
Targeting Nrf2 with Probiotics and Postbiotics in the Treatment of Periodontitis


Abstract
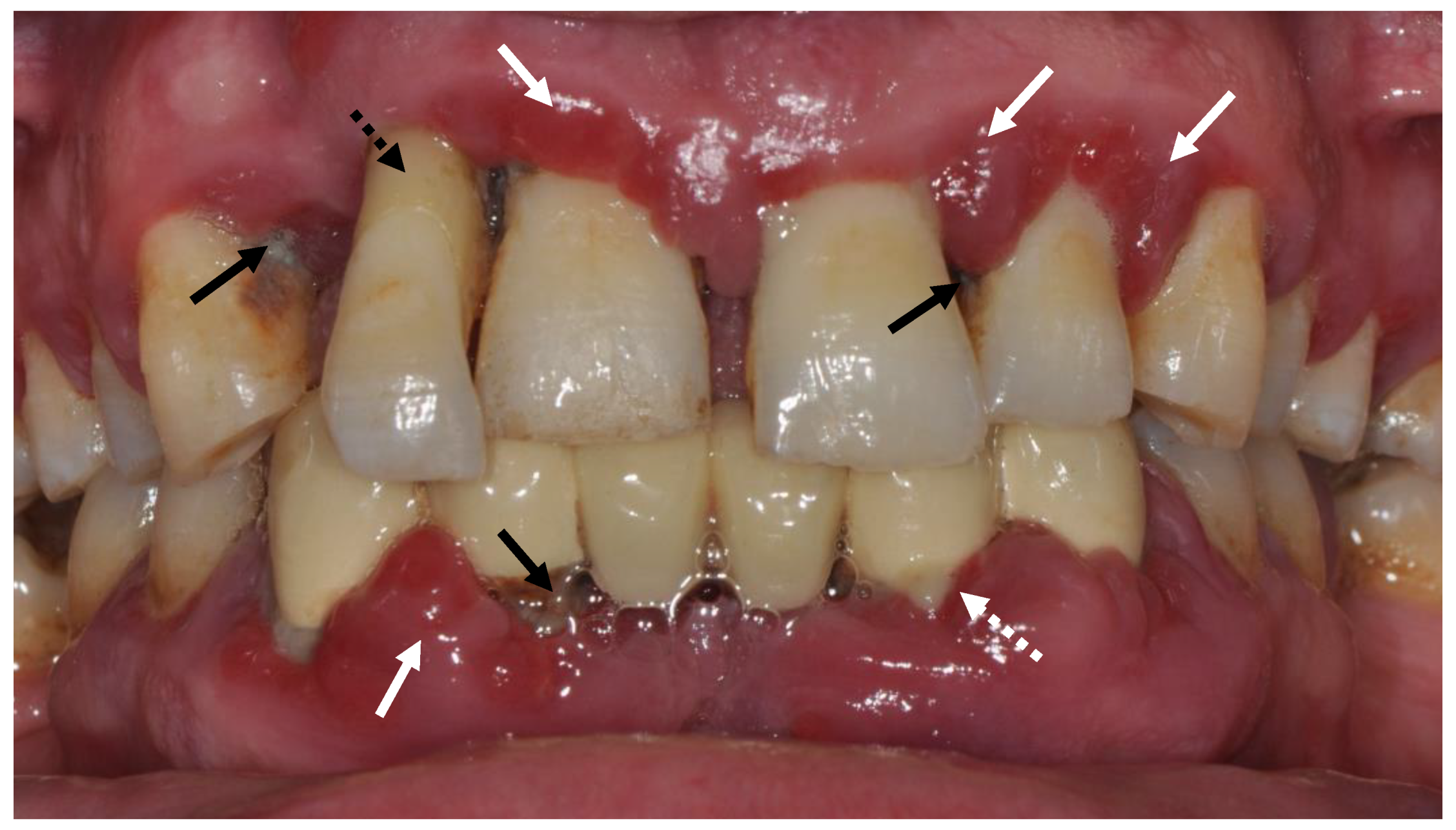
:1. Periodontal Health and Disease
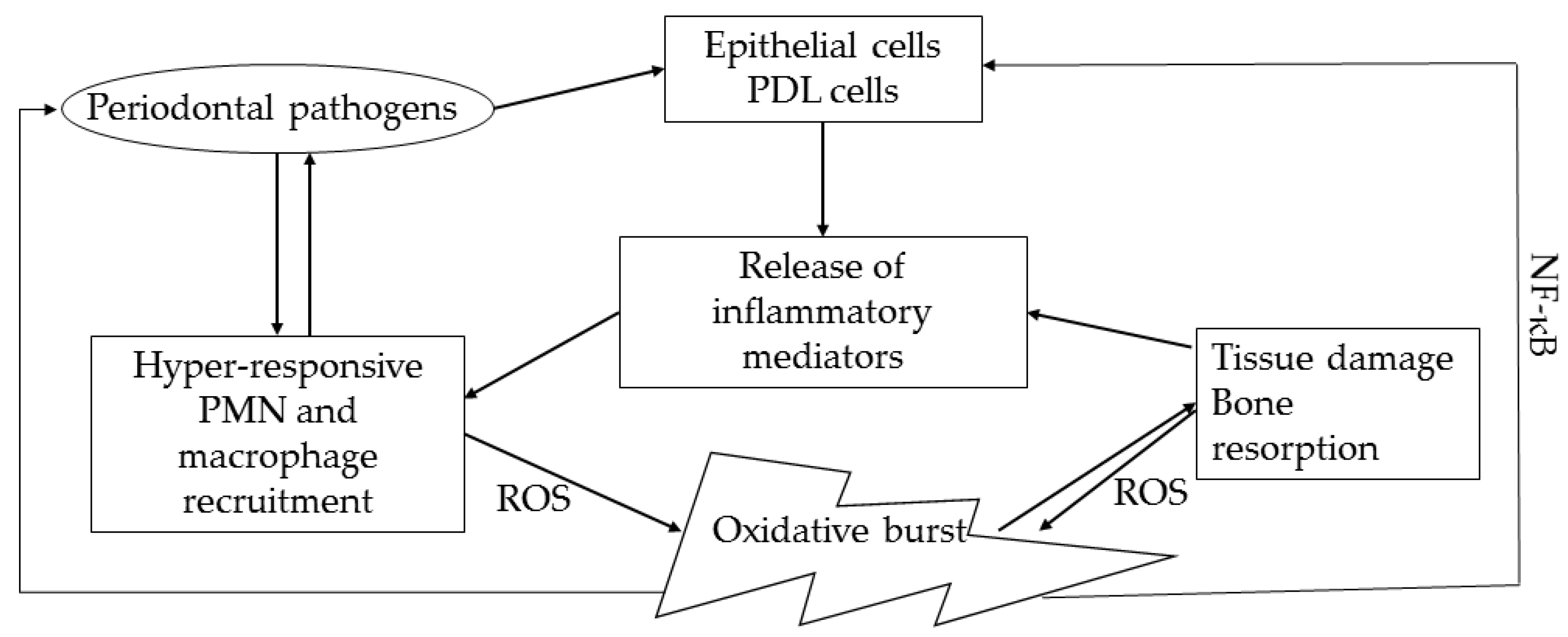
2. Oxidative Stress in Pathogenesis of Periodontal Disease
3. Anti-Oxidative Mechanisms in Periodontal Tissues
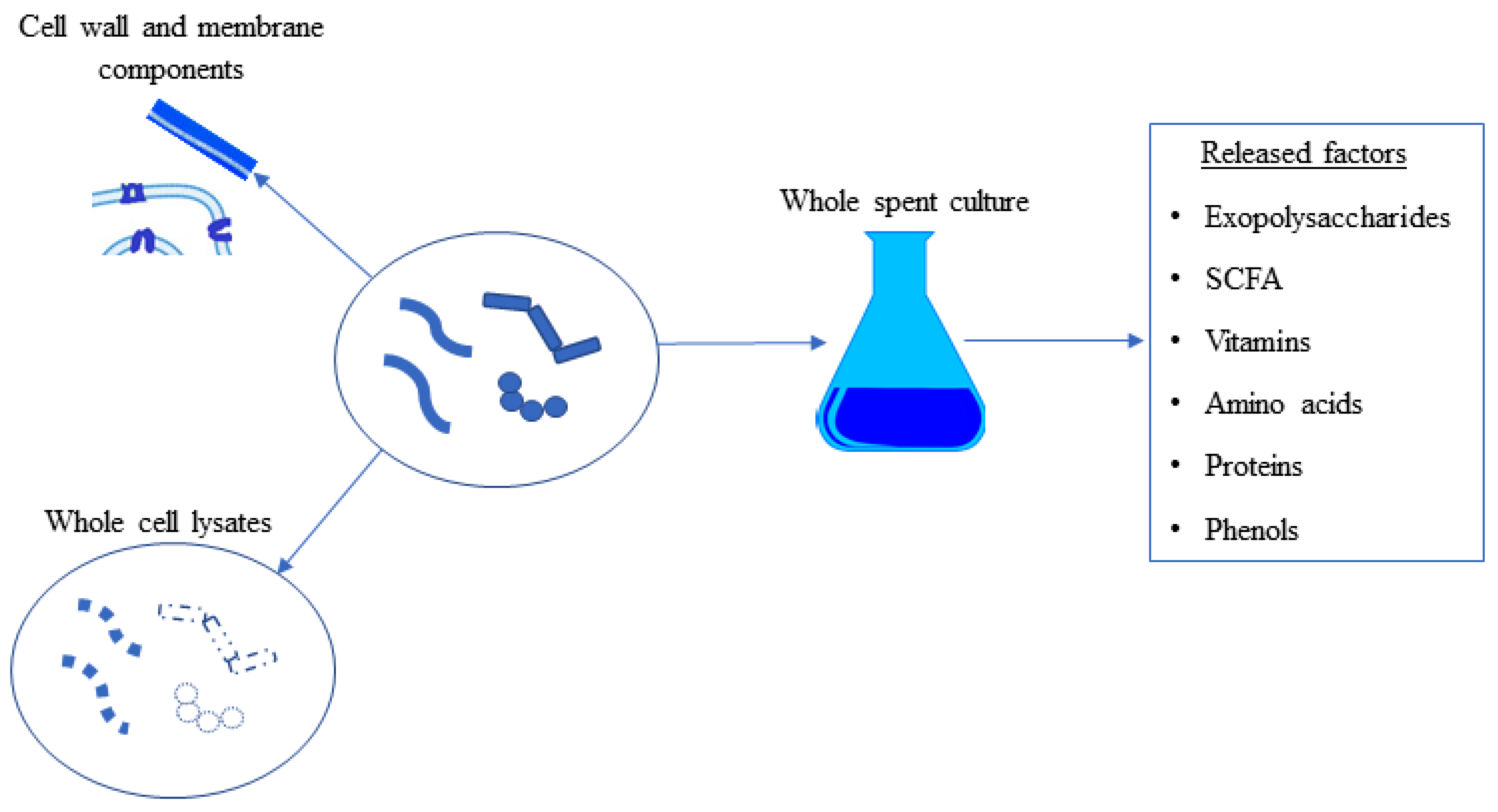
4. Probiotics and Postbiotics
5. Antioxidant Abilities of Probiotics and Postbiotics
6. Probiotics and Nrf2 Activation
7. Postbiotics and Nrf2 Activation
7.1. Exopolysaccharides (EPS)
7.2. Short-Chain Fatty Acids (SCFA)
7.3. Carotenoids
7.4. Bioconverted Metabolites as Postbiotics
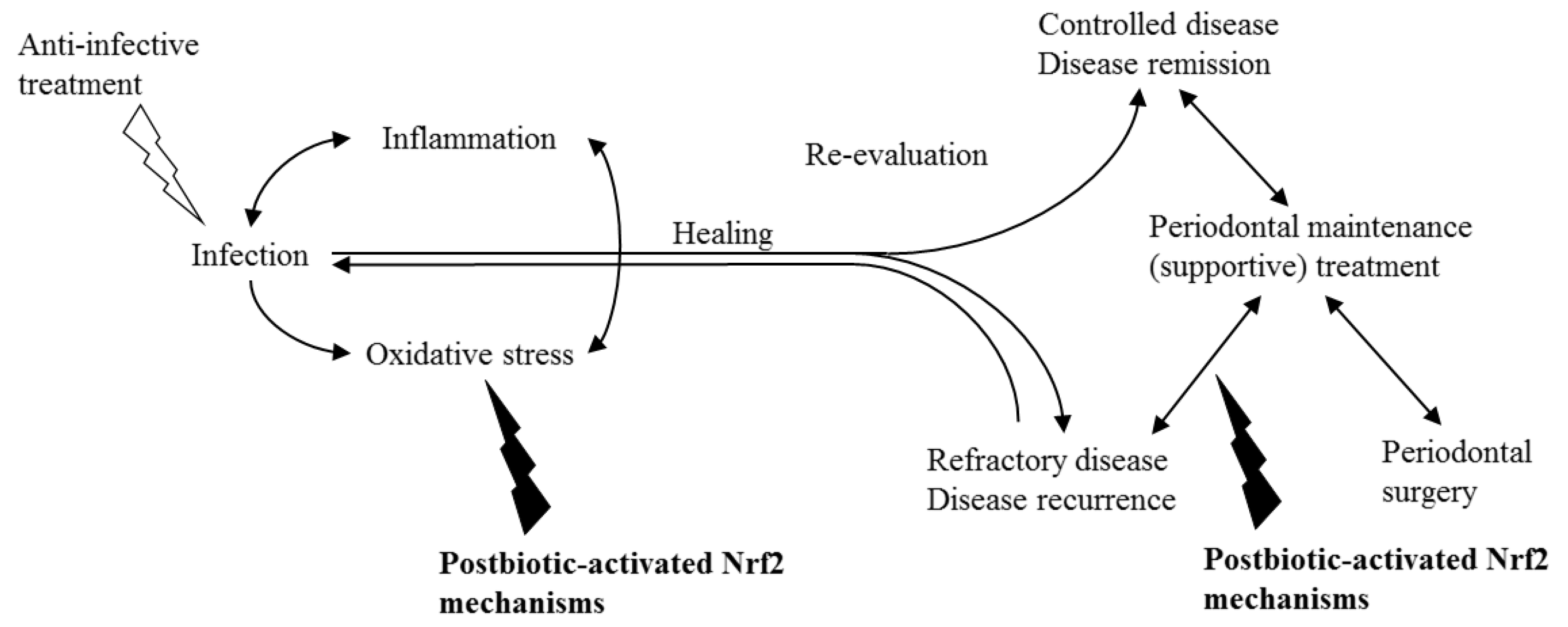
8. Postbiotics as Nrf2 Activators in Periodontitis: Scientific Evidence
9. Postbiotics as Nrf2 Activators and Their Potential Use as Antioxidants in Periodontitis—Future Perspective
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chapple, I.L.C.; Mealey, B.L.; Van Dyke, T.E.; Bartold, P.M.; Dommisch, H.; Eickholz, P.; Geisinger, M.L.; Genco, R.J.; Glogauer, M.; Goldstein, M.; et al. Periodontal Health and Gingival Diseases and Conditions on an Intact and a Reduced Periodontium: Consensus Report of Workgroup 1 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Periodontol. 2018, 89, S74–S84. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.; Abusleme, L.; Bravo, D.; Dutzan, N.; Garcia-Sesnich, J.; Vernal, R.; Hernández, M.; Gamonal, J. Host Response Mechanisms in Periodontal Diseases. J. Appl. Oral Sci. 2015, 23, 329–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dyke, T.E.; Sheilesh, D. Risk Factors for Periodontitis. J. Int. Acad. Periodontol. 2005, 7, 3–7. [Google Scholar] [PubMed]
- Cekici, A.; Kantarci, A.; Hasturk, H.; Van Dyke, T.E. Inflammatory and Immune Pathways in the Pathogenesis of Periodontal Disease. Periodontol. 2000 2014, 64, 57–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-García, M.; Hernández-Lemus, E. Periodontal Inflammation and Systemic Diseases: An Overview. Front. Physiol. 2021, 12, 709438. [Google Scholar] [CrossRef]
- Landzberg, M.; Doering, H.; Aboodi, G.M.; Tenenbaum, H.C.; Glogauer, M. Quantifying Oral Inflammatory Load: Oral Neutrophil Counts in Periodontal Health and Disease. J. Periodontal Res. 2015, 50, 330–336. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Chavakis, T.; Hajishengallis, E.; Lambris, J.D. Neutrophil Homeostasis and Inflammation: Novel Paradigms from Studying Periodontitis. J. Leukoc. Biol. 2015, 98, 539–548. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Korostoff, J.M. Revisiting the Page & Schroeder Model: The Good, the Bad and the Unknowns in the Periodontal Host Response 40 Years Later. Periodontol. 2000 2017, 75, 116–151. [Google Scholar] [CrossRef]
- Hajishengallis, G. Immunomicrobial Pathogenesis of Periodontitis: Keystones, Pathobionts, and Host Response. Trends Immunol. 2014, 35, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Bagan, J.; Saez, G.; Tormos, C.; Gavalda, C.; Sanchis, J.M.; Bagan, L.; Scully, C. Oxidative stress and recurrent aphthous stomatitis. Clin. Oral Investig. 2014, 18, 1919–1923. [Google Scholar] [CrossRef]
- Tvarijonaviciute, A.; Aznar-Cayuela, C.; Rubio, C.P.; Ceron, J.J.; López-Jornet, P. Evaluation of salivary oxidate stress biomarkers, nitric oxide and C-reactive protein in patients with oral lichen planus and burning mouth syndrome. J. Oral Pathol. Med. 2017, 46, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Sardaro, N.; Della Vella, F.; Incalza, M.A.; Di Stasio, D.; Lucchese, A.; Contaldo, M.; Laudadio, C.; Petruzzi, M. Oxidative stress and oral mucosal diseases: An overview. In Vivo 2019, 33, 289–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, R.B.; Carnelio, S.; Shenoy, R.P.; Gyawali, P.; Mukherjee, M. Oxidative stress and antioxidant defense in oral lichen planus and oral lichenoid reaction. Scand. J. Clin. Lab. 2010, 70, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cai, W.; Zhao, S.; Shi, L.; Chen, Y.; Li, X.; Sun, X.; Mao, Y.; He, B.; Hou, Y.; et al. Oxidative Stress-Related Biomarkers in Saliva and Gingival Crevicular Fluid Associated with Chronic Periodontitis: A Systematic Review and Meta-Analysis. J. Clin. Periodontol. 2019, 46, 608–622. [Google Scholar] [CrossRef] [PubMed]
