
A Comparative Study of Milk Fat Extracted from the Milk of Different Goat Breeds in China: Fatty Acids, Triacylglycerols and Thermal and Spectroscopic Characterization

 ,
,  ,
, 
 ,
,  ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Milk Sample Collection
2.2. Chemicals and Reagents
2.3. Total Fat Extraction
2.4. Analysis of Extracted Fat Composition
2.4.1. FA Composition
2.4.2. FA Distribution at sn-2 and sn-1, 3 Positions
2.4.3. Separation and Identification of TAG Species by UPCC-Q-TOF-MS
2.5. Physicochemical Characterization of the Extracted Fat
2.5.1. Melting and Crystallization Profiles
2.5.2. Infrared Spectroscopy
2.6. Statistical Analysis
3. Results and Discussion
3.1. FA Compositions and Distribution
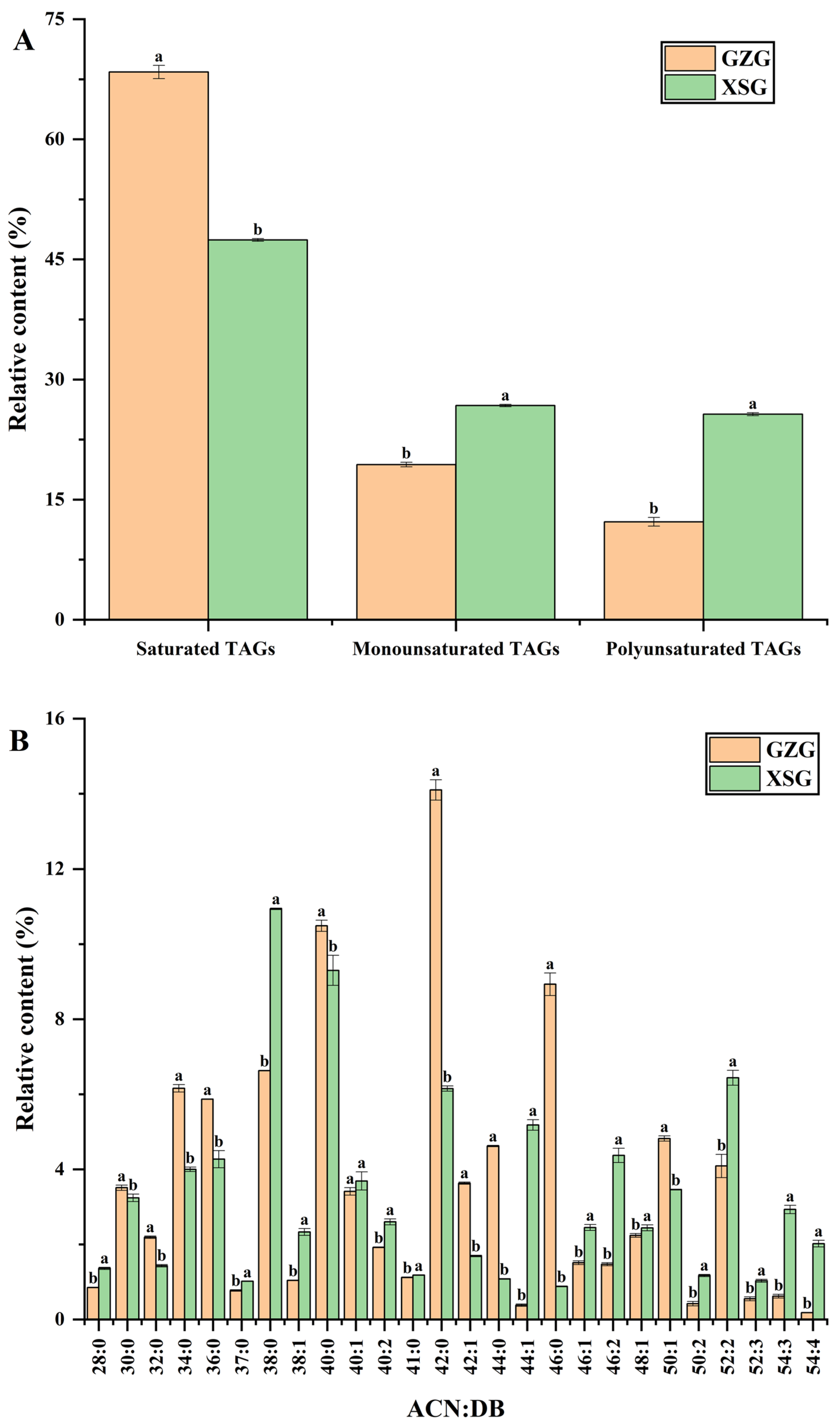
3.2. TAG Molecular Species Profiling
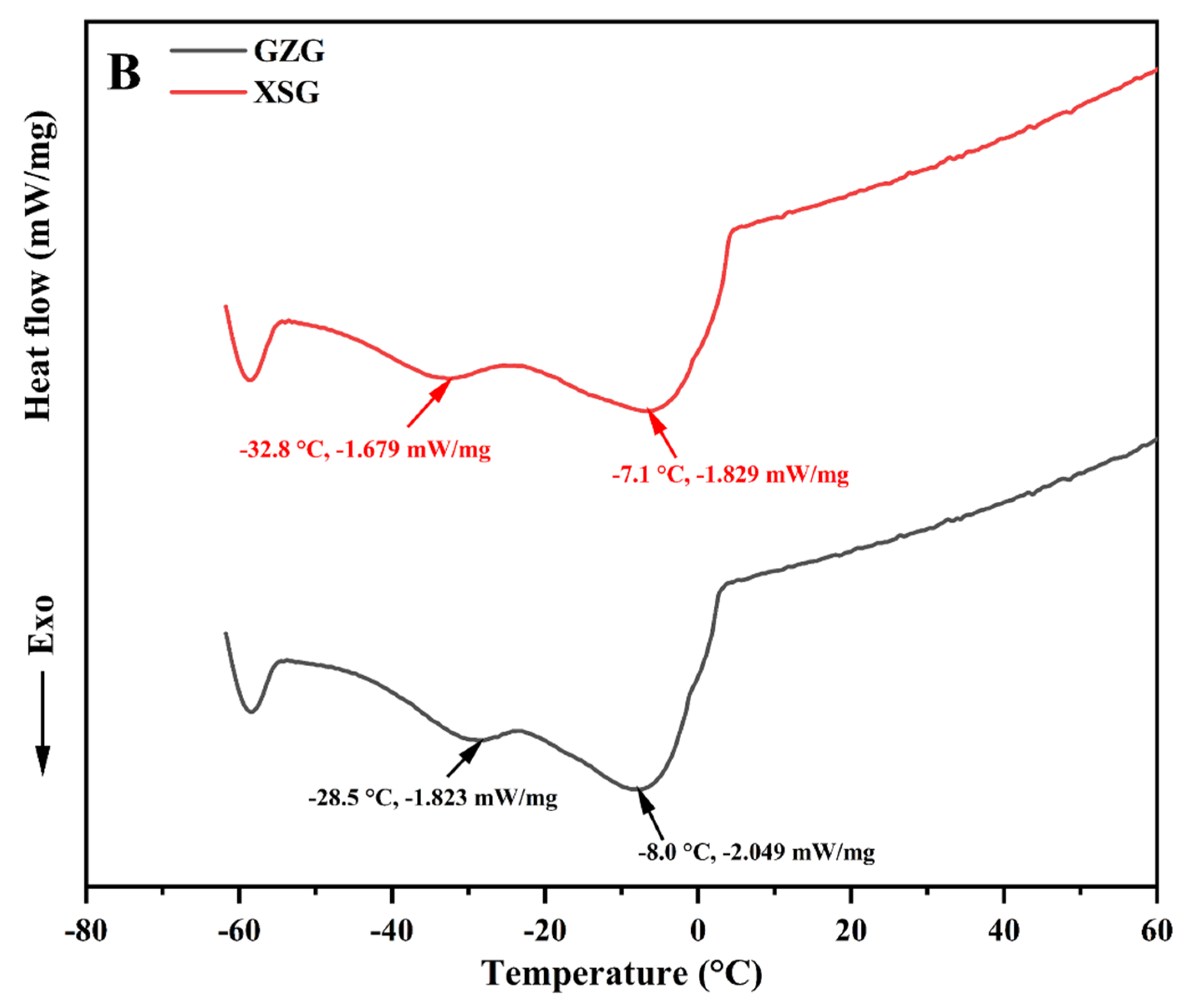
3.3. Melting and Crystallization Thermal Profiles
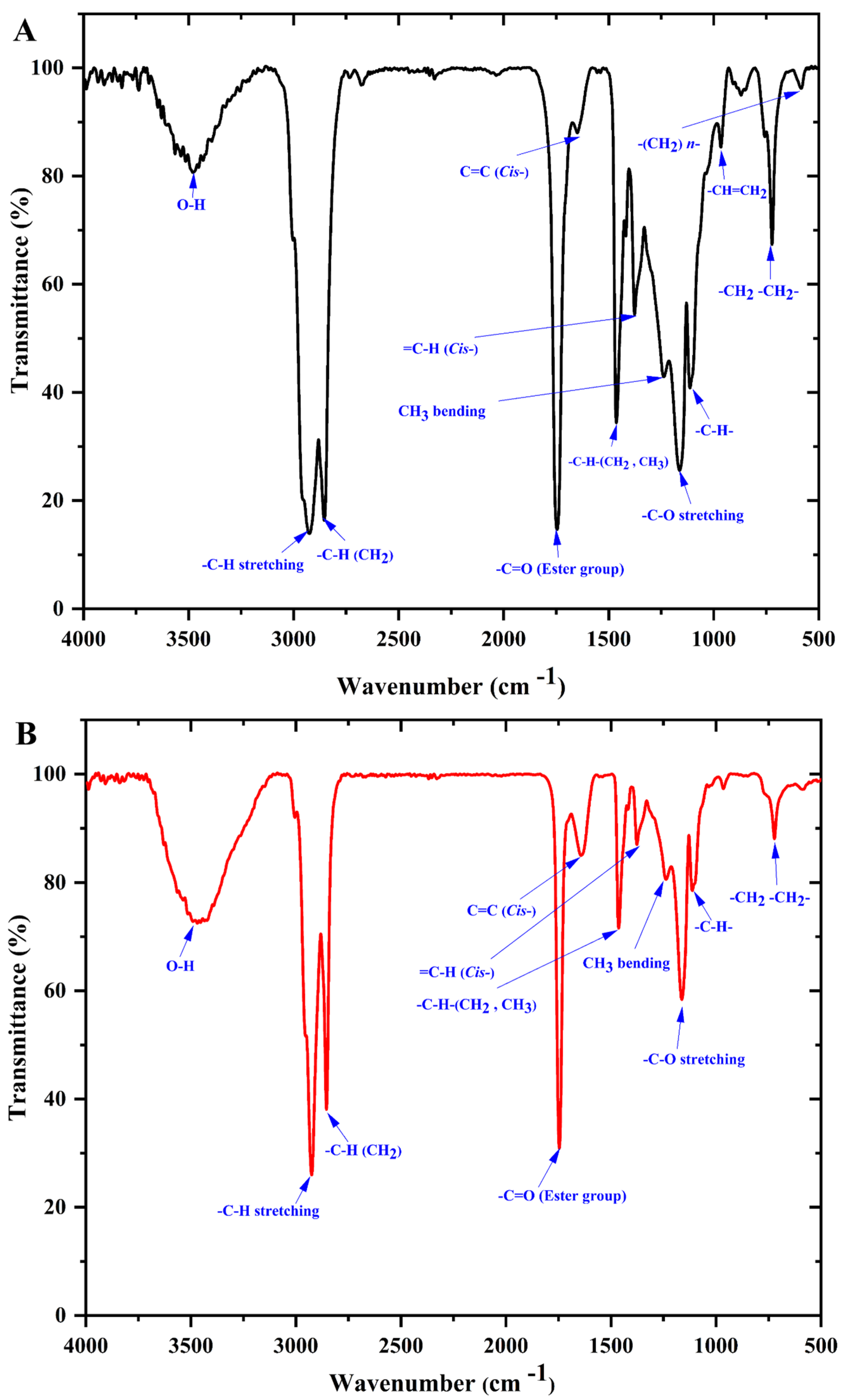
3.4. Infrared Spectroscopy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GM | Goat milk |
| GZG | Guanzhong goat milk |
| XSG | Xinong Saanen goat milk |
| MF | Milk fat |
| GMF | Goat milk fat |
| IF | Infant formula |
| FAs | Fatty acids |
| TAGs | Triacylglycerols |
| PN | Partition number |
| ACN | Acyl carbon numbers |
| DB | Double bond number |
| FT-IR | Fourier-transform infrared spectroscopy |
| DSC | Differential scanning calorimetry |
| UPCC | Ultra-performance convergence chromatography |
| UPLC | Ultra-performance liquid chromatography |
| Q-TOF-MS | Quadrupole time-of-flight mass spectrometry |
| RP-HPLC | Reverse-phase high-performance liquid chromatography |
| APCI-MS | Atmospheric pressure chemical ionization mass spectrometry |
| HR-MS/MS | High-resolution tandem mass spectrometry |
| GC | Gas chromatography |
| FAMEs | Fatty acid methyl esters |
| TLC | Thin-layer chromatography plates |
| DAG | Diglyceride |
| MAG | Monoacylglycerol |
| SC-SFAs | Short-chain saturated fatty acids |
| MC-SFAs | Medium-chain saturated fatty acids |
| LC-SFAs | Long-chain saturated fatty acids |
| SFAs | Saturated fatty acids |
| UFAs | Unsaturated fatty acids |
| MUFAs | Monounsaturated fatty acids |
| PUFAs | Polyunsaturated fatty acids |
| OCS-FAs | Odd chain saturated fatty acids |
| ECS-FAs | Even chain saturated fatty acids |
| n-6 | All n-6 fatty acids |
| n-3 | All n-3 fatty acids |
| n-6/n-3 | The ratio between all n-6 and all n-3 |
| L/Ln | The ratio between C18:2 n-6c and C18:3 n-3c |
| Bu | Butyric acid (C4:0) |
| Co | Caproic acid (C6:0) |
| Cy | Caprylic acid (C8:0) |
| Ca | Capric acid (C10:0) |
| De | Decenoic acid (C10:1) |
| Ud | Undecanoic acid (C11:0) |
| La | Lauric acid (C12:0) |
| Lo | Lauroleic acid (C12:1) |
| Tr | Tridecanoic acid (C13:0) |
| M | Myristic acid (C14:0) |
| Mo | Myristoleic acid (C14:1 n-5c) |
| Pe | Pentadecylic acid (C15:0) |
| Pen | Pentadecenoic acid (C15:1 n-5c) |
| P | Palmitic acid (C16:0) |
| Po | Palmitoleic acid (C16:1 n-7c) |
| Ma | Margaric acid (C17:0) |
| H | Heptadecenoic acid (C17:1 n-7t) |
| S | Stearic acid (C18:0) |
| O | Oleic acid (C18:1 n-9c) |
| L | Linoleic acid (C18:2 n-6c) |
| Gln | Gamma-linolenic acid (C18:3 n-6c) |
| Ln | Alpha-linolenic acid (C18:3 n-3c) |
| N | Nonadecanoic acid (C19:0) |
| No | Nonadecenoic acid (C19:1) |
| Ar | Arachidic acid (C20:0) |
