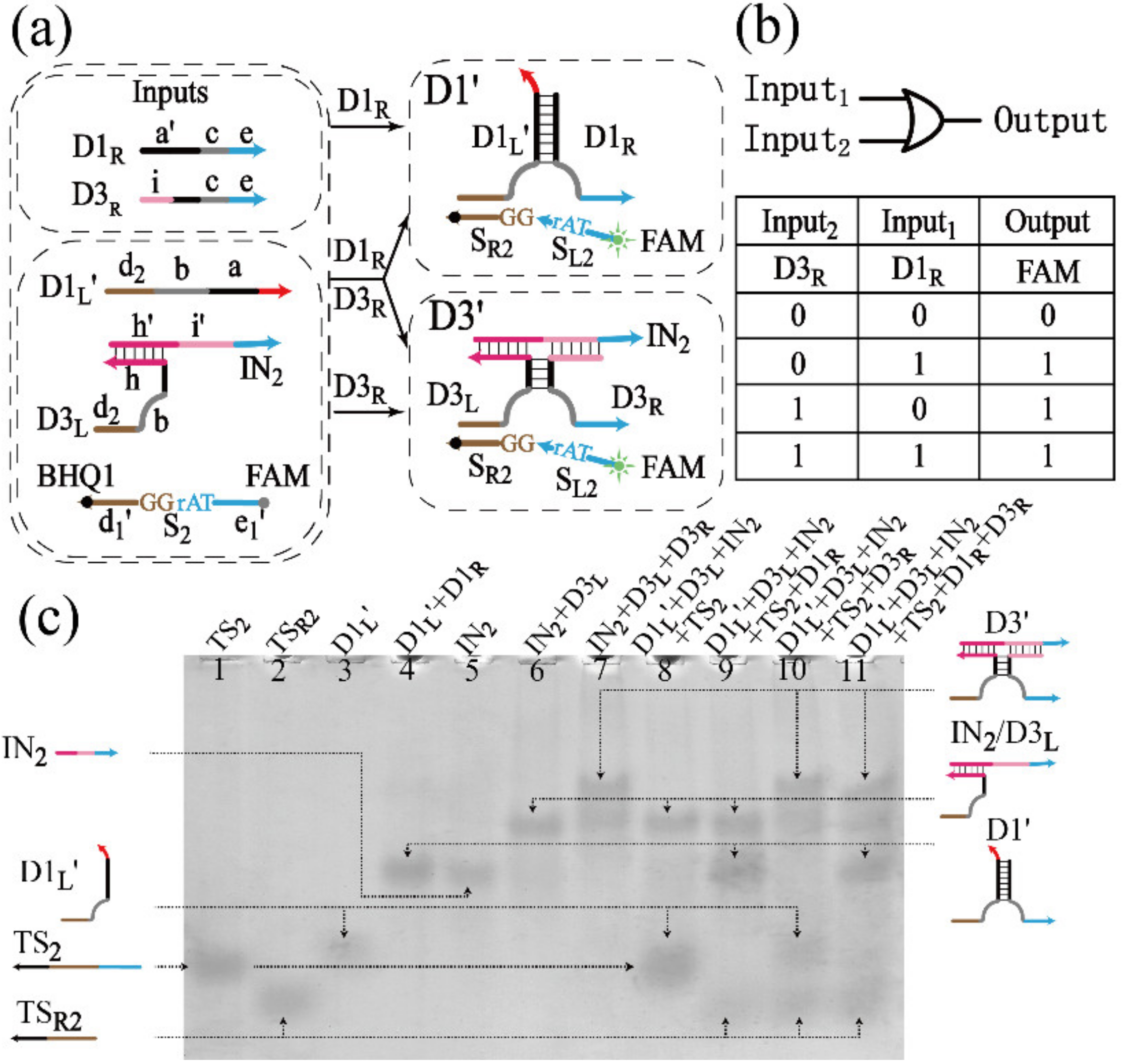
3.1.2. Realization of the OR Gate
3.2. Implrmentation of Arithmetic Functions
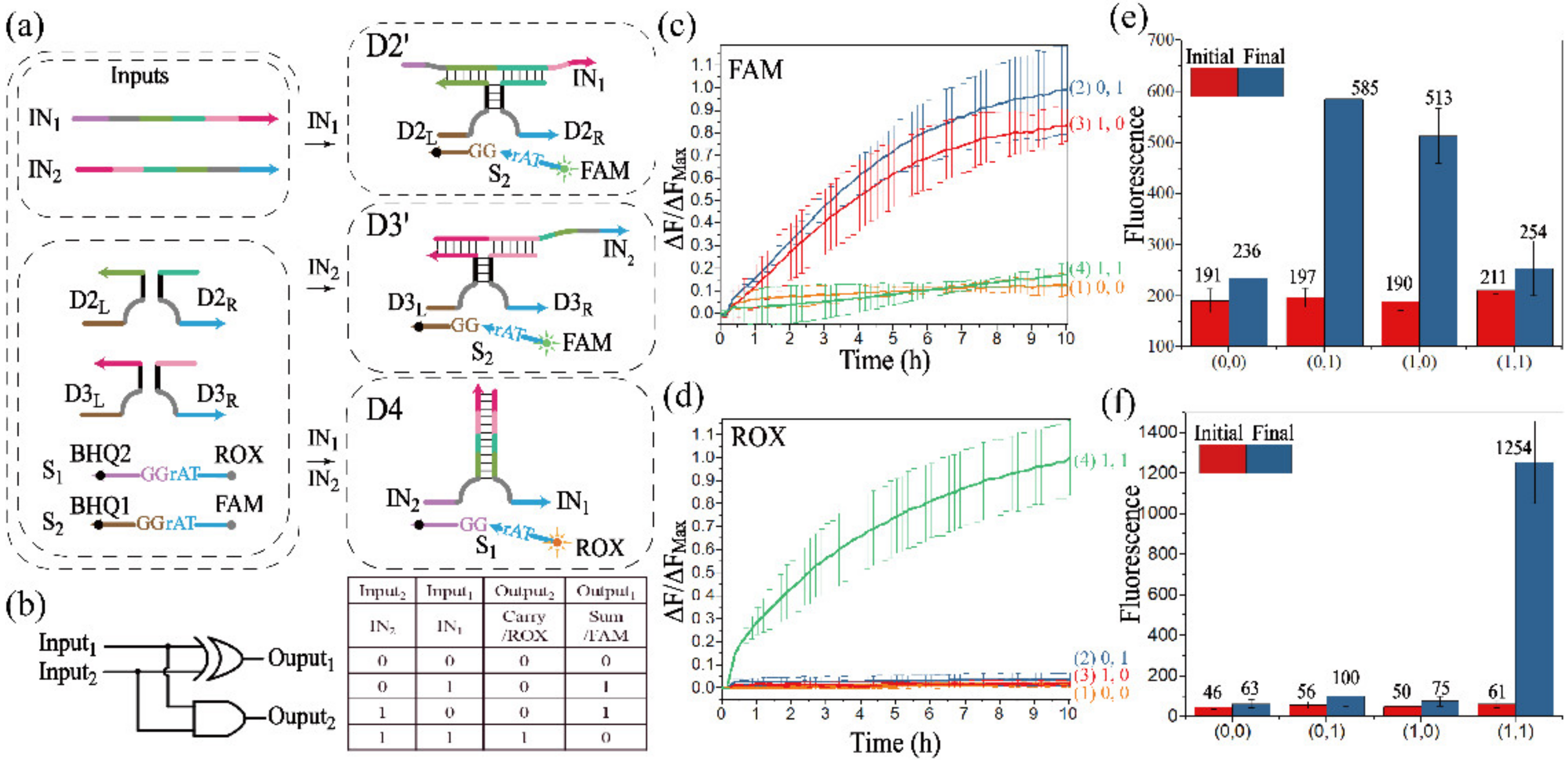
3.2.1. Realization of the Half-Adder
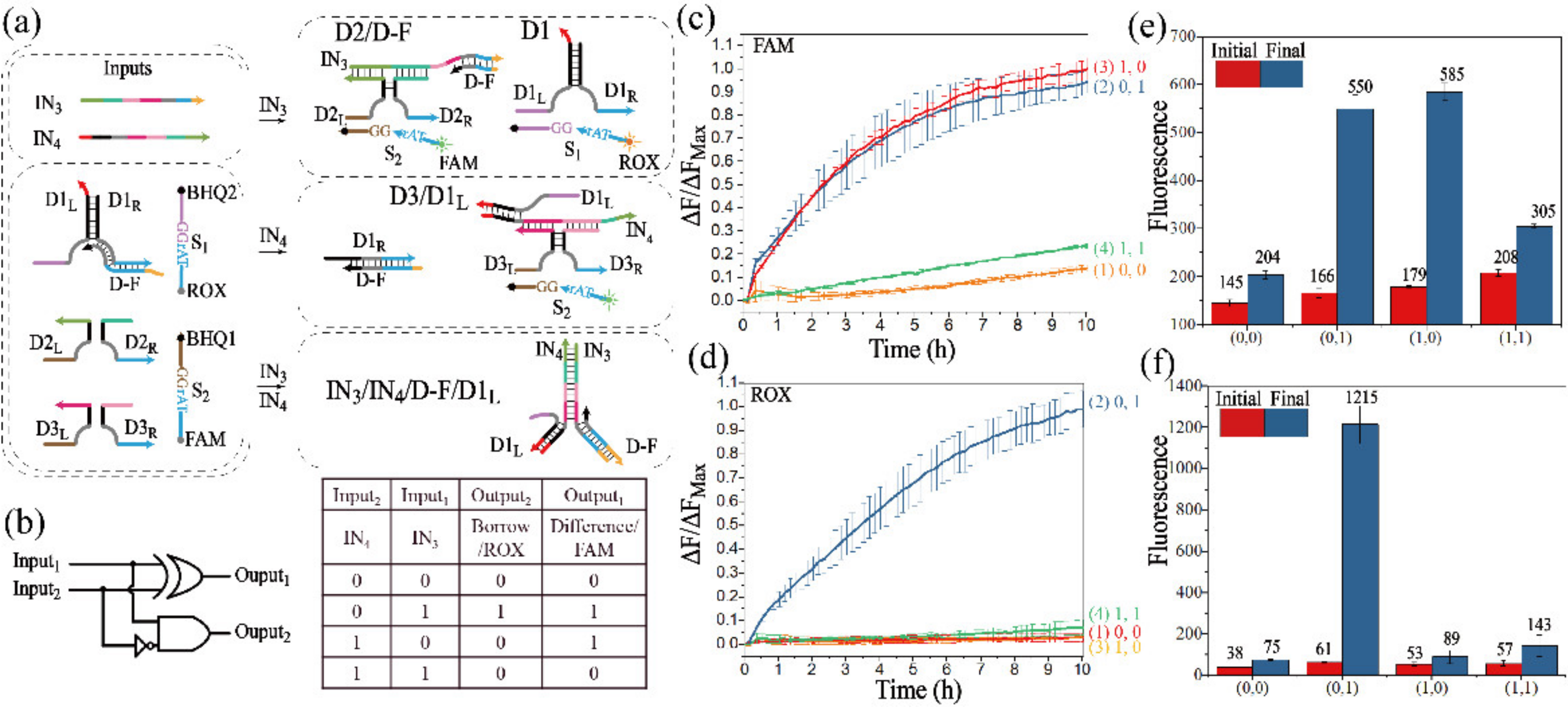
3.2.2. Realization of the Half-Subtractor
3.3. Implementation of Non-Arithmetic Functions
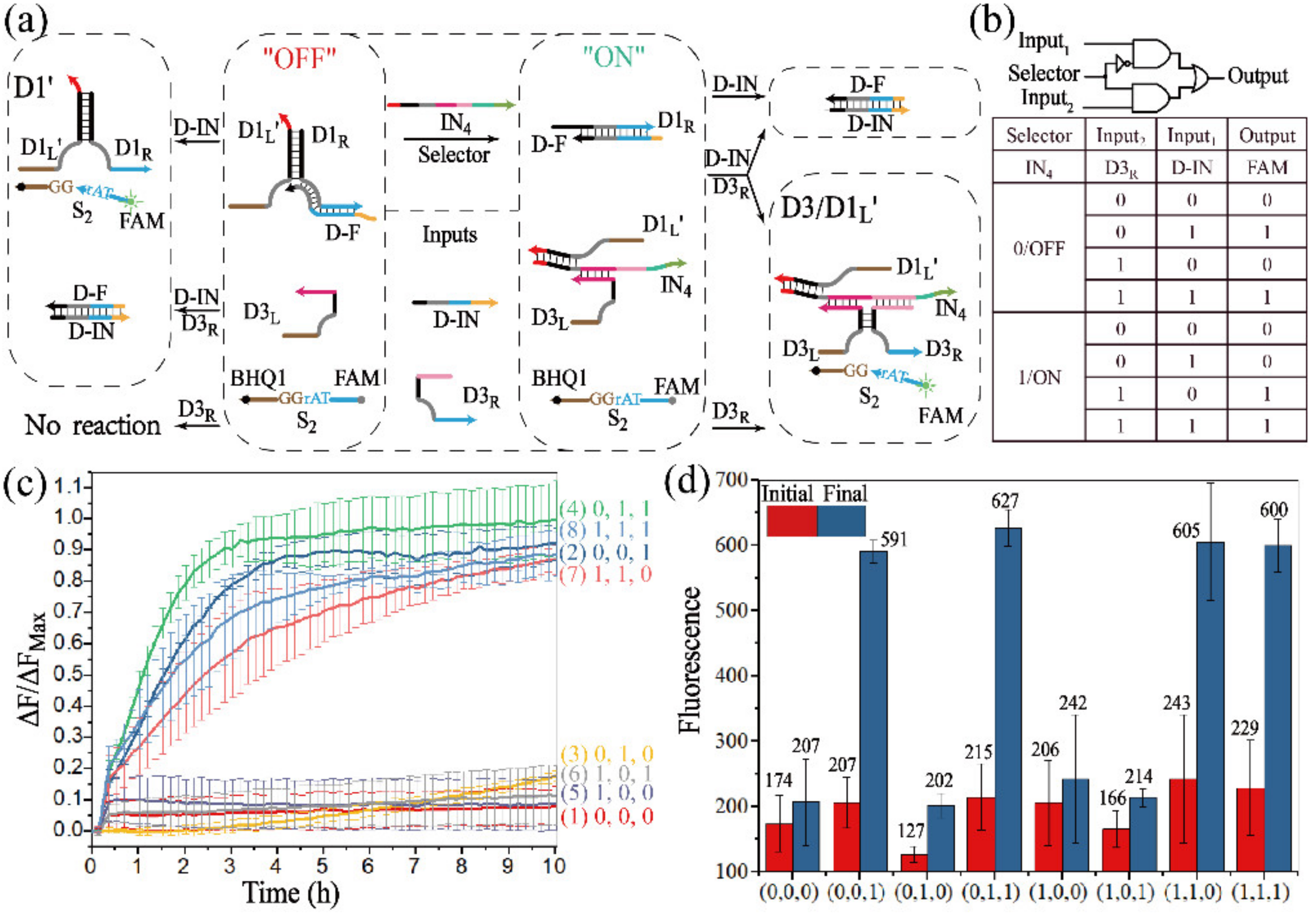
3.3.1. Realization of the 2:1 MUX
3.3.2. Realization of the 1:2 DEMUX
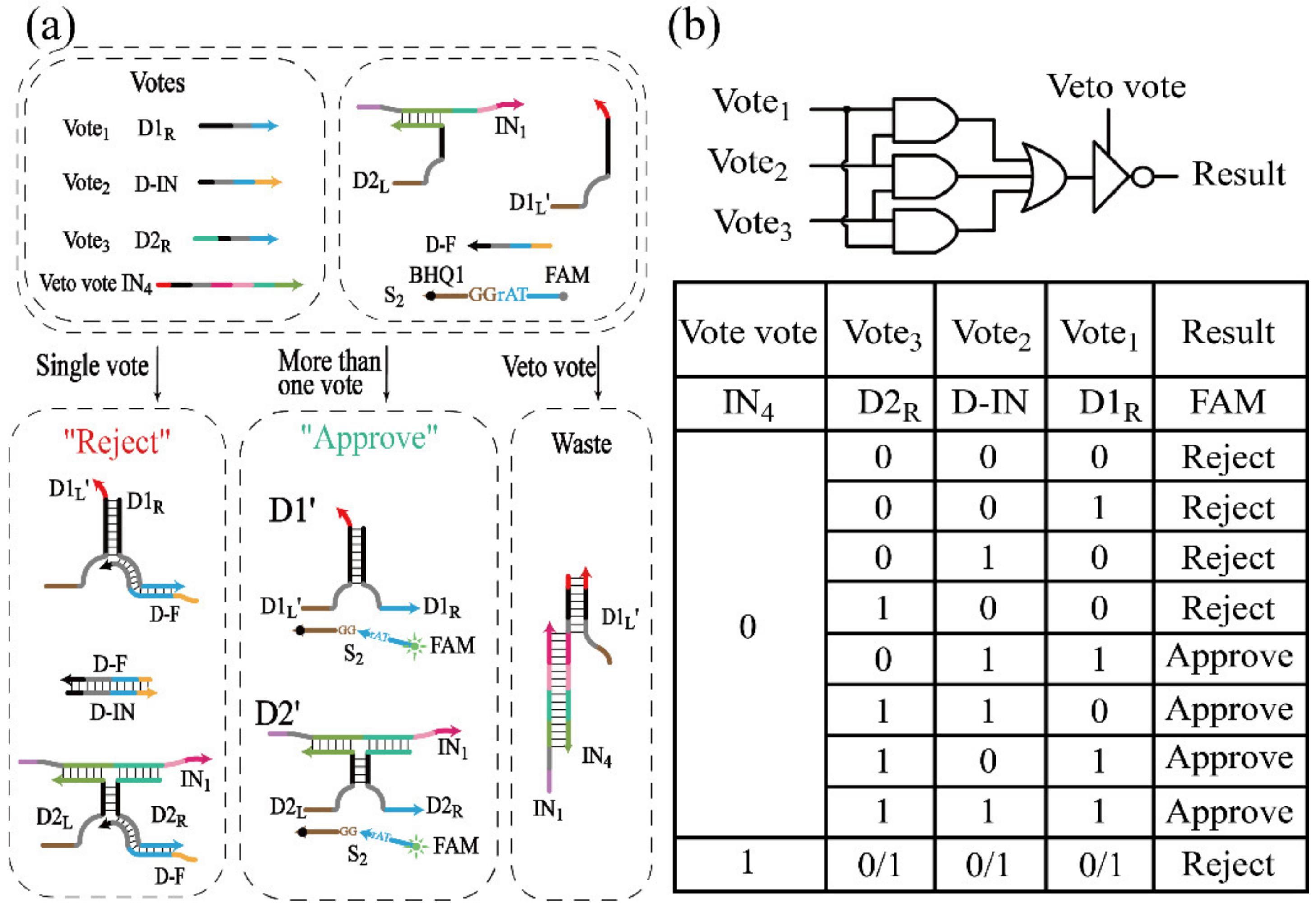
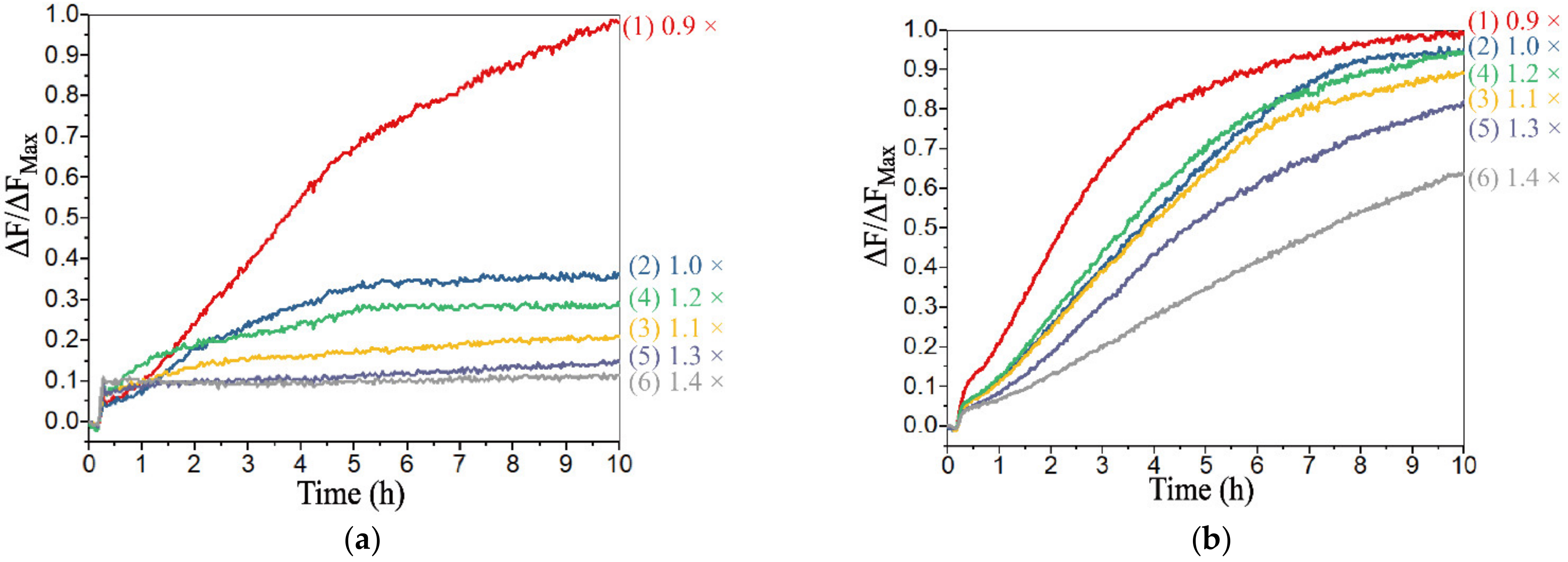
3.4. Realization of Threshold Control and DNA Voter with One-Vote Veto Function
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abendroth, J.M.; Bushuyev, O.S.; Weiss, P.S.; Barrett, C.J. Controlling Motion at the Nanoscale: Rise of the Molecular Machines. ACS Nano 2015, 9, 7746–7768. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Reif, J. Nucleic Acid Databases and Molecular-Scale Computing. ACS Nano 2019, 13, 6256–6268. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Lu, D.; Liang, H.; Xie, S.; Zhang, X.; Liu, Q.; Yuan, Q.; Tan, W. mRNA-Initiated, Three-Dimensional DNA Amplifier Able to Function inside Living Cells. J. Am. Chem. Soc. 2018, 140, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Jiang, J.; Yuan, R.; Zhuo, Y.; Chai, Y. Highly Ordered and Field-Free 3D DNA Nanostructure: The Next Generation of DNA Nanomachine for Rapid Single-Step Sensing. J. Am. Chem. Soc. 2018, 140, 9361–9364. [Google Scholar] [CrossRef] [PubMed]