- Sczepanik, S.F.C.; Grossi, M.L.; Casati, M.; Goldberg, M.; Glogauer, M.; Fine, N.; Tenenbaum, H.C. Periodontitis Is an Inflammatory Disease of Oxidative Stress: We Should Treat It That Way. Periodontol. 2000 2020, 84, 45–68. [Google Scholar] [CrossRef]
- Dias, I.H.; Matthews, J.B.; Chapple, I.L.; Wright, H.J.; Dunston, C.R.; Griffiths, H.R. Activation of the neutrophil respiratory burst by plasma from periodontitis patients is mediated by pro-inflammatory cytokines. J. Clin. Periodontol. 2011, 38, 1–7. [Google Scholar] [CrossRef]
- Chapple, I.L.C.; Matthews, J.B. The Role of Reactive Oxygen and Antioxidant Species in Periodontal Tissue Destruction. Periodontol. 2000 2007, 43, 160–232. [Google Scholar] [CrossRef]
- Ling, M.R.; Chapple, I.L.C.; Matthews, J.B. Neutrophil superoxide release and plasma C-reactive protein levels pre- and post-periodontal therapy. J. Clin. Periodontol. 2016, 43, 652–658. [Google Scholar] [CrossRef]
- Katsuragi, H.; Ohtake, M.; Kurasawa, I.; Saito, K. Intracellular production and extracellular release of oxygen radicals by PMNs and oxidative stress on PMNs during phagocytosis of periodontopathic bacteria. Odontology 2003, 91, 13–18. [Google Scholar] [CrossRef]
- Chapple, I.L.C. Role of Free Radicals and Antioxidants in the Pathogenesis of the Inflammatory Periodontal Diseases. Clin. Mol. Pathol. 1996, 49, M247. [Google Scholar] [CrossRef] [Green Version]
- AItman, L.C.; Baker, C.; Fleckman, P.; Luchtel, D.; Oda, D. Neutrophil-Mediated Damage to Human Gingival Epithelial Cells. J. Periodontal Res. 1992, 27, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Choi, Y.G.; Baik, J.Y.; Han, S.Y.; Jeong, D.W.; Bae, Y.S.; Kim, N.; Lee, S.Y. A Crucial Role for Reactive Oxygen Species in RANKL-Induced Osteoclast Differentiation. Blood 2005, 106, 852–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, I.H.K.; Chapple, I.L.C.; Milward, M.; Grant, M.M.; Hill, E.; Brown, J.; Griffiths, H.R. Sulforaphane Restores Cellular Glutathione Levels and Reduces Chronic Periodontitis Neutrophil Hyperactivity In Vitro. PLoS ONE 2013, 8, e66407. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, L.; Mesgarzadeh, J.; Xu, I.; Powers, E.T.; Wiseman, R.L.; Bollong, M.J. Defining the functional targets of Cap ‘n’collar transcription factors NRF1, NRF2, and NRF3. Antioxidants 2020, 9, 1025. [Google Scholar] [CrossRef]
- Chapple, I.L.C.; Brock, G.; Eftimiadi, C.; Matthews, J.B. Glutathione in Gingival Crevicular Fluid and Its Relation to Local Antioxidant Capacity in Periodontal Health and Disease. Mol. Pathol. 2002, 55, 367. [Google Scholar] [CrossRef] [Green Version]
- Brock, G.R.; Butterworth, C.J.; Matthews, J.B.; Chapple, I.L.C. Local and Systemic Total Antioxidant Capacity in Periodontitis and Health. J. Clin. Periodontol. 2004, 31, 515–521. [Google Scholar] [CrossRef]
- Niture, S.K.; Khatri, R.; Jaiswal, A.K. Regulation of Nrf2—An Update. Free Radic. Biol. Med. 2014, 66, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Sima, C.; Aboodi, G.M.; Lakschevitz, F.S.; Sun, C.; Goldberg, M.B.; Glogauer, M. Nuclear Factor Erythroid 2-Related Factor 2 Down-Regulation in Oral Neutrophils Is Associated with Periodontal Oxidative Damage and Severe Chronic Periodontitis. Am. J. Pathol. 2016, 186, 1417–1426. [Google Scholar] [CrossRef] [Green Version]
- Hyeon, S.; Lee, H.; Yang, Y.; Jeong, W. Nrf2 Deficiency Induces Oxidative Stress and Promotes RANKL-Induced Osteoclast Differentiation. Free Radic. Biol. Med. 2013, 65, 789–799. [Google Scholar] [CrossRef]