| Go | Gadoleic acid (C20:1) |
| ARA | Arachidonic acid (C20:4 n-6c) |
| Beh | Behenic acid (C22:0) |
| Er | Erucic acid (C22:1) |
References
- Wang, G.; Chen, S.; Chao, T.; Ji, Z.; Hou, L.; Qin, Z.; Wang, J. Analysis of genetic diversity of Chinese dairy goats via microsatellite markers. J. Anim. Sci. 2017, 95, 2304–2313. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.A.; Lu, C.D. Current status of global dairy goat production: An overview. Asian Australas. J. Anim. Sci. 2019, 32, 1219. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B. Animal Genetic Resources in China, Sheep and Goat, China National Commission of Animal Genetic Resources End; China Agricultural Press: Beijing, China, 2011. [Google Scholar]
- Qi, J.; Ling, J.; Tian, H. China’s Livestock and Poultry Genetic Resources Condition; China Agriculture Press: Beijing, China, 2004. [Google Scholar]
- Lu, J.; Zhang, Y.; Song, B.; Zhang, S.; Pang, X.; Sari, R.N.; Liu, L.; Wang, J.; Lv, J. Comparative analysis of oligosaccharides in Guanzhong and Saanen goat milk by using LC–MS/MS. Carbohydr. Polym. 2020, 235, 115965. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Pi, X.; Ji, Z.; Qin, Z.; Hou, L.; Chao, T.; Wang, J. Investigation of the diversity and origins of Chinese dairy goats via the mitochondrial DNA D-loop. J. Anim. Sci. 2015, 93, 949–955. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, Z.; Liu, C.; Liu, Y. Lipid profiling and microstructure characteristics of goat milk fat from different stages of lactation. J. Agric. Food Chem. 2020, 68, 7204–7213. [Google Scholar] [CrossRef]
- Dolatowska-Żebrowska, K.; Ostrowska-Ligęza, E.; Wirkowska-Wojdyła, M.; Bryś, J.; Górska, A. Characterization of thermal properties of goat milk fat and goat milk chocolate by using DSC, PDSC and TGA methods. J. Therm. Anal. Calorim. 2019, 138, 2769–2779. [Google Scholar] [CrossRef] [Green Version]
- Reddy, R.S.; Ramachandra, C.; Hiregoudar, S.; Nidoni, U.; Ram, J.; Kammar, M. Influence of processing conditions on functional and reconstitution properties of milk powder made from Osmanabadi goat milk by spray drying. Small Rumin. Res. 2014, 119, 130–137. [Google Scholar] [CrossRef]
- Nosratpour, M.; Wang, Y.; Woo, M.W.; Selomulya, C. Characterisation of thermal and structural behaviour of lipid blends composed of fish oil and milkfat. Food Res. Int. 2020, 137, 109377. [Google Scholar] [CrossRef]
- Gantner, V.; Mijić, P.; Baban, M.; Škrtić, Z.; Turalija, A. The overall and fat composition of milk of various species. Mljekarstvo 2015, 65, 223–231. [Google Scholar] [CrossRef] [Green Version]
- Lopez, C.; Briard-Bion, V.; Ménard, O. Polar lipids, sphingomyelin and long-chain unsaturated fatty acids from the milk fat globule membrane are increased in milks produced by cows fed fresh pasture based diet during spring. Food Res. Int. 2014, 58, 59–68. [Google Scholar] [CrossRef]
- Getaneh, G.; Mebrat, A.; Wubie, A.; Kendie, H. Review on goat milk composition and its nutritive value. J. Nutrs. Health Sci. 2016, 3, 401–409. [Google Scholar]
- Pisanu, S.; Marogna, G.; Pagnozzi, D.; Piccinini, M.; Leo, G.; Tanca, A.; Roggio, A.M.; Roggio, T.; Uzzau, S.; Addis, M.F. Characterization of size and composition of milk fat globules from Sarda and Saanen dairy goats. Small Rumin. Res. 2013, 109, 141–151. [Google Scholar] [CrossRef]
