
Gal-2 Increases H3K4me3 and H3K9ac in Trophoblasts and Preeclampsia

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample for Statistical Analyses
2.2. Cell Culture
2.3. Immunocytochemical Staining
2.3.1. Staining Protocol for the BeWo Cells
2.3.2. Staining Protocol for the HVT Cells
2.4. Evaluation of Staining
2.5. Statistical Analysis
3. Results
3.1. The Histone Modifications H3K4me3 and H3K9ac Correlate Significantly with Gal-2
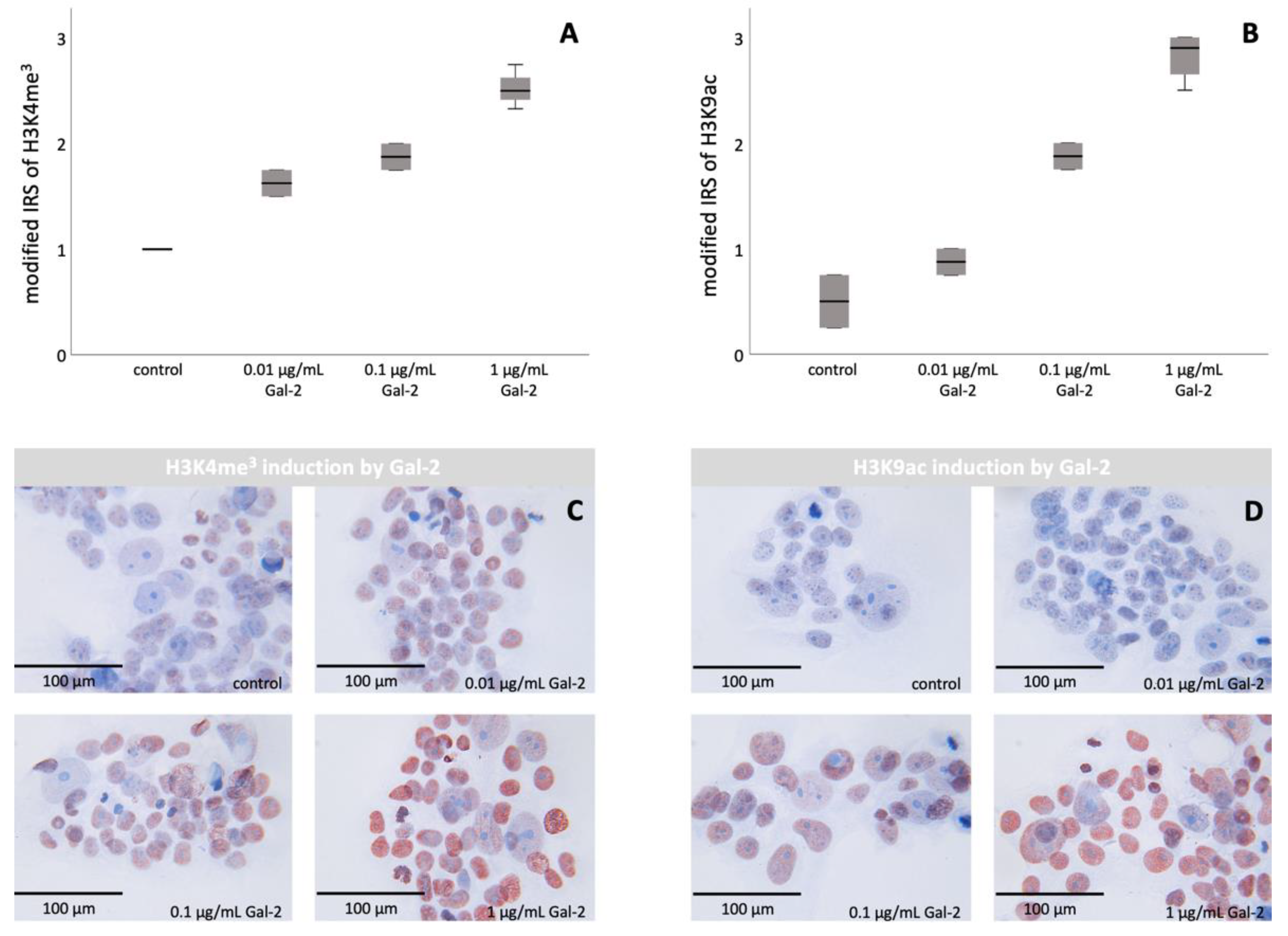
3.2. Gal-2 Promotes the Histone Modifications H3K4me3 and H3K9ac in BeWo Cells
3.3. Gal-2 Promotes the Histone Modifications H3K4me3 and H3K9ac in HVT Cells
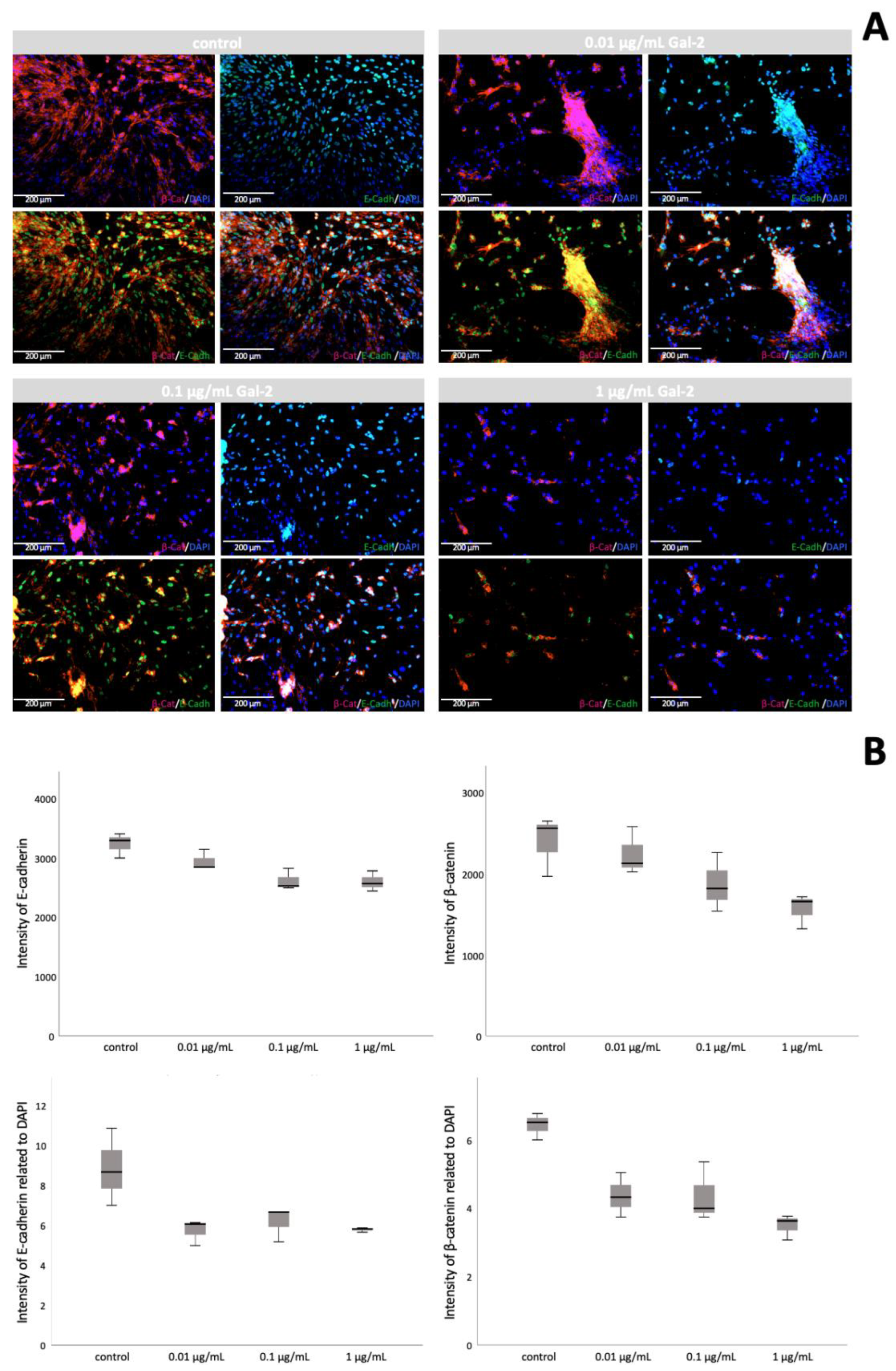
3.4. Gal-2 Promotes Cell Fusion in HVT Cell Culture
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BeWo cells | Trophoblast cell line, originates from human chorioncarcinoma |
| DAPI | 4′,6-diamidino-2-phenylindole |
| EVT | Extravillous trophoblast |
| FCS | Fetal calf serum |
| Gal-1 | Galectin-1 |
| Gal-2 | Galectin-2 |
| H3K14 | Histone H3 protein, modified at the 14th lysine residue |
| H3K27 | Histone H3 protein, modified at the 27th lysine residue |
| H3K4me3 | Trimethylated 4th lysine residue of the histone H3 protein |
| H3K9ac | Acetylated 9th lysine residue of the histone H3 protein |
| HDAC | Histone deacetlyase |
| HVT cells | Human villous trophoblast cell line |
| IRS | International Remmele Score |
| LGALS1 | Gene coding for galectin-1 |
| PE | Preeclampsia |
| RPMI-1640 | Roswell Park Memorial Institute 1640 cell culture medium |
References
- Hypertensive Pregnancy Disorders: Diagnosis and Therapy. Guideline of the German Society of Gynecology and Obstetrics (S2k-Level, AWMF-Registry No. 015/018, March 2019). Available online: http://www.awmf.org/leitlinien/detail/II/015-018.html (accessed on 2 April 2022).