- Kataoka, K.; Ekuni, D.; Tomofuji, T.; Irie, K.; Kunitomo, M.; Uchida, Y.; Fukuhara, D.; Morita, M. Visualization of Oxidative Stress Induced by Experimental Periodontitis in Keap1-Dependent Oxidative Stress Detector-Luciferase Mice. Int. J. Mol. Sci. Artic. 2016, 17, 1907. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yang, H.; Wen, Y.; Li, B.; Zhao, Y.; Xing, J.; Zhang, M.; Chen, Y. Nrf2 Inhibits Periodontal Ligament Stem Cell Apoptosis under Excessive Oxidative Stress. Int. J. Mol. Sci. Artic. Int. J. Mol. Sci. 2017, 18, 1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasnak, G.; Könönen, E.; Syrjänen, S.; Gürsoy, M.; Zeidán-Chuliá, F.; Firatli, E.; Gürsoy, U.K. NFE2L2/NRF2, OGG1, and Cytokine Responses of Human Gingival Keratinocytes against Oxidative Insults of Various Origin. Mol. Cell. Biochem. 2019, 452, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Dong, G.; Guo, L.; Graves, D.T. The function of dendritic cells in modulating the host response. Mol. Oral Microbiol. 2018, 33, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sztukowska, M.; Ojo, A.; Scott, D.A.; Wang, H.; Lamont, R.J. FOXO Responses to Porphyromonas gingivalis in Epithelial Cells. Cell. Microbiol. 2015, 17, 1605–1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelle, P.; Caribé, V.; Cunha Villar, C.; Romito, G.A.; Pacanaro, A.P.; Cassaro Strunz, C.M.; Yoshio Takada, J.; Luiz, A.; Machado, C.; De Padua Mansur, A. Influence of the Treatment of Periodontal Disease in Serum Concentration of Sirtuin 1 and Mannose-Binding Lectin. J. Periodontol. 2020, 91, 900–905. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, Y.; Gao, J.; Li, T.; Gan, X.; Yu, H.; Xueqi Gan, C.; Key, S. Sirtuin 3 Deficiency Exacerbates Age-Related Periodontal Disease. J. Periodont. Res. 2021, 56, 1163–1173. [Google Scholar] [CrossRef]
- Carcuac, O.; Berglundh, T. Composition of human peri-implantitis and periodontitis lesions. J. Dent. Res. 2014, 93, 1083–1088. [Google Scholar] [CrossRef] [Green Version]
- Liskmann, S.; Zilmer, M.; Vihalemm, T.; Salum, O.; Fischer, K. Correlation of Peri-Implant Health and Myeloperoxidase Levels: A Cross-Sectional Clinical Study. Clin. Oral Implants Res. 2004, 15, 546–552. [Google Scholar] [CrossRef]
- Tözüm, T.F.; Akman, A.C.; Yamalik, N.; Tulunoglu, I.; Turkyilmaz, I.; Karabulut, E.; Kilinc, K.; Cehreli, M.C. Analysis of the Inflammatory Process Around Endosseous Dental Implants and Natural Teeth: Myeloperoxidase Level and Nitric Oxide Metabolism. Int. J. Oral Maxillofac. Implants. 2007, 22, 969–979. [Google Scholar]
- Güncü, G.N.; Tözüm, T.F.; Güncü, B.M.; Yamalik, N.; Tümer, C.; Karabulut, E.; Kilinç, K. Myeloperoxidase as a Measure of Polymorphonuclear Leukocyte Response in Inflammatory Status Around Immediately and Delayed Loaded Dental Implants: A Randomized Controlled Clinical Trial. Clin. Implant Dent. Relat. Res. 2007, 10, 30–39. [Google Scholar] [CrossRef]
- Kasnak, G.; Firatli, E.; Könönen, E.; Olgac, V.; Zeidán-Chuliá, F.; Gursoy, U.K. Elevated Levels of 8-OHdG and PARK7/DJ-1 in Peri-Implantitis Mucosa. Clin. Implant Dent. Relat. Res. 2018, 20, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An Evolving Term within the Functional Foods Field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Cuevas-González, P.F.; Liceaga, A.M.; Aguilar-Toalá, J.E. Postbiotics and Paraprobiotics: From Concepts to Applications. Food Res. Int. 2020, 136, 109502. [Google Scholar] [CrossRef] [PubMed]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics—A Step Beyond Pre- and Probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 416763. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef]
- Valdes-Lopez, O.; Ntatsi, G.; Fotopoulos, V.; Georgiadou, E.C.; Kowalska, E.; Patla, K.; Kulbat, K.; Smolí Nska, B.; Leszczy’nska, J.; Leszczy’nska, L. Influence of Heavy Metals (Ni, Cu, and Zn) on Nitro-Oxidative Stress Responses, Proteome Regulation and Allergen Production in Basil (Ocimum basilicum L.) Plants. Front. Plant Sci. 2018, 9, 862. [Google Scholar] [CrossRef] [Green Version]