- Man, Y.B.C. Analysis of canola oil in virgin coconut oil using FTIR spectroscopy and chemometrics. J. Food Pharm. Sci. 2013, 1, 5–9. [Google Scholar]
- Nurrulhidayah, A.; Rohman, A.; Amin, I.; Shuhaimi, M.; Khatib, A. Analysis of chicken fat as adulterant in butter using Fourier transform infrared spectroscopy and chemometrics. Grasas Aceites 2013, 64, 349–355. [Google Scholar]
- Nurrulhidayah, A.; Che Man, Y.; Amin, I.; Arieff Salleh, R.; Farawahidah, M.; Shuhaimi, M.; Khatib, A. FTIR-ATR spectroscopy based metabolite fingerprinting as a direct determination of butter adulterated with lard. Int. J. Food Prop. 2015, 18, 372–379. [Google Scholar] [CrossRef]
- Fadzlillah, N.A.; Rohman, A.; Ismail, A.; Mustafa, S.; Khatib, A. Application of FTIR-ATR spectroscopy coupled with multivariate analysis for rapid estimation of butter adulteration. J. Oleo Sci. 2013, 62, 555–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakry, I.A.; Ali, A.H.; Abdeen, E.-S.M.; Ghazal, A.F.; Wei, W.; Wang, X. Comparative characterisation of fat fractions extracted from Egyptian and Chinese camel milk. Int. Dairy J. 2020, 105, 104691. [Google Scholar] [CrossRef]
- Tan, C.; Man, Y.C. Comparative differential scanning calorimetric analysis of vegetable oils: I. Effects of heating rate variation. Phytochem. Anal. Int. J. Plant Chem. Biochem. Tech. 2002, 13, 129–141. [Google Scholar] [CrossRef]
- Ali, A.H.; Wei, W.; Abed, S.M.; Korma, S.A.; Mousa, A.H.; Hassan, H.M.; Jin, Q.; Wang, X. Impact of technological processes on buffalo and bovine milk fat crystallization behavior and milk fat globule membrane phospholipids profile. LWT Food Sci. Technol. 2018, 90, 424–432. [Google Scholar] [CrossRef]
- Ben Amara-Dali, W.; Lopez, C.; Lesieur, P.; Ollivon, M. Crystallization properties and polymorphism of triacylglycerols in goat’s milk fat globules. J. Agric. Food Chem. 2008, 56, 4511–4522. [Google Scholar] [CrossRef]
- MacGibbon, A.K.H.; Taylor, M.W. Composition and structure of bovine milk lipids. In Advanced Dairy Chemistry, 3rd ed.; Fox, P.F., McSweeney, P.L.H., Eds.; Springer: New York, NY, USA, 2006; Volume 2, pp. 1–42. [Google Scholar]
- Liu, Z.Q.; Rochfort, S.; Cocks, B. Milk lipidomics: What we know and what we don’t. Prog. Lipid Res. 2018, 71, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Johanson, R.A.; Shaw, M.A.; Buccafusca, R.; Berry, G.T. Expanded MALDI-TOF MS detection of lipids in crude tissue extracts. Faseb. J. 2006, 20, A1350. [Google Scholar]
- Gidden, J.; Liyanage, R.; Durham, B.; Lay, J.O. Reducing fragmentation observed in the matrix-assisted laser desorption/ionization time-of-flight mass spectrometric analysis of triacylglycerols in vegetable oils. Rapid Commun. Mass Spectrom. 2007, 21, 1951–1957. [Google Scholar] [CrossRef]
- Garcia, J.S.; Sanvido, G.B.; Saraiva, S.A.; Zacca, J.J.; Cosso, R.G.; Eberlin, M.N. Bovine milk powder adulteration with vegetable oils or fats revealed by MALDI-QTOF MS. Food Chem. 2012, 131, 722–726. [Google Scholar] [CrossRef]
- Picariello, G.; Sacchi, R.; Addeo, F. One-step characterization of triacylglycerols from animal fat by MALDI-TOF MS. Eur. J. Lipid Sci. Technol. 2007, 109, 511–524. [Google Scholar] [CrossRef]