- Deepak, S.; Manojkumar, R. DNA Computing: Methodologies and Challenges. In DNA- and RNA-Based Computing Systems, 1st ed.; Evgeny, K., Ed.; Wiley-VCH: Weinheim, Germany, 2021; pp. 15–30. [Google Scholar]
- Geng, H.; Zhou, C.; Guo, C. DNA-based digital comparator systems constructed by multifunctional nanoswitches. Nanoscale 2019, 11, 21856–21866. [Google Scholar] [CrossRef]
- Cao, B.; Li, X.; Zhang, X.; Wang, B.; Zhang, Q.; Wei., X. Designing Uncorrelated Address Constrain for DNA Storage by DMVO Algorithm. IEEE/ACM Trans. Comput. Biol. Bioinform. 2020, 1. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Zhang, X.; Wu, J.; Wang, B.; Zhang, Q.; Wei, X. Minimum free energy coding for DNA storage. IEEE Trans. NanoBioscience 2021, 20, 212–222. [Google Scholar] [CrossRef]
- Xiao, M.; Lai, W.; Wang, F.; Li, L.; Fan, C.; Pei, H. Programming Drug Delivery Kinetics for Active Burst Release with DNA Toehold Switches. J. Am. Chem. Soc. 2019, 141, 20354–20364. [Google Scholar] [CrossRef]
- Li, S.; Jiang, Q.; Liu, S.; Zhang, Y.; Tian, Y.; Song, C.; Wang, J.; Zou, Y.; Anderson, G.J.; Han, J.Y.; et al. A DNA nanorobot functions as a cancer therapeutic in response to a molecular trigger in vivo. Nat. Biotechnol. 2018, 36, 258–264. [Google Scholar] [CrossRef]
- Kamar, O.; Sun, S.C.; Lin, C.H.; Chung, W.Y.; Lee, M.S.; Liao, Y.C.; Kolpashchikov, D.M.; Chuang, M.C. A mutation-resistant deoxyribozyme OR gate for highly selective detection of viral nucleic acids. Chem Commun. 2017, 53, 10592–10595. [Google Scholar] [CrossRef]