- Kullisaar, T.; Zilmer, M.; Mikelsaar, M.; Vihalemm, T.; Annuk, H.; Kairane, C.; Kilk, A. Two Antioxidative Lactobacilli Strains as Promising Probiotics. Int. J. Food Microbiol. 2002, 72, 215–224. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; del Carmen, S.; Miyoshi, A.; Azevedo, V.; Sesma, F.; Langella, P.; Bermúdez-Humarán, L.G.; Watterlot, L.; Perdigon, G.; de Moreno de LeBlanc, A. Use of Superoxide Dismutase and Catalase Producing Lactic Acid Bacteria in TNBS Induced Crohn’s Disease in Mice. J. Biotechnol. 2011, 151, 287–293. [Google Scholar] [CrossRef]
- Amaretti, A.; di Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant Properties of Potentially Probiotic Bacteria: In Vitro and in Vivo Activities. Appl. Microb. Cell Physiol. 2013, 97, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Ejtahed, H.S.; Mohtadi-Nia, J.; Homayouni-Rad, A.; Niafar, M.; Asghari-Jafarabadi, M.; Mofid, V. Probiotic Yogurt Improves Antioxidant Status in Type 2 Diabetic Patients. Nutrition 2012, 28, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Kodali, V.P.; Sen, R. Antioxidant and free radical scavenging activities of an exopolysaccharide from a probiotic bacterium. Biotech. J. 2008, 3, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Afify, A.E.M.R.; Romeilah, R.M.; Sultan, S.I.M.; Hussein, M.M.; Afify, A.E.M.R.; Romeilah, R.M.; Shaimaa, I.M.; Hussein, M.M. Antioxidant Activity And Biological Evaluations Of Probiotic Bacteria Strains. Int. J. Acad. Res. 2012, 4, 131–139. [Google Scholar] [CrossRef]
- Fakruddin, M.; Ahmed, M.M. Antimicrobial and Antioxidant Activities of Saccharomyces cerevisiae IFST062013, a Potential Probiotic. BMC Complement Altern. Med. 2017, 17, 64. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, J.S.; Kim, Y.; Jeong, Y.; Kim, J.E.; Paek, N.S.; Kang, C.H. Antioxidant and Probiotic Properties of Lactobacilli and Bifidobacteria of Human Origins. Biotechnol. Bioprocess Eng. 2020, 25, 421–430. [Google Scholar] [CrossRef]
- Gomes, A.C.; de Sousa, R.G.M.; Botelho, P.B.; Gomes, T.L.N.; Prada, P.O.; Mota, J.F. The Additional Effects of a Probiotic Mix on Abdominal Adiposity and Antioxidant Status: A Double-Blind, Randomized Trial. Obesity 2017, 25, 30–38. [Google Scholar] [CrossRef]
- Yang, S.J.; Lee, J.E.; Lim, S.M.; Kim, Y.J.; Lee, N.K.; Paik, H.D. Antioxidant and Immune-Enhancing Effects of Probiotic Lactobacillus plantarum 200655 Isolated from Kimchi. Food Sci. Biotechnol. 2017, 25, 30–38. [Google Scholar] [CrossRef]
- Bonfili, L.; Cecarini, V.; Cuccioloni, M.; Angeletti, M.; Berardi, S.; Scarpona, S.; Rossi, G.; Eleuteri, A.M. SLAB51 Probiotic Formulation Activates SIRT1 Pathway Promoting Antioxidant and Neuroprotective Effects in an AD Mouse Model. Mol. Neurobiol. 2018, 55, 7987–8000. [Google Scholar] [CrossRef] [Green Version]
- Aboulgheit, A.; Karbasiafshar, C.; Zhang, Z.; Sabra, M.; Shi, G.; Tucker, A.; Sodha, N.; Abid, M.R.; Sellke, F.W. Lactobacillus plantarum Probiotic Induces Nrf2-Mediated Antioxidant Signaling and ENOS Expression Resulting in Improvement of Myocardial Diastolic Function. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H839–H849. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Yang, Z.; Yang, W.; Huang, L.; Xu, C.; Liu, M.; Ge, J.; Wang, Y.; Jiang, S. Illicium verum Extracts and Probiotics with Added Glucose Oxidase Promote Antioxidant Capacity through Upregulating Hepatic and Jejunal Nrf2/Keap1 of Weaned Piglets. J. Anim. Sci. 2020, 98, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Maherinia, H.; Peeri, M.; Azarbayjani, M.A.; Delfan, M. Aerobic Exercise Training Combined with Probiotic Supplement Improves Antioxidant Defence of Cardiomyocytes by Regulating Nrf2 and Caspase3 Gene Expression in Type 2 Diabetic Rats. Comp. Exerc. Physiol. 2022, 0, 1–10. [Google Scholar] [CrossRef]