- Perrenoud, A.G.-G.; Boccard, J.; Veuthey, J.-L.; Guillarme, D. Analysis of basic compounds by supercritical fluid chromatography: Attempts to improve peak shape and maintain mass spectrometry compatibility. J. Chromatogr. A 2012, 1262, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Li, J.R.; Li, M.; Xia, B.; Ding, L.S.; Xu, H.X.; Zhou, Y. Efficient optimization of ultra-high-performance supercritical fluid chromatographic separation of Rosa sericea by response surface methodology. J. Sep. Sci. 2013, 36, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Gao, B.; Zhang, X.; Xu, Y.; Shi, H.; Yu, L.L. Chemical profiling of triacylglycerols and diacylglycerols in cow milk fat by ultra-performance convergence chromatography combined with a quadrupole time-of-flight mass spectrometry. Food Chem. 2014, 143, 199–204. [Google Scholar] [CrossRef]
- Xu, J.; Chen, D.; Yan, X.; Chen, J.; Zhou, C. Global characterization of the photosynthetic glycerolipids from a marine diatom Stephanodiscus sp. by ultra performance liquid chromatography coupled with electrospray ionization-quadrupole-time of flight mass spectrometry. Anal. Chim. Acta. 2010, 663, 60–68. [Google Scholar] [CrossRef]
- Zhang, X.; Qi, C.; Zhang, Y.; Wei, W.; Jin, Q.; Xu, Z.; Tao, G.; Wang, X. Identification and quantification of triacylglycerols in human milk fat using ultra-performance convergence chromatography and quadrupole time-of-flight mass spectrometery with supercritical carbon dioxide as a mobile phase. Food Chem. 2019, 275, 712–720. [Google Scholar] [CrossRef]
- Gao, B.; Luo, Y.; Lu, W.; Liu, J.; Zhang, Y.; Yu, L.L. Triacylglycerol compositions of sunflower, corn and soybean oils examined with supercritical CO2 ultra-performance convergence chromatography combined with quadrupole time-of-flight mass spectrometry. Food Chem. 2017, 218, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhang, Y.; Yuan, F.; Gao, B.; Wang, Z.; Yu, L. Triacylglycerols composition analysis of olive oils by ultra-performance convergence chromatography combined with quadrupole time-of-flight mass spectrometry. Int. J. Food Sci. Technol. 2019, 54, 871–879. [Google Scholar] [CrossRef]
- Floch, J. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Ali, A.H.; Abd El-Wahed, E.M.; Abed, S.M.; Korma, S.A.; Wei, W.; Wang, X. Analysis of triacylglycerols molecular species composition, total fatty acids, and sn-2 fatty acids positional distribution in different types of milk powders. J. Food Meas. Charact. 2019, 13, 2613–2625. [Google Scholar] [CrossRef]
- Zheng, L.; Jin, J.; Huang, J.; Wang, Y.; Korma, S.A.; Wang, X.; Jin, Q. Effects of heat pretreatment of wet-milled corn germ on the physicochemical properties of oil. J. Food Sci. Technol. 2018, 55, 3154–3162. [Google Scholar] [CrossRef]
- Abed, S.M.; Zou, X.; Ali, A.H.; Jin, Q.; Wang, X. Profiling of triacylglycerol composition in arachidonic acid single cell oil from Mortierella alpina by using ultra-performance liquid chromatography-electrospray ionization-quadrupole-time-of-flight mass spectrometry. J. Food Compos. Anal. 2017, 62, 245–253. [Google Scholar] [CrossRef]
- Park, Y.; Juárez, M.; Ramos, M.; Haenlein, G. Physico-chemical characteristics of goat and sheep milk. Small Rumin. Res. 2007, 68, 88–113. [Google Scholar] [CrossRef] [Green Version]