- Han, C.; Han, L.; Huang, P.; Chen, Y.; Wang, Y.; Xue, F. Syncytiotrophoblast-derived extracellular vesicles in pathophysiology of preeclampsia. Front. Physiol. 2019, 10, 1236. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Gohner, C.; San Martin, S.; Vattai, A.; Hutter, S.; Parraga, M.; Jeschke, U.; Schleussner, E.; Markert, U.R.; Fitzgerald, J. Unique trophoblast stem cell- and pluripotency marker staining patterns depending on gestational age and placenta-associated pregnancy complications. Cell Adh. Migr. 2016, 10, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Lee, M.; Jeyabalan, A.; Roberts, J.M. The relationship of hypovitaminosis D and IL-6 in preeclampsia. Am. J. Obs. Gynecol. 2014, 210, 149.e1–149.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petsas, G.; Jeschke, U.; Richter, D.U.; Minas, V.; Hammer, A.; Kalantaridou, S.; Toth, B.; Tsatsanis, C.; Friese, K.; Makrigiannakis, A. Aberrant expression of corticotropin-releasing hormone in pre-eclampsia induces expression of FasL in maternal macrophages and extravillous trophoblast apoptosis. Mol. Hum. Reprod. 2012, 18, 535–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, S.A.; Green, E.S.; Care, A.S.; Moldenhauer, L.M.; Prins, J.R.; Hull, M.L.; Barry, S.C.; Dekker, G. Therapeutic potential of regulatory T Cells in preeclampsia-opportunities and challenges. Front. Immunol. 2019, 10, 478. [Google Scholar] [CrossRef] [PubMed]
- Alijotas-Reig, J.; Esteve-Valverde, E.; Ferrer-Oliveras, R.; Llurba, E.; Gris, J.M. Tumor necrosis factor-alpha and pregnancy: Focus on biologics. An updated and comprehensive review. Clin. Rev. Allergy Immunol. 2017, 53, 40–53. [Google Scholar] [CrossRef]
- Scholz, C.; Toth, B.; Santoso, L.; Kuhn, C.; Franz, M.; Mayr, D.; Jeschke, U.; Friese, K.; Schiessl, B. Distribution and maturity of dendritic cells in diseases of insufficient placentation. Am. J. Reprod. Immunol. 2008, 60, 238–245. [Google Scholar] [CrossRef]
- Minas, V.; Mylonas, I.; Schiessl, B.; Mayr, D.; Schulze, S.; Friese, K.; Jeschke, U.; Makrigiannakis, A. Expression of the blood-group-related antigens Sialyl Lewis a, Sialyl Lewis x and Lewis y in term placentas of normal, preeclampsia, IUGR- and HELLP-complicated pregnancies. Histochem. Cell Biol. 2007, 128, 55–63. [Google Scholar] [CrossRef]
- Chakraborty, D.; Cui, W.; Rosario, G.X.; Scott, R.L.; Dhakal, P.; Renaud, S.J.; Tachibana, M.; Rumi, M.A.; Mason, C.W.; Krieg, A.J.; et al. HIF-KDM3A-MMP12 regulatory circuit ensures trophoblast plasticity and placental adaptations to hypoxia. Proc. Natl. Acad. Sci. USA 2016, 113, E7212–E7221. [Google Scholar] [CrossRef] [Green Version]
- Pozharny, Y.; Lambertini, L.; Clunie, G.; Ferrara, L.; Lee, M.J. Epigenetics in women’s health care. Mt. Sinai J. Med. 2010, 77, 225–235. [Google Scholar] [CrossRef]
- Zhu, H.; Kong, L. LncRNA CRNDE regulates trophoblast cell proliferation, invasion, and migration via modulating miR-1277. Am. J. Transl. Res. 2019, 11, 5905–5918. [Google Scholar] [PubMed]
- Alpoim, P.N.; Godoi, L.C.; Pinheiro, M.B.; Freitas, L.G.; Carvalho, M.D.G.; Dusse, L.M. The unexpected beneficial role of smoking in preeclampsia. Clin. Chim. Acta 2016, 459, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Ye, Y.; Zhang, J.; Ruan, C.C.; Gao, P.J. Immune imbalance is associated with the development of preeclampsia. Medicine 2019, 98, e15080. [Google Scholar] [CrossRef] [PubMed]