- Xu, H.; Wang, J.; Cai, J.; Feng, W.; Wang, Y.; Liu, Q.; Cai, L. Protective Effect of Lactobacillus rhamnosus GG and Its Supernatant against Myocardial Dysfunction in Obese Mice Exposed to Intermittent Hypoxia Is Associated with the Activation of Nrf2 Pathway. Int. J. Biol. Sci. 2019, 15, 2471. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, B.J.; Liu, K.H.; Owens, J.A.; Hunter-Chang, S.; Camacho, M.C.; Eboka, R.U.; Chandrasekharan, B.; Baker, N.F.; Darby, T.M.; Robinson, B.S.; et al. Gut-Resident Lactobacilli Activate Hepatic Nrf2 and Protect Against Oxidative Liver Injury. Cell Metab. 2020, 31, 956–968. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Wang, C.; Zhang, L.; Zhao, Y.; Duan, C.; Zhang, X.; Gao, L.; Li, S. Lactobacillus plantarum NA136 Improves the Non-Alcoholic Fatty Liver Disease by Modulating the AMPK/Nrf2 Pathway. Appl. Microbiol. Biotechnol. 2019, 103, 5843–5850. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, B.J.; Robinson, B.S.; Owens, J.; Liu, K.; Eboka, R.; Darby, T.; Luo, L.; Jones, D.; Jones, R.; Neish, A. Regulation of the Hepatic Antioxidant Response by the Probiotic Lactobacillus rhamnosus GG. FASEB J. 2019, 33, 369.5. [Google Scholar] [CrossRef]
- Endo, H.; Niioka, M.; Kobayashi, N.; Tanaka, M.; Watanabe, T. Butyrate-Producing Probiotics Reduce Nonalcoholic Fatty Liver Disease Progression in Rats: New Insight into the Probiotics for the Gut-Liver Axis. PLoS ONE 2013, 8, e63388. [Google Scholar] [CrossRef] [Green Version]
- Feng, T.; Wang, J. Oxidative Stress Tolerance and Antioxidant Capacity of Lactic Acid Bacteria as Probiotic: A Systematic Review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef]
- Lebeer, S.; Claes, I.J.J.; Verhoeven, T.L.A.; Vanderleyden, J.; De Keersmaecker, S.C.J. Exopolysaccharides of Lactobacillus rhamnosus GG form a protective shield against innate immune factors in the intestine. Microb. Biotechnol. 2011, 4, 368–374. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ji, J.; Chen, X.; Jiang, M.; Rui, X.; Dong, M. Structural Elucidation and Antioxidant Activities of Exopolysaccharides from Lactobacillus helveticus MB2-1. Carbohydr. Polym. 2014, 102, 351–359. [Google Scholar] [CrossRef]
- Rani, R.P.; Anandharaj, M.; David Ravindran, A. Characterization of a Novel Exopolysaccharide Produced by Lactobacillus gasseri FR4 and Demonstration of Its in Vitro Biological Properties. Int. J. Biol. Macromol. 2018, 109, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Q.; Gao, N.; Wang, Z.; Li, F.; Li, J.; Shan, A. Exopolysaccharides Produced by Lactobacillus rhamnosus GG Alleviate Hydrogen Peroxide-Induced Intestinal Oxidative Damage and Apoptosis through the Keap1/Nrf2 and Bax/Bcl-2 Pathways in vitro. Food Func. 2021, 12, 9632–9641. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 2015, 74, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.M.A.E.; Bast, A.; Vanhoutvin, S.A.L.W.; Fischer, M.A.J.G.; Kodde, A.; Troost, F.J.; Venema, K.; Brummer, R.J.M. Butyrate Modulates Oxidative Stress in the Colonic Mucosa of Healthy Humans. Clin. Nutr. 2009, 28, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Jia, Y.; Yang, S.; Zhao, N.; Hu, Y.; Hong, J.; Gao, S.; Zhao, R. Sodium Butyrate Protects against High-Fat Diet-Induced Oxidative Stress in Rat Liver by Promoting Expression of Nuclear Factor E2-Related Factor 2. Br. J. Nutr. 2019, 122, 400–410. [Google Scholar] [CrossRef]
- Guan, X.; Li, W.; Meng, H. A double-edged sword: Role of butyrate in the oral cavity and the gut. Mol. Oral Microbiol. 2021, 36, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Ben-Dor, A.; Steiner, M.; Gheber, L.; Danilenko, M.; Dubi, N.; Linnewiel, K.; Zick, A.; Sharoni, Y.; Levy, J. Carotenoids Activate the Antioxidant Response Element Transcription System. Mol. Cancer Ther. 2005, 4, 177–186. [Google Scholar] [CrossRef]