- Zhong, R.; Xiao, M.; Zhu, C.; Shen, X.; Tang, Q.; Zhang, W.; Wang, L.; Song, S.; Qu, X.; Pei, H.; et al. Logic Catalytic Interconversion of G-Molecular Hydrogel. ACS Appl. Mater. Interfaces 2018, 10, 4512–4518. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, X.; Xia, X.; Xiang, H. Catalytic hairpin assembly-based double-end DNAzyme cascade-feedback amplification for sensitive fluorescence detection of HIV-1 DNA. Anal. Chim. Acta 2020, 1096, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Seo, M.W.; Jung, C.; Lee, J.Y.; Park, H.G. Simple and universal platform for logic gate operations based on molecular beacon probes. Small 2012, 8, 2203–2212. [Google Scholar] [CrossRef]
- Lin, X.; Liu, Y.; Deng, J.; Lyu, Y.; Qian, P.; Li, Y.; Wang, S. Multiple advanced logic gates made of DNA-Ag nanocluster and the application for intelligent detection of pathogenic bacterial genes. Chem. Sci. 2018, 9, 1774–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Yu, L.; Yang, S.; Tang, X.; Chang, K.; Chen, M. Boolean logic gate based on DNA strand displacement for biosensing: Current and emerging strategies. Nanoscale Horiz. 2021, 6, 298–310. [Google Scholar] [CrossRef]
- Bi, S.; Yue, S.; Zhang, S. Hybridization chain reaction: A versatile molecular tool for biosensing, bioimaging, and biomedicine. Chem. Sov. Rev. 2017, 46, 4281–4298. [Google Scholar] [CrossRef] [PubMed]