- Castro-Gómez, M.; Rodriguez-Alcalá, L.M.; Calvo, M.V.; Romero, J.; Mendiola, J.; Ibañez, E.; Fontecha, J. Total milk fat extraction and quantification of polar and neutral lipids of cow, goat, and ewe milk by using a pressurized liquid system and chromatographic techniques. J. Dairy Sci. 2014, 97, 6719–6728. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.-Q.; Huang, J.-H.; Jin, Q.-Z.; Guo, Z.; Liu, Y.-F.; Cheong, L.-Z.; Xu, X.-B.; Wang, X.-G. Model for human milk fat substitute evaluation based on triacylglycerol composition profile. J. Agric. Food Chem. 2013, 61, 167–175. [Google Scholar] [CrossRef]
- Lunn, J.; Theobald, H.E. The health effects of dietary unsaturated fatty acids. Nutr. Bull. 2010, 32, 82–84. [Google Scholar] [CrossRef]
- Sun, C.; Zou, X.; Yao, Y.; Jin, J.; Xia, Y.; Huang, J.; Jin, Q.; Wang, X. Evaluation of fatty acid composition in commercial infant formulas on the Chinese market: A comparative study based on fat source and stage. Int. Dairy J. 2016, 63, 42–51. [Google Scholar] [CrossRef]
- Bakry, I.A.; Korma, S.A.; Wei, W.; Nafea, A.E.; Mahdi, A.A.; Ziedan, N.I.; Wang, X.G. Changes in the fatty acid content of Egyptian human milk across the lactation stages and in comparison with Chinese human milk. Eur. Food Res. Technol. 2021, 247, 1035–1048. [Google Scholar] [CrossRef]
- Koletzko, B.; Baker, S.; Cleghorn, G.; Neto, U.F.; Gopalan, S.; Hernell, O.; Hock, Q.S.; Jirapinyo, P.; Lonnerdal, B.; Pencharz, P. Global standard for the composition of infant formula: Recommendations of an ESPGHAN coordinated international expert group. J. Pediatr. Gastroenterol. Nutr. 2005, 41, 584–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Wang, T.; Liu, R.; Chang, M.; Jin, Q.; Wang, X. Chemical characterization of fourteen kinds of novel edible oils: A comparative study using chemometrics. LWT-Food Sci. Technol. 2020, 118, 108725. [Google Scholar] [CrossRef]
- Sawaya, W.; Safi, W.; Al-Shalhat, A.; Al-Mohammad, M. Chemical composition and nutritive value of goat milk. J. Dairy Sci. 1984, 67, 1655–1659. [Google Scholar] [CrossRef]
- Kompan, D.; Komprej, A. The effect of fatty acids in goat milk on health. In Milk Production-an Up-to-Date Overview of Animal Nutrition, Management and Health; IntechOpen: London, UK, 2012; pp. 3–28. [Google Scholar]
- Korma, S.A.; Zou, X.; Ali, A.H.; Abed, S.M.; Jin, Q.; Wang, X. Preparation of structured lipids enriched with medium-and long-chain triacylglycerols by enzymatic interesterification for infant formula. Food Bioprod. Process. 2018, 107, 121–130. [Google Scholar] [CrossRef]
- Prosser, C.; Svetashev, V.; Vyssotski, M.; Lowry, D. Composition and distribution of fatty acids in triglycerides from goat infant formulas with milk fat. J. Dairy Sci. 2010, 93, 2857–2862. [Google Scholar] [CrossRef] [Green Version]
- Nagachinta, S.; Akoh, C.C. Synthesis of structured lipid enriched with omega fatty acids and sn-2 palmitic acid by enzymatic esterification and its incorporation in powdered infant formula. J. Agric. Food Chem. 2013, 61, 4455–4463. [Google Scholar] [CrossRef]
- Sun, C.; Wei, W.; Zou, X.; Huang, J.; Jin, Q.; Wang, X. Evaluation of triacylglycerol composition in commercial infant formulas on the Chinese market: A comparative study based on fat source and stage. Food Chem. 2018, 252, 154–162. [Google Scholar] [CrossRef]