- Eddy, A.C.; Chapman, H.; George, E.M. Acute hypoxia and chronic ischemia induce differential total changes in placental epigenetic modifications. Reprod. Sci. 2019, 26, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Apicella, C.; Ruano, C.S.M.; Mehats, C.; Miralles, F.; Vaiman, D. The role of epigenetics in placental development and the etiology of preeclampsia. Int. J. Mol. Sci. 2019, 20, 2837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Z.; Zhang, L.; Tang, Q.; Xu, Y.; Liu, S.; Li, H. Circulating levels of IFN-gamma, IL-1, IL-17 and IL-22 in pre-eclampsia: A systematic review and meta-analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 248, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, B.; Schleußner, E. Die Plazenta; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Magee, L.A.; Rey, E.; Asztalos, E.; Hutton, E.; Singer, J.; Helewa, M.; Lee, T.; Logan, A.G.; Ganzevoort, W.; Welch, R.; et al. Management of non-severe pregnancy hypertension—A summary of the CHIPS Trial (Control of Hypertension in Pregnancy Study) research publications. Pregnancy Hypertens 2019, 18, 156–162. [Google Scholar] [CrossRef]
- Nguyen, T.P.H.; Patrick, C.J.; Parry, L.J.; Familari, M. Using proteomics to advance the search for potential biomarkers for preeclampsia: A systematic review and meta-analysis. PLoS ONE 2019, 14, e0214671. [Google Scholar] [CrossRef] [Green Version]
- Izumi, Y.; Matsuo, K.; Namatame, H. Structural analysis of lysine-4 methylated histone H3 proteins using synchrotron radiation circular dichroism spectroscopy. Chirality 2018, 30, 536–540. [Google Scholar] [CrossRef]
- Park, Y.; Lee, H.J.; Jung, Y.J.; Kwon, H.Y.; Kim, H.; Lee, J.; Kim, Y.H.; Kim, H.O.; Maeng, Y.S.; Kwon, J.Y. CD133+/C-kit+Lin(-) endothelial progenitor cells in fetal circulation demonstrate impaired differentiation potency in severe preeclampsia. Pregnancy Hypertens 2019, 15, 146–153. [Google Scholar] [CrossRef]
- Ushida, T.; Macdonald-Goodfellow, S.K.; Quadri, A.; Tse, M.Y.; Winn, L.M.; Pang, S.C.; Adams, M.A.; Kotani, T.; Kikkawa, F.; Graham, C.H. Persistence of risk factors associated with maternal cardiovascular disease following aberrant inflammation in rat pregnancy. Biol. Reprod. 2017, 97, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Meister, S.; Hahn, L.; Beyer, S.; Kuhn, C.; Jegen, M.; von Schonfeldt, V.; Corradini, S.; Schulz, C.; Kolben, T.M.; Hester, A.; et al. Epigenetic modification via H3K4me3 and H3K9ac in human placenta is reduced in preeclampsia. J. Reprod. Immunol. 2021, 145, 103287. [Google Scholar] [CrossRef] [PubMed]
- Kamrani, A.; Alipourfard, I.; Ahmadi-Khiavi, H.; Yousefi, M.; Rostamzadeh, D.; Izadi, M.; Ahmadi, M. The role of epigenetic changes in preeclampsia. Biofactors 2019, 45, 712–724. [Google Scholar] [CrossRef] [PubMed]
- Patsouras, M.D.; Vlachoyiannopoulos, P.G. Evidence of epigenetic alterations in thrombosis and coagulation: A systematic review. J. Autoimmun. 2019, 104, 102347. [Google Scholar] [CrossRef]
- Rahat, B.; Najar, R.A.; Hamid, A.; Bagga, R.; Kaur, J. The role of aberrant methylation of trophoblastic stem cell origin in the pathogenesis and diagnosis of placental disorders. Prenat. Diagn. 2017, 37, 133–143. [Google Scholar] [CrossRef]
- Bojic-Trbojevic, Z.; Jovanovic Krivokuca, M.; Vilotic, A.; Kolundzic, N.; Stefanoska, I.; Zetterberg, F.; Nilsson, U.J.; Leffler, H.; Vicovac, L. Human trophoblast requires galectin-3 for cell migration and invasion. Sci. Rep. 2019, 9, 2136. [Google Scholar] [CrossRef]
- Fischer, I.; Redel, S.; Hofmann, S.; Kuhn, C.; Friese, K.; Walzel, H.; Jeschke, U. Stimulation of syncytium formation in vitro in human trophoblast cells by galectin-1. Placenta 2010, 31, 825–832. [Google Scholar] [CrossRef]