- Jin, R.; Zeng, C.; Zhou, M.; Chen, Y. Atomically Precise Colloidal Metal Nanoclusters and Nanoparticles: Fundamentals and Opportunities. Chem. Rev. 2016, 116, 10346–10413. [Google Scholar] [CrossRef]
- Seeling, G.; Soloveichik, D.; Zhang, D.Y.; Winfree, E. Enzyme-free nucleic acid logic circuits. Science 2006, 314, 1585–1588. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, Q.; Liu, Y.; Wang, B.; Zhou, S. A molecular device: A DNA molecular lock driven by the nicking enzymes. Comput. Struct. Biotechnol. J. 2020, 18, 2107–2116. [Google Scholar] [CrossRef]
- Pan, L.; Wang, Z.; Li, Y.; Xu, F.; Zhang, Q.; Zhang, C. Nicking enzyme-controlled toehold regulation for DNA logic circuits. Nanoscale 2017, 9, 18223–18228. [Google Scholar] [CrossRef]
- Lv, M.; Zhou, W.; Fan, D.; Guo, Y.; Zhu, X.; Ren, J.; Wang, E. Illuminating Diverse Concomitant DNA Logic Gates and Concatenated Circuits with Hairpin DNA-Templated Silver Nanoclusters as Universal Dual-Output Generators. Adv. Mater. 2020, 32, e1908480. [Google Scholar] [CrossRef] [PubMed]
- Qing, Z.; He, X.; He, D.; Wang, K.; Xu, F.; Qing, T.; Yang, X. Poly(thymine)-Templated Selective Formation of Fluorescent Copper Nanoparticles. Angew. Chem. Int. Ed. Engl. 2013, 52, 9719–9722. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Zhu, J.; Liu, Y.; Wang, E.; Dong, S. Label-free and enzyme-free platform for the construction of advanced DNA logic devices based on the assembly of graphene oxide and DNA-templated AgNCs. Nanoscale 2016, 8, 3834–3840. [Google Scholar] [CrossRef]