- Hagi, T.; Kobayashi, M.; Nomura, M. Aerobic Condition Increases Carotenoid Production Associated with Oxidative Stress Tolerance in Enterococcus gilvus. FEMS Microbiol. Lett. 2014, 350, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Seo, D.H.; Park, Y.S.; Cha, I.T.; Seo, M.J. Isolation of Lactobacillus plantarum subsp. plantarum Producing C30 Carotenoid 4,4′-Diaponeurosporene and the Assessment of Its Antioxidant Activity. J. Microbiol. Biotechnol. 2019, 29, 1925–1930. [Google Scholar] [CrossRef]
- Lee, Y.; Yoon, Y.; Choi, K.H. Probiotics-Mediated Bioconversion and Periodontitis. Food Sci. Anim. 2021, 41, 905–922. [Google Scholar] [CrossRef] [PubMed]
- Muro Urista, C.; Lvarez Fernández, R.A.; Riera Rodriguez, F.; Cuenca, A.A.; Jurado, A.T. Review: Production and Functionality of Active Peptides from Milk. Food Sci. Technol. Int. 2011, 17, 293–317. [Google Scholar] [CrossRef] [PubMed]
- Suthanthangjai, W.; Kilmartin, P.A.; Phillips, A.R.J.; Davies, K.; Ansell, J. Bioconversion of Pinot Noir Anthocyanins into Bioactive Phenolic Compounds by Lactic Acid Bacteria. Nutr. Aging. 2014, 2, 145–149. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Hong, G.E.; Zhang, H.; Yang, C.Y.; Han, K.H.; Mandal, K.; Lee, C.H. Production of the Isoflavone Aglycone and Antioxidant Activities in Black Soymilk Using Fermentation with Streptococcus thermophilus S10. Food Sci. Biotechnol. 2015, 24, 537–544. [Google Scholar] [CrossRef]
- Lee, J.; Kim, M.; Kim, D.; Yu, C.; Kim, B.S.; Lee, J.S.; Kang, S.S. Anti-Biofilm Effect of Bioconversion of Whey by Lactic Acid Bacteria against Foodborne Pathogenic Bacteria. Curr. Top. Lact. Acid Bact. Probiotics 2020, 6, 25–31. [Google Scholar] [CrossRef]
- Rogozinska, M.; Korsak, D.; Mroczek, J.; Biesaga, M. Catabolism of Hydroxycinnamic Acids in Contact with Probiotic Lactobacillus. J. Appl. Microbiol. 2021, 131, 1464–1473. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Hong, Q.; Wang, Y.; Liang, Q.; Tan, H.; Xiao, C.; Tang, X.; Shao, S.; Zhou, S.; Gao, Y. Ferulic Acid Induces Heme Oxygenase-1 via Activation of ERK and Nrf2. Drug Discov. Ther. 2011, 5, 299–305. [Google Scholar] [CrossRef] [Green Version]
- San Miguel, S.M.; Opperman, L.A.; Allen, E.P.; Zielinski, J.; Svoboda, K.K.H. Bioactive Antioxidant Mixtures Promote Proliferation and Migration on Human Oral Fibroblasts. Arch. Oral Biol. 2011, 56, 812–822. [Google Scholar] [CrossRef]
- Yang, S.Y.; Cheol Pyo, M.; Nam, M.H.; Lee, K.W. ERK/Nrf2 Pathway Activation by Caffeic Acid in HepG2 Cells Alleviates Its Hepatocellular Damage Caused by t-Butylhydroperoxide-Induced Oxidative Stress. BMC Complement. Altern. Med. 2019, 19, 139. [Google Scholar] [CrossRef] [Green Version]
- Özden, F.O.; Lütfioğlu, M.; Demir, E.; Bilgici, B. Antioxidant Effect of Caffeic Acid Phenethyl Ester in Experimentally Induced Periodontitis. Clin. Oral Investig. 2021, 25, 4959–4966. [Google Scholar] [CrossRef]
- D’Amico, D.; Andreux, P.A.; Valdés, P.; Singh, A.; Rinsch, C.; Auwerx, J. Impact of the Natural Compound Urolithin A on Health, Disease, and Aging. Trends Mol. Med. 2021, 27, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the Gut Barrier Integrity by a Microbial Metabolite through the Nrf2 Pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef] [Green Version]
- Tabasco, R.; Sánchez-Patán, F.; Monagas, M.; Bartolomé, B.; Victoria Moreno-Arribas, M.; Peláez, C.; Requena, T. Effect of Grape Polyphenols on Lactic Acid Bacteria and Bifidobacteria Growth: Resistance and Metabolism. Food Microbiol. 2011, 28, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.T.; Wu, C.H.; Ho, C.Y.; Yen, G.C. Catechin Protects against Ketoprofen-Induced Oxidative Damage of the Gastric Mucosa by up-Regulating Nrf2 in vitro and in vivo. J. Nutr. Biochem. 2013, 24, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cui, Y.J.; Bai, S.S.; Yang, Z.J.; Miao-Cai; Megrous, S.; Aziz, T.; Sarwar, A.; Li, D.; Yang, Z.N. Antioxidant Activity of Novel Casein-Derived Peptides with Microbial Proteases as Characterized via Keap1-Nrf2 Pathway in HepG2 Cells. J. Microbiol. Biotechnol. 2021, 31, 1163–1174. [Google Scholar] [CrossRef]