- Hutter, S.; Martin, N.; von Schonfeldt, V.; Messner, J.; Kuhn, C.; Hofmann, S.; Andergassen, U.; Knabl, J.; Jeschke, U. Galectin 2 (gal-2) expression is downregulated on protein and mRNA level in placentas of preeclamptic (PE) patients. Placenta 2015, 36, 438–445. [Google Scholar] [CrossRef]
- Loser, K.; Sturm, A.; Voskort, M.; Kupas, V.; Balkow, S.; Auriemma, M.; Sternemann, C.; Dignass, A.U.; Luger, T.A.; Beissert, S. Galectin-2 suppresses contact allergy by inducing apoptosis in activated CD8+ T cells. J Immunol. 2009, 182, 5419–5429. [Google Scholar] [CrossRef] [Green Version]
- Nio-Kobayashi, J.; Takahashi-Iwanaga, H.; Iwanaga, T. Immunohistochemical localization of six galectin subtypes in the mouse digestive tract. J. Histochem. Cytochem. 2009, 57, 41–50. [Google Scholar] [CrossRef]
- Hutter, S.; Knabl, J.; Andergassen, U.; Hofmann, S.; Kuhn, C.; Mahner, S.; Arck, P.; Jeschke, U. Placental Expression Patterns of Galectin-1, Galectin-2, Galectin-3 and Galectin-13 in Cases of Intrauterine Growth Restriction (IUGR). Int. J. Mol. Sci. 2016, 17, 523. [Google Scholar] [CrossRef] [PubMed]
- Unverdorben, L.; Haufe, T.; Santoso, L.; Hofmann, S.; Jeschke, U.; Hutter, S. Prototype and chimera-type galectins in placentas with spontaneous and recurrent miscarriages. Int. J. Mol. Sci. 2016, 17, 644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hokama, A.; Mizoguchi, E.; Mizoguchi, A. Roles of galectins in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 5133–5137. [Google Scholar] [CrossRef] [PubMed]
- Paclik, D.; Berndt, U.; Guzy, C.; Dankof, A.; Danese, S.; Holzloehner, P.; Rosewicz, S.; Wiedenmann, B.; Wittig, B.M.; Dignass, A.U.; et al. Galectin-2 induces apoptosis of lamina propria T lymphocytes and ameliorates acute and chronic experimental colitis in mice. J. Mol. Med. 2008, 86, 1395–1406. [Google Scholar] [CrossRef]
- Charkiewicz, K.; Goscik, J.; Raba, G.; Laudanski, P. Syndecan 4, galectin 2, and death receptor 3 (DR3) as novel proteins in pathophysiology of preeclampsia. J. Matern. Fetal Neonatal Med. 2021, 34, 2965–2970. [Google Scholar] [CrossRef]
- Hutter, S.; Morales-Prieto, D.M.; Andergassen, U.; Tschakert, L.; Kuhn, C.; Hofmann, S.; Markert, U.R.; Jeschke, U. Gal-1 silenced trophoblast tumor cells (BeWo) show decreased syncytium formation and different miRNA production compared to non-target silenced BeWo cells. Cell Adh. Migr. 2016, 10, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Gillenwater, A.; Xu, X.C.; Estrov, Y.; Sacks, P.G.; Lotan, D.; Lotan, R. Modulation of Galectin-1 Content in Human Head and Neck Squamous Carcinoma Cells by Sodium Butyrate. Int. J. Cancer 1998, 75, 217–224. [Google Scholar] [CrossRef]
- Jensen, H.; Hagemann-Jensen, M.; Lauridsen, F.; Skov, S. Regulation of NKG2D-ligand cell surface expression by intracellular calcium after HDAC-inhibitor treatment. Mol. Immunol. 2013, 53, 255–264. [Google Scholar] [CrossRef]
- Kultima, K.; Nystrom, A.M.; Scholz, B.; Gustafson, A.L.; Dencker, L.; Stigson, M. Valproic acid teratogenicity: A toxicogenomics approach. Environ. Health Perspect. 2004, 112, 1225–1235. [Google Scholar] [CrossRef]
- Maier, M.S.; Legare, M.E.; Hanneman, W.H. The aryl hydrocarbon receptor agonist 3,3’,4,4’,5-pentachlorobiphenyl induces distinct patterns of gene expression between hepatoma and glioma cells: Chromatin remodeling as a mechanism for selective effects. Neurotoxicology 2007, 28, 594–612. [Google Scholar] [CrossRef]