- Bader, A.; Cockroft, S.L. Simultaneous G-Quadruplex DNA Logic. Chemistry 2018, 24, 4820–4824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Yue, S.; Zhao, T.; Luo, B.; Bi, S. Exonuclease-assisted target recycling amplification for label-free chemiluminescence assay and molecular logic operations. Chem. Commun. 2017, 53, 12201–12204. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, J.; Jiang, S.; Liu, Y.; Yan, H. DNAzyme-Based Logic Gate-Mediated DNA Self-Assembly. Nano Lett. 2016, 16, 736–741. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, G.; Dalchau, N.; Muscat, R.A.; Phillips, A.; Seelig, G. A spatially localized architecture for fast and modular DNA computing. Nat. Nanotechnol. 2017, 12, 920–927. [Google Scholar] [CrossRef]
- Yang, J.; Wu, R.; Li, Y.; Wang, Z.; Pan, L.; Zhang, Q.; Lu, Z.; Zhang, C. Entropy-driven DNA logic circuits regulated by DNAzyme. Nucleic Acids Res. 2018, 46, 8532–8541. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Chang, D.; Li, Y. Discovery and Biosensing Applications of Diverse RNA-Cleaving DNAzymes. Acc. Chem. Res. 2017, 50, 2273–2283. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, Q.; Chang, D.; Gu, J.; Brennan, J.D.; Li, Y. A DNAzyme Feedback Amplification Strategy for Biosensing. Angew. Chem. Int. Ed. Engl. 2017, 56, 6142–6146. [Google Scholar] [CrossRef]