- Yokoji-Takeuchi, M.; Takahashi, N.; Yamada-Hara, M.; Sulijaya, B.; Tsuzuno, T.; Aoki-Nonaka, Y.; Tabeta, K.; Kishino, S.; Ogawa, J.; Yamazaki, K. A Bacterial Metabolite Induces Nrf2-Mediated Anti-Oxidative Responses in Gingival Epithelial Cells by Activating the MAPK Signaling Pathway. Arch. Oral Biol. 2020, 110, 104602. [Google Scholar] [CrossRef]
- Campbell, N.K.; Fitzgerald, H.K.; Dunne, A. Regulation of Inflammation by the Antioxidant Haem Oxygenase 1. Nat. Rev. Immunol. 2021, 21, 411–425. [Google Scholar] [CrossRef]
- Vo, T.T.T.; Huang, H.W.; Wee, Y.; Feng, S.W.; Cheng, H.C.; Tuan, V.P.; Lee, I.T. Surfactin reduces particulate matter–induced VCAM-1–dependent monocyte adhesion in human gingival fibroblasts by increasing Nrf2-dependent HO-1 expression. J. Periodont. Res. 2022, 57, 115–130. [Google Scholar] [CrossRef]
- Graziani, F.; Karapetsa, D.; Alonso, B.; Herrera, D. Nonsurgical and surgical treatment of periodontitis: How many options for one disease? Periodontol. 2000 2017, 75, 152–188. [Google Scholar] [CrossRef]
- Van der Weijden, G.A.; Dekkers, G.J.; Slot, D.E. Success of non-surgical periodontal therapy in adult periodontitis patients: A retrospective analysis. Int. J. Dent. Hyg. 2019, 17, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Bartold, P.M.; Van Dyke, T.E. Host modulation: Controlling the inflammation to control the infection. Periodontol. 2000 2017, 75, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Preshaw, P.M. Host modulation therapy with anti-inflammatory agents. Periodontol. 2000 2018, 76, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Mazzon, E.; Muià, C.; Crisafulli, C.; Terrana, D.; Greco, S.; Britti, D.; Santori, D.; Oteri, G.; Cordasco, G.; et al. Effects of Etanercept, a Tumour Necrosis Factor-a Antagonist, in an Experimental Model of Periodontitis in Rats. Br. J. Pharmacol. 2007, 150, 286–297. [Google Scholar] [CrossRef] [Green Version]
- Araújo, R.F.D.; Souza, T.O.; Moura, L.M.D.; Torres, K.P.; Souza, L.B.D.; Alves, M.D.S.C.F.; Rocha, H.O.; de Araújo, A.A. Atorvastatin decreases bone loss, inflammation and oxidative stress in experimental periodontitis. PLoS ONE 2013, 8, e75322. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.D.R.; Lopes, A.P.; Martins, C.; Brito, G.A.; Carneiro, V.C.; Goes, P. The effect of Calendula offiinalis on oxidative stress and bone loss in experimental periodontitis. Front. Physiol. 2017, 1, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, A.H.; Freire, J.M.D.O.; de Sousa, L.H.T.; Parente, A.T.; de Sousa, N.A.; Arriaga, A.M.C.; da Silva, F.R.L.; Melo, I.M.; da Silva, I.I.C.; Pereira, K.M.A.; et al. Stemodia maritima L. extract decreases inflammation, oxidative stress, and alveolar bone loss in an experimental periodontitis rat model. Front. Physiol. 2017, 8, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Araújo, A.A.; De Morais, H.B.; De Medeiros, C.A.C.X.; De Castro Brito, G.A.; Guedes, P.M.M.; Hiyari, S.; Pirih, F.Q.; De Araújo Júnior, R.F. Gliclazide Reduced Oxidative Stress, Inflammation, and Bone Loss in an Experimental Periodontal Disease Model. J. Appl. Oral Sci. 2019, 27, 1–14. [Google Scholar] [CrossRef]
- Donos, N.; Calciolari, E.; Brusselaers, N.; Goldoni, M.; Bostanci, N.; Belibasakis, G.N. The Adjunctive Use of Host Modulators in Non-Surgical Periodontal Therapy. A Systematic Review of Randomized, Placebo-Controlled Clinical Studies. J. Clin. Periodontol. 2020, 47, 199–238. [Google Scholar] [CrossRef] [Green Version]
- Sanz, M.; Herrera, D.; Kebschull, M.; Chapple, I.; Jepsen, S.; Berglundh, T.; Sculean, A.; Tonetti, M.S. Treatment of stage I–III periodontitis—The EFP S3 level clinical practice guideline. J. Clin. Periodontol. 2020, 47, 4–60. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probiotics | Antioxidant Source | Mode of Action | Reference |