- Ohkouchi, C.; Kumamoto, K.; Saito, M.; Ishigame, T.; Suzuki, S.I.; Takenoshita, S.; Harris, C.C. ING2, a tumor associated gene, enhances PAI1 and HSPA1A expression with HDAC1 and mSin3A through the PHD domain and Cterminal. Mol. Med. Rep. 2017, 16, 7367–7374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, H.S.; Lee, C.; Lee, K.S.; Park, E.J.; Seong, R.H.; Hong, S.; Jeon, S.H. Twist2 regulates CD7 expression and galectin-1-induced apoptosis in mature T-cells. Mol. Cells 2009, 28, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Hattori, N.; Imao, Y.; Nishino, K.; Hattori, N.; Ohgane, J.; Yagi, S.; Tanaka, S.; Shiota, K. Epigenetic regulation of Nanog gene in embryonic stem and trophoblast stem cells. Genes Cells 2007, 12, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Fan, W.; Lei, J.; Zhou, Y.; Xu, H.; Kapoor, I.; Zhu, G.; Wang, J. Galectin-1 attenuates cardiomyocyte hypertrophy through splice-variant specific modulation of CaV1.2 calcium channel. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 218–229. [Google Scholar] [CrossRef]
- Seetharaman, J.; Kanigsberg, A.; Slaaby, R.; Leffler, H.; Barondes, S.H.; Rini, J.M. X-ray crystal structure of the human galectin-3 carbohydrate recognition domain at 2.1-A resolution. J. Biol. Chem. 1998, 273, 13047–13052. [Google Scholar] [CrossRef] [Green Version]
- Sturm, A.; Lensch, M.; Andre, S.; Kaltner, H.; Wiedenmann, B.; Rosewicz, S.; Dignass, A.U.; Gabius, H.J. Human galectin-2: Novel inducer of T cell apoptosis with distinct profile of caspase activation. J. Immunol. 2004, 173, 3825–3837. [Google Scholar] [CrossRef] [Green Version]
- Dvorankova, B.; Lacina, L.; Smetana, K.; Lensch, M., Jr.; Manning, J.C.; Andre, S.; Gabius, H.J. Human galectin-2: Nuclear presence in vitro and its modulation by quiescence/stress factors. Histol. Histopathol. 2008, 23, 167–178. [Google Scholar]
- Drwal, E.; Rak, A.; Gregoraszczuk, E. Co-culture of JEG-3, BeWo and syncBeWo cell lines with adrenal H295R cell line: An alternative model for examining endocrine and metabolic properties of the fetoplacental unit. Cytotechnology 2018, 70, 285–297. [Google Scholar] [CrossRef]
- Wiest, I.; Seliger, C.; Walzel, H.; Friese, K.; Jeschke, U. Induction of apoptosis in human breast cancer and trophoblast tumor cells by galectin-1. Anticancer Res. 2005, 25, 1575–1580. [Google Scholar]
- Rogenhofer, N.; Ochsenkuhn, R.; von Schonfeldt, V.; Assef, R.B.; Thaler, C.J. Antitrophoblast antibodies are associated with recurrent miscarriages. Fertil Steril. 2012, 97, 361–366. [Google Scholar] [CrossRef]
- Issman, L.; Brenner, B.; Talmon, Y.; Aharon, A. Cryogenic transmission electron microscopy nanostructural study of shed microparticles. PLoS ONE 2013, 8, e83680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudoh, A.; Miyakawa, K.; Matsunaga, S.; Matsushima, Y.; Kosugi, I.; Kimura, H.; Hayakawa, S.; Sawasaki, T.; Ryo, A. H11/HSPB8 Restricts HIV-2 Vpx to Restore the Anti-Viral Activity of SAMHD1. Front. Microbiol. 2016, 7, 883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knofler, M.; Haider, S.; Saleh, L.; Pollheimer, J.; Gamage, T.; James, J. Human placenta and trophoblast development: Key molecular mechanisms and model systems. Cell. Mol. Life Sci. 2019, 76, 3479–3496. [Google Scholar] [CrossRef] [Green Version]
- Sheridan, M.A.; Fernando, R.C.; Gardner, L.; Hollinshead, M.S.; Burton, G.J.; Moffett, A.; Turco, M.Y. Establishment and differentiation of long-term trophoblast organoid cultures from the human placenta. Nat. Protoc. 2020, 15, 3441–3463. [Google Scholar] [CrossRef] [PubMed]