- Chandrasekaran, A.R.; Levchenko, O.; Patel, D.S.; MacIsaac, M.; Halvorsen, K. Addressable configurations of DNA nanostructures for rewritable memory. Nucleic Acids Res. 2017, 45, 11459–11465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, B.I.; Pollak, N.M.; Stefanovic, D.; Macdonald, J. Repeated Reuse of Deoxyribozyme-Based Logic Gates. Nano Lett. 2019, 19, 7655–7661. [Google Scholar] [CrossRef] [PubMed]
- Orbach, R.; Mostinski, L.; Wang, F.; Willner, I. Nucleic acid driven DNA machineries synthesizing Mg2+-dependent DNAzymes: An interplay between DNA sensing and logic-gate operations. Chem. Eur. J. 2012, 18, 14689–14694. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lu, Y. A Highly Sensitive and Selective Catalytic DNA Biosensor for Lead Ions. J. Am. Chem. Soc. 2000, 122, 10466–10467. [Google Scholar] [CrossRef]
- Elbaz, J.; Lioubashevski, O.; Wang, F.; Remacle, F.; Levine, R.D.; Willner, I. DNA computing circuits using libraries of DNAzyme subunits. Nat. Nanotechnol. 2010, 5, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Mokany, E.; Bone, S.M.; Young, P.E.; Doan, T.B.; Todd, A.V. MNAzymes, a Versatile New Class of Nucleic Acid Enzymes That Can Function as Biosensors and Molecular Switches. J. Am. Chem. Soc. 2010, 132, 1051–1059. [Google Scholar] [CrossRef]
- Chen, Y.; Song, Y.; He, Z.; Wang, Z.; Liu, W.; Wang, F.; Zhang, X.; Zhou, X. pH-controlled DNAzymes: Rational design and their applications in DNA-machinery devices. Nano Res. 2016, 9, 3084–3092. [Google Scholar] [CrossRef]