- Haddad, I.; Mozzon, M.; Frega, N.G. Trends in fatty acids positional distribution in human colostrum, transitional, and mature milk. Eur. Food Res. Technol. 2012, 235, 325–332. [Google Scholar] [CrossRef]
- Sun, C.; Wei, W.; Su, H.; Zou, X.; Wang, X. Evaluation of sn-2 fatty acid composition in commercial infant formulas on the Chinese market: A comparative study based on fat source and stage. Food Chem. 2018, 242, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Stergiadis, S.; Nørskov, N.P.; Purup, S. Comparative Nutrient Profiling of Retail Goat and Cow Milk. Nutrients 2019, 11, 2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Gutiérrez, V.; Barron, L.J. Methods for the analysis of triacylglycerols. J. Chromatogr. B Biomed. Sci. Appl. 1995, 671, 133–168. [Google Scholar] [CrossRef]
- Mottram, H.R.; Woodbury, S.E.; Evershed, R.P. Identification of triacylglycerol positional isomers present in vegetable oils by high performance liquid chromatography/atmospheric pressure chemical ionization mass spectrometry. Rapid Commun. Mass Spectrom. 1997, 11, 1240–1252. [Google Scholar] [CrossRef]
- Mottram, H.R.; Evershed, R.P. Elucidation of the composition of bovine milk fat triacylglycerols using high-performance liquid chromatography–atmospheric pressure chemical ionisation mass spectrometry. J. Chromatogr. A 2001, 926, 239–253. [Google Scholar] [CrossRef]
- Lísa, M.; Velínská, H.; Holcapek, M. Regioisomeric characterization of triacylglycerols using silver-ion HPLC/MS and randomization synthesis of standards. Anal. Chem. 2009, 81, 3903–3910. [Google Scholar] [CrossRef]
- Fontecha, J.; Diaz, V.; Fraga, M.; Juárez, M. Triglyceride analysis by gas chromatography in assessment of authenticity of goat milk fat. J. Am. Oil Chem. Soc. 1998, 75, 1893–1896. [Google Scholar] [CrossRef]
- Smiddy, M.A.; Huppertz, T.; van Ruth, S.M. Triacylglycerol and melting profiles of milk fat from several species. Int. Dairy J. 2012, 24, 64–69. [Google Scholar] [CrossRef]
- Yao, Y.; Zhao, G.; Xiang, J.; Zou, X.; Jin, Q.; Wang, X. Lipid composition and structural characteristics of bovine, caprine and human milk fat globules. Int. Dairy J. 2016, 56, 64–73. [Google Scholar] [CrossRef]
- Beccaria, M.; Sullini, G.; Cacciola, F.; Donato, P.; Dugo, P.; Mondello, L. High performance characterization of triacylglycerols in milk and milk-related samples by liquid chromatography and mass spectrometry. J. Chromatogr. A 2014, 1360, 172–187. [Google Scholar] [CrossRef]
- Riskó, T.C.; Csapó, Z. Goat Keeping and Goat Milk Products in Human Nutrition-Review. Appl. Stud. Agribusiness Commerce 2019, 13, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, D.; Dell’Orto, V.; Varisco, G.; Agazzi, A.; Savoini, G. Enrichment in n-3 fatty acids of goat’s colostrum and milk by maternal fish oil supplementation. Small Rumin. Res. 2006, 64, 22–29. [Google Scholar] [CrossRef]
- Kallio, H.; Nylund, M.; Boström, P.; Yang, B. Triacylglycerol regioisomers in human milk resolved with an algorithmic novel electrospray ionization tandem mass spectrometry method. Food Chem. 2017, 233, 351–360. [Google Scholar] [CrossRef]
- Wei, W.; Feng, Y.; Zhang, X.; Cao, X.; Feng, F. Synthesis of structured lipid 1, 3-dioleoyl-2-palmitoylglycerol in both solvent and solvent-free system. LWT Food Sci. Technol. 2015, 60, 1187–1194. [Google Scholar] [CrossRef]