- Kakkassery, V.; Wirtz, C.; Schargus, M.; Grisanti, S.; Tura, A.; Ranjbar, M.; Dick, H.B.; Reinehr, S.; Joachim, S.C. Epidermal Growth Factor Is Increased in Conjunctival Malignant Melanoma. In Vivo 2021, 35, 3603–3612. [Google Scholar] [CrossRef]
- Coutifaris, C.; Kao, L.C.; Sehdev, H.M.; Chin, U.; Babalola, G.O.; Blaschuk, O.W.; Strauss, J.F. E-cadherin expression during the differentiation of human trophoblasts. Development 1991, 113, 767–777. [Google Scholar] [CrossRef]
- Gauster, M.; Siwetz, M.; Orendi, K.; Moser, G.; Desoye, G.; Huppertz, B. Caspases rather than calpains mediate remodelling of the fodrin skeleton during human placental trophoblast fusion. Cell Death Differ. 2010, 17, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Shankar, K.; Kang, P.; Zhong, Y.; Borengasser, S.J.; Wingfield, C.; Saben, J.; Gomez-Acevedo, H.; Thakali, K.M. Transcriptomic and epigenomic landscapes during cell fusion in BeWo trophoblast cells. Placenta 2015, 36, 1342–1351. [Google Scholar] [CrossRef]
- Bakheit, K.H.; Bayoumi, N.K.; Eltom, A.M.; Elbashir, M.I.; Adam, I. Cytokines profiles in Sudanese women with preeclampsia. Hypertens Pregnancy 2009, 28, 224–229. [Google Scholar] [CrossRef]
- He, P.; Chen, Z.; Sun, Q.; Li, Y.; Gu, H.; Ni, X. Reduced expression of 11beta-hydroxysteroid dehydrogenase type 2 in preeclamptic placentas is associated with decreased PPARgamma but increased PPARalpha expression. Endocrinology 2014, 155, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Okae, H.; Toh, H.; Sato, T.; Hiura, H.; Takahashi, S.; Shirane, K.; Kabayama, Y.; Suyama, M.; Sasaki, H.; Arima, T. Derivation of human trophoblast stem cells. Cell Stem Cell 2018, 22, 50–63.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pattillo, R.A.; Gey, G.O.; Delfs, E.; Mattingly, R.F. Human hormone production in vitro. Science 1968, 159, 1467–1469. [Google Scholar] [CrossRef] [PubMed]
- Rahat, B.; Thakur, S.; Bagga, R.; Kaur, J. Epigenetic regulation of STAT5A and its role as fetal DNA epigenetic marker during placental development and dysfunction. Placenta 2016, 44, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, U.; Schiessl, B.; Mylonas, I.; Kunze, S.; Kuhn, C.; Schulze, S.; Friese, K.; Mayr, D. Expression of the proliferation marker Ki-67 and of p53 tumor protein in trophoblastic tissue of preeclamptic, HELLP, and intrauterine growth-restricted pregnancies. Int. J. Gynecol. Pathol. 2006, 25, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Tache, V.; Ciric, A.; Moretto-Zita, M.; Li, Y.; Peng, J.; Maltepe, E.; Milstone, D.S.; Parast, M.M. Hypoxia and trophoblast differentiation: A key role for PPARgamma. Stem Cells Dev. 2013, 22, 2815–2824. [Google Scholar] [CrossRef] [Green Version]
- Ruebner, M.; Langbein, M.; Strissel, P.L.; Henke, C.; Schmidt, D.; Goecke, T.W.; Faschingbauer, F.; Schild, R.L.; Beckmann, M.W.; Strick, R. Regulation of the human endogenous retroviral Syncytin-1 and cell-cell fusion by the nuclear hormone receptors PPARgamma/RXRalpha in placentogenesis. J. Cell Biochem. 2012, 113, 2383–2396. [Google Scholar] [CrossRef] [PubMed]
- Kudo, Y.; Boyd, C.A.R.; Sargent, I.L.; Redman, C.W.G. Hypoxia alters expression and function of syncytin and its receptor during trophoblast cell fusion of human placental BeWo cells: Implications for impaired trophoblast syncytialisation in pre-eclampsia. Biochim. Biophys. Acta Mol. Basis Dis. 2003, 1638, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zang, C.; Rosenfeld, J.A.; Schones, D.E.; Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.-Y.; Peng, W.; Zhang, M.Q. Combinatorial patterns of histone acetylations and methylations in the human genome. Nat. Genet. 2008, 40, 897–903. [Google Scholar] [CrossRef] [Green Version]