- Zhou, W.; Saran, R.; Chen, Q.; Ding, J.; Liu, J. A New Na+-Dependent RNA-Cleaving DNAzyme with over 1000-fold Rate Acceleration by Ethanol. ChemBioChem 2016, 17, 159–163. [Google Scholar] [CrossRef]
- Saran, R.; Liu, J. A Silver DNAzyme. Anal. Chem. 2016, 88, 4014–4020. [Google Scholar] [CrossRef]
- Zheng, X.; Yang, J.; Zhou, C.; Zhang, C.; Zhang, Q.; Wei, X. Allosteric DNAzyme-based DNA logic circuit: Operations and dynamic analysis. Nucleic Acids Res. 2019, 47, 1097–1109. [Google Scholar] [CrossRef] [Green Version]
- Stojanovic, M.N.; Mitchell, T.E.; Stefanovic, D. Deoxyribozyme-based logic gates. J. Am. Chem. Soc. 2002, 124, 3555–3561. [Google Scholar] [CrossRef]
- Orbach, R.; Remacle, F.; Levine, R.D.; Willner, I. DNAzyme-based 2:1 and 4:1 multiplexers and 1:2 demultiplexer. Chem. Sci. 2014, 5, 1074–1081. [Google Scholar] [CrossRef]
- Chen, J.; Pan, J.; Chen, S. A label-free and enzyme-free platform with a visible output for constructing versatile logic gates using caged G-quadruplex as the signal transducer. Chem. Sci. 2018, 9, 300–306. [Google Scholar] [CrossRef] [Green Version]
- Carr, C.E.; Marky, L.A. Melting Behavior of a DNA Four-Way Junction Using Spectroscopic and Calorimetric Techniques. J. Am. Chem. Soc. 2017, 139, 14443–14455. [Google Scholar] [CrossRef]
- Shlyakhtenko, L.S.; Potaman, V.N.; Sinden, R.R.; Gall, A.A.; Lyubchenko, Y.L. Structure and dynamics of three-way DNA junctions: Atomic force microscopy studies. Nucleic Acids Res. 2000, 28, 3472–3477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bone, S.M.; Todd, A.V. MNAzymes provide a universal mechanism for triggering DNAzyme synthesis cascades. Chem. Commun. 2014, 50, 13243–13246. [Google Scholar] [CrossRef]