- Rybak, O. Milk fat in structure formation of dairy products: A review. Ukr. Food J. 2016, 5, 499–514. [Google Scholar] [CrossRef]
- Sbihi, H.M.; Nehdi, I.A.; Tan, C.P.; Al-Resayes, S.I. Characteristics and fatty acid composition of milk fat from Saudi Aradi goat. Grasas Aceites 2015, 66, e101. [Google Scholar] [CrossRef] [Green Version]
- Rohman, A.; Gupitasari, I.; Purwanto, P.; Triyana, K.; Rosman, A.S.; Ahmad, S.A.S.; Yusof, F.M. Quantification of lard in the mixture with olive oil in cream cosmetics based on FTIR spectra and chemometrics for halal authentication. J. Teknol. 2014, 69, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Nicolaou, N.; Xu, Y.; Goodacre, R. Fourier transform infrared spectroscopy and multivariate analysis for the detection and quantification of different milk species. J. Dairy Sci. 2010, 93, 5651–5660. [Google Scholar] [CrossRef]
- Lu, H. A comparative study of storage stability in virgin coconut oil and extra virgin olive oil upon thermal treatment. Int. Food Res. J. 2009, 16, 343–354. [Google Scholar]
- Guillen, M.D.; Cabo, N. Study of the effects of smoke flavourings on the oxidative stability of the lipids of pork adipose tissue by means of Fourier transform infrared spectroscopy. Meat Sci. 2004, 66, 647–657. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | GZG * | XSG ** | ||||
|---|---|---|---|---|---|---|
| Total FAs | sn-2 FAs | sn-1, 3 FAs ¥ | Total FAs | sn-2 FAs | sn-1, 3 FAs ¥ | |
| C4:0 | 1.05 ± 0.04 a | ND | 1.58 ± 0.06 a˗ | 1.30 ± 0.16 a | ND | 1.95 ± 0.24 a˗ |
| C6:0 | 1.77 ± 0.16 a | ND | 2.66 ± 0.24 a˗ | 1.86 ± 0.08 a | ND | 2.79 ± 0.12 a˗ |
| C8:0 | 2.44 ± 0.08 a | 1.23 ± 0.08 A | 3.05 ± 0.08 a˗ | 2.48 ± 0.01 a | 1.10 ± 0.08 A | 3.17 ± 0.03 a˗ |
| C10:0 | 10.36 ± 0.16 a | 9.77 ± 0.82 A | 10.66 ± 0.16 a˗ | 9.48 ± 0.09 b | 7.88 ± 0.01 B | 10.28 ± 0.14 a˗ |
| C11:0 | 0.08 ± 0.03 a | ND | 0.13 ± 0.04 a˗ | 0.09 ± 0.01 a | ND | 0.14 ± 0.01 a˗ |
| C12:0 | 4.67 ± 0.24 a | 5.83 ± 0.08 A | 4.09 ± 0.33 a˗ | 3.72 ± 0.02 b | 5.71 ± 0.02 A | 2.73 ± 0.03 b˗ |
| C13:0 | 0.40 ± 0.02 a | 0.71 ± 0.02 A | 0.25 ± 0.01 a˗ | 0.07 ± 0.01 b | 0.09 ± 0.01 B | 0.06 ± 0.01 b˗ |
| C14:0 | 11.99 ± 0.82 a | 18.76 ± 0.98 A | 8.61 ± 0.73 a˗ | 9.75 ± 0.09 b | 16.98 ± 0.82 A | 6.14 ± 0.27 b˗ |
| C14:1 n-5 c | 0.38 ± 0.02 a | 1.12 ± 0.08 | 0.01 ± 0.00 b˗ | 0.19 ± 0.01 b | ND | 0.29 ± 0.01 a˗ |
| C15:0 | 1.09 ± 0.04 a | 1.20 ± 0.16 A | 1.04 ± 0.02 a˗ | 0.70 ± 0.16 b | 0.97 ± 0.02 A | 0.57 ± 0.25 a˗ |
| C15:1 n-5 c | 0.48 ± 0.02 a | 0.61 ± 0.09 | 0.42 ± 0.02 a˗ | 0.24 ± 0.10 b | ND | 0.36 ± 0.01 b˗ |
| C16:0 | 31.40 ± 0.82 a | 34.11 ± 0.10 A | 30.05 ± 0.78 a˗ | 23.84 ± 0.82 b | 31.65 ± 0.80 B | 19.94 ± 0.82 b˗ |
| C16:1 n-7 c | 1.33 ± 0.02 a | 1.17 ± 0.02 A | 1.41 ± 0.03 a˗ | 1.11 ± 0.01 b | 1.14 ± 0.02 A | 1.09 ± 0.02 b˗ |
| C17:0 | 0.72 ± 0.03 b | 0.59 ± 0.02 B | 0.79 ± 0.04 b˗ | 0.98 ± 0.02 a | 1.09 ± 0.01 A | 0.93 ± 0.03 a˗ |
| C17:1 n-7 t | 0.41 ± 0.01 b | 0.55 ± 0.03 B | 0.35 ± 0.00 b˗ | 0.54 ± 0.02 a | 0.81 ± 0.08 A | 0.41 ± 0.02 a˗ |
| C18:0 | 6.61 ± 0.03 b | 5.86 ± 0.02 B | 6.99 ± 0.04 b˗ | 13.49 ± 0.82 a | 9.65 ± 0.09 A | 15.41 ± 1.27 a˗ |
| C18:1 n-9 c | 21.79 ± 0.98 b | 15.87 ± 0.82 B | 24.75 ± 1.06 b˗ | 27.18 ± 0.81 a | 20.86 ± 0.09 A | 30.34 ± 1.17 a˗ |
| C18:2 n-6 c | 2.54 ± 0.24 a | 2.69 ± 0.01 A | 2.47 ± 0.36 a˗ | 1.90 ± 0.08 b | 1.83 ± 0.02 B | 1.94 ± 0.11 a˗ |