- Yavartanoo, M.; Choi, J.K. ENCODE: A sourcebook of epigenomes and chromatin language. Genom. Inform. 2013, 11, 2. [Google Scholar] [CrossRef]
- Toudic, C.; Vargas, A.; Xiao, Y.; St-Pierre, G.; Bannert, N.; Lafond, J.; Rassart, E.; Sato, S.; Barbeau, B. Galectin-1 interacts with the human endogenous retroviral envelope protein syncytin-2 and potentiates trophoblast fusion in humans. FASEB J. 2019, 33, 12873–12887. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, W.; Liu, C.; Wang, W.; Li, W.; Shu, Q.; Chen, Z.J.; Sun, K. Critical role of histone acetylation by p300 in human placental 11beta-HSD2 expression. J. Clin. Endocrinol. Metab. 2013, 98, E1189–E1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knabl, J.; Hiden, U.; Huttenbrenner, R.; Riedel, C.; Hutter, S.; Kirn, V.; Gunthner-Biller, M.; Desoye, G.; Kainer, F.; Jeschke, U. GDM Alters expression of placental estrogen receptor alpha in a cell type and gender-specific manner. Reprod. Sci. 2015, 22, 1488–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Antibody | Species Isotyp | Company | Dilution |
|---|---|---|---|
| H3K4me3 | Rabbit IgG polyclonal | Abcam ab8580 | 1:100 |
| H3K9ac | Rabbit IgG monoclonal, Y28 | Abcam ab3129 | 1:200 |
| Goat-Anti-Mouse Alexa Fluor 488 | Goat IgG, polyclonal | Dianova 115-226-062 | 1:100 |
| Goat-Anti-Rabbit Cy3 | Goat IgG, polyclonal | Dianova 111-165-144 | 1:500 |
| E-Cadherin | Mouse IgG1, monoclonal HECD-1 | Calbiochem | 1:50 |
| β-Catenin | Rabbit IgG, polyclonal | Diagnostic BioSystems | 1:50 |
| Gal-2 in All Placentas | Gal-2 Only in Controls | Gal-2 Only in PE | |||||
|---|---|---|---|---|---|---|---|
| Syn | Dec | Syn | Dec | Syn | Dec | ||
| H3K4me3 | Syn | 0.486 * 0.012 | 0.450 * 0.021 | 0.390 0.188 | 0.366 0.219 | 0.247 0.416 | 0.285 0.345 |
| Dec | 0.402 * 0.042 | 0.286 0.156 | 0.193 0.527 | 0.234 0.441 | 0.322 0.283 | 0.126 0.682 | |
| H3K9ac | Syn | 0.463 * 0.017 | 0.368 0.065 | 0.019 0.951 | 0.020 0.948 | 0.556 * 0.048 | 0.456 0.117 |
| Dec | 0.559 ** 0.003 | 0.545 ** 0.004 | 0.197 0.519 | 0.151 0.621 | 0.759 ** 0.003 | 0.752 ** 0.003 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hahn, L.; Meister, S.; Mannewitz, M.; Beyer, S.; Corradini, S.; Hasbargen, U.; Mahner, S.; Jeschke, U.; Kolben, T.; Burges, A. Gal-2 Increases H3K4me3 and H3K9ac in Trophoblasts and Preeclampsia. Biomolecules 2022, 12, 707. https://doi.org/10.3390/biom12050707
Hahn L, Meister S, Mannewitz M, Beyer S, Corradini S, Hasbargen U, Mahner S, Jeschke U, Kolben T, Burges A. Gal-2 Increases H3K4me3 and H3K9ac in Trophoblasts and Preeclampsia. Biomolecules. 2022; 12(5):707. https://doi.org/10.3390/biom12050707
Chicago/Turabian StyleHahn, Laura, Sarah Meister, Mareike Mannewitz, Susanne Beyer, Stefanie Corradini, Uwe Hasbargen, Sven Mahner, Udo Jeschke, Thomas Kolben, and Alexander Burges. 2022. "Gal-2 Increases H3K4me3 and H3K9ac in Trophoblasts and Preeclampsia" Biomolecules 12, no. 5: 707. https://doi.org/10.3390/biom12050707
APA StyleHahn, L., Meister, S., Mannewitz, M., Beyer, S., Corradini, S., Hasbargen, U., Mahner, S., Jeschke, U., Kolben, T., & Burges, A. (2022). Gal-2 Increases H3K4me3 and H3K9ac in Trophoblasts and Preeclampsia. Biomolecules, 12(5), 707. https://doi.org/10.3390/biom12050707