- Fan, D.; Wang, K.; Zhu, J.; Xia, Y.; Han, Y.; Liu, Y.; Wang, E. DNA-based visual majority logic gate with one-vote veto function. Chem. Sci. 2015, 6, 1973–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, D.; Wang, E.; Dong, S. An intelligent universal system yields double results with half the effort for engineering a DNA “Contrary Logic Pairs” library and various DNA combinatorial logic circuits. Mater. Horiz. 2017, 4, 924–931. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Zhang, Q.; Zhang, X.; Liu, Y.; Yao, Y.; Kasabov, N. Construction of Multiple Logic Circuits Based on Allosteric DNAzymes. Biomolecules 2022, 12, 495. https://doi.org/10.3390/biom12040495
Liu X, Zhang Q, Zhang X, Liu Y, Yao Y, Kasabov N. Construction of Multiple Logic Circuits Based on Allosteric DNAzymes. Biomolecules. 2022; 12(4):495. https://doi.org/10.3390/biom12040495
Chicago/Turabian StyleLiu, Xin, Qiang Zhang, Xun Zhang, Yuan Liu, Yao Yao, and Nikola Kasabov. 2022. "Construction of Multiple Logic Circuits Based on Allosteric DNAzymes" Biomolecules 12, no. 4: 495. https://doi.org/10.3390/biom12040495
APA StyleLiu, X., Zhang, Q., Zhang, X., Liu, Y., Yao, Y., & Kasabov, N. (2022). Construction of Multiple Logic Circuits Based on Allosteric DNAzymes. Biomolecules, 12(4), 495. https://doi.org/10.3390/biom12040495