|---|---|---|---|
| Bifidobacterium, Lactobacillus, Lactococcus, Streptococcus thermophilus | Intact cells and spent culture media * | Scavenging activity on radicals | [51] |
| Lactobacillus bulgaricus, Streptococcus thermophilus, Bifidobacterium lactis Bb12 and Lactobacillus acidophilus La5 strains | Yogurt including indicated probiotics | Increased erythrocyte superoxide dismutase and glutathione peroxidase activities and total antioxidant status in Type 2 diabetes | [52] |
| Bacillus coagulans RK-02 | B. coagulans derived exopolysaccharides * | Superoxide radical scavenging activity | [53] |
| Lactobacillus rhamnosus GG, Lactobacillus retueria (ATCC 20016), Bifidobacterium breve (ATCC 15700), Probionebacterium freudenreichii ssp. | Cell free culture extract * | DPPH scavenging activity | [54] |
| Saccharomyces cerevisiae IFST062013 | Cell extract and autolysate * | Hydroxyl radical and nitric oxide scavencing activity | [55] |
| Human originated Lactobacilli and Bifidobacteria strains | Living cells | DPPH and ABTS scavenging activity | [56] |
| Lactobacillus acidophilus LA-14, Lactobacillus casei LC-11, Lactococcus lactis LL-23, Bifidobacterium bifidum BB-06, and Bifidobacterium lactis BL-4 | Commercial probiotic Danisco® | Increased activity of glutathione peroxidase | [57] |
| Lactobacillus plantarum 200655 | Living cells isolated from kimchi | DPPH and ABTS scavenging activity | [58] |
| SLAB51 Probiotic Formulation | Living cells | Activates SIRT1 pathway promoting antioxidant effects | [59] |
| Source | Antioxidant Activity | Reference |
|---|---|---|
| Lactobacillus plantarum | Increased production of caffeic acid by L. plantarum in combination with apple juice promotes Nrf2 activation, resulting in improved diastolic function in chronic ischemic myocardium. | [60] |
| Commercial probiotic | Probiotics combined with Illicium verum extract and glucose oxidase enzyme upregulate hepatic and jejunal Nrf2/Keap1 pathway. | [61] |
| Commercial probiotic | Probiotic supplement improves antioxidant defence of cardiomyocytes by regulating Nrf2 and caspase3 gene expression in type 2 diabetic rats. | [62] |
| Lactobacillus rhamnosus GG and its spent culture supernatant | Improvement of myocardial dysfunction in obese mice exposed to intermittent hypoxia by activating Nrf2 Pathway | [63] |
| Lactobacillus rhamnosus GG | Significant induction of Nrf2 target transcripts in liver tissue via situmulation of xenobiotic | [64] |
| Lactobacillus plantarum NA136 | Increased the nuclear translocation of Nrf2 leading to improved antioxidant response in case of non-alcoholic fatty liver disease | [65] |
| Gut-resident Lactobacilli | Activation of Nrf2 response against oxidative liver injury through gut-liver axis | [66] |
| MIYAIRI 588–a butyrate-producing probiotic strain | Reduced nonalcoholic fatty liver disease progression via Nrf2 pathway | [67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karaca, B.; Yilmaz, M.; Gursoy, U.K. Targeting Nrf2 with Probiotics and Postbiotics in the Treatment of Periodontitis. Biomolecules 2022, 12, 729. https://doi.org/10.3390/biom12050729
Karaca B, Yilmaz M, Gursoy UK. Targeting Nrf2 with Probiotics and Postbiotics in the Treatment of Periodontitis. Biomolecules. 2022; 12(5):729. https://doi.org/10.3390/biom12050729
Chicago/Turabian StyleKaraca, Basar, Mustafa Yilmaz, and Ulvi Kahraman Gursoy. 2022. "Targeting Nrf2 with Probiotics and Postbiotics in the Treatment of Periodontitis" Biomolecules 12, no. 5: 729. https://doi.org/10.3390/biom12050729
APA StyleKaraca, B., Yilmaz, M., & Gursoy, U. K. (2022). Targeting Nrf2 with Probiotics and Postbiotics in the Treatment of Periodontitis. Biomolecules, 12(5), 729. https://doi.org/10.3390/biom12050729