| C18:3 n-6 c | ND | ND | ND | 0.30 ± 0.08 | ND | 0.45 ± 0.12 |
| C18:3 n-3 c | 0.64 ± 0.01 a | ND | 0.96 ± 0.01 a˗ | 0.32 ± 0.02 b | ND | 0.48 ± 0.02 b˗ |
| C20:0 | ND | ND | ND | 0.13 ± 0.01 | ND | 0.20 ± 0.01 |
| C20:4 n-6 c | 0.25 ± 0.02 a | ND | 0.38 ± 0.02 a˗ | 0.23 ± 0.02 a | ND | 0.35 ± 0.02 a˗ |
| Sums and ratios of FAs | ||||||
| ΣSC-SFAs | 2.82 ± 0.20 a | ND | 4.24 ± 0.31 a˗ | 3.16 ± 0.24 a | ND | 4.74 ± 0.37 a˗ |
| ΣMC-SFAs | 29.94 ± 1.35 a | 36.30 ± 1.98 A | 26.78 ± 1.03 a˗ | 25.59 ± 0.22 b | 31.76 ± 0.88 B | 22.51 ± 0.11 b˗ |
| ΣLC-SFAs | 39.82 ± 0.92 a | 41.76 ± 1.11 A | 38.86 ± 0.83 a˗ | 39.14 ± 0.16 a | 43.36 ± 0.88 A | 37.04 ± 0.21 b˗ |
| ΣSFAs | 72.58 ± 2.48 a | 78.06 ± 3.09 A | 69.88 ± 2.17 a˗ | 67.89 ± 0.62 a | 75.12 ± 1.76 A | 64.29 ± 0.05 b˗ |
| ΣUFAs | 27.82 ± 1.33 b | 22.01 ± 1.05 B | 30.74 ± 1.47 b˗ | 32.01 ± 0.96 a | 24.64 ± 0.20 A | 35.70 ± 1.34 a˗ |
| ΣMUFAs | 24.39 ± 1.06 b | 19.32 ± 1.04 B | 26.93 ± 1.07 b˗ | 29.26 ± 0.83 a | 22.81 ± 0.19 A | 32.49 ± 1.16 a˗ |
| ΣPUFAs | 3.43 ± 0.27 a | 2.69 ± 0.01 A | 3.80 ± 0.40 a˗ | 2.75 ± 0.13 b | 1.83 ± 0.02 B | 3.21 ± 0.19 a˗ |
| ΣOCS-FAs | 1.81 ± 0.07 a | 1.79 ± 0.19 A | 1.82 ± 0.02 a˗ | 1.68 ± 0.15 a | 2.06 ± 0.01 A | 1.49 ± 0.22 a˗ |
| ΣECS-FAs | 50.00 ± 1.67 a | 58.73 ± 1.90 A | 45.64 ± 1.55 a˗ | 47.08 ± 0.09 a | 58.28 ± 1.70 A | 41.48 ± 0.72 b˗ |
| Σn-6 | 2.79 ± 0.26 a | 2.69 ± 0.01 A | 2.84 ± 0.39 a˗ | 2.43 ± 0.15 a | 1.83 ± 0.02 B | 2.73 ± 0.21 a˗ |
| Σn-3 | 0.64 ± 0.01 a | ND | 0.96 ± 0.01 a˗ | 0.32 ± 0.02 b | ND | 0.48 ± 0.02 b˗ |
| n-6/n-3 | 4.35 ± 0.35 b | ND | 2.96 ± 0.37 b˗ | 7.64 ± 0.85 a | ND | 5.73 ± 0.74 a˗ |
| L/Ln | 3.96 ± 0.33 b | ND | 2.57 ± 0.35 b˗ | 5.97 ± 0.56 a | ND | 4.06 ± 0.45 a˗ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korma, S.A.; Li, L.; Wei, W.; Liu, P.; Zhang, X.; Bakry, I.A.; An, P.; Abdrabo, K.A.E.; Manzoor, M.F.; Umair, M.; et al. A Comparative Study of Milk Fat Extracted from the Milk of Different Goat Breeds in China: Fatty Acids, Triacylglycerols and Thermal and Spectroscopic Characterization. Biomolecules 2022, 12, 730. https://doi.org/10.3390/biom12050730
Korma SA, Li L, Wei W, Liu P, Zhang X, Bakry IA, An P, Abdrabo KAE, Manzoor MF, Umair M, et al. A Comparative Study of Milk Fat Extracted from the Milk of Different Goat Breeds in China: Fatty Acids, Triacylglycerols and Thermal and Spectroscopic Characterization. Biomolecules. 2022; 12(5):730. https://doi.org/10.3390/biom12050730
Chicago/Turabian StyleKorma, Sameh A., Li Li, Wei Wei, Pengzhan Liu, Xinghe Zhang, Ibrahim A. Bakry, Peipei An, Khaled A. E. Abdrabo, Muhammad Faisal Manzoor, Muhammad Umair, and et al. 2022. "A Comparative Study of Milk Fat Extracted from the Milk of Different Goat Breeds in China: Fatty Acids, Triacylglycerols and Thermal and Spectroscopic Characterization" Biomolecules 12, no. 5: 730. https://doi.org/10.3390/biom12050730
APA StyleKorma, S. A., Li, L., Wei, W., Liu, P., Zhang, X., Bakry, I. A., An, P., Abdrabo, K. A. E., Manzoor, M. F., Umair, M., Cacciotti, I., Lorenzo, J. M., & Conte-Junior, C. A. (2022). A Comparative Study of Milk Fat Extracted from the Milk of Different Goat Breeds in China: Fatty Acids, Triacylglycerols and Thermal and Spectroscopic Characterization. Biomolecules, 12(5), 730. https://doi.org/10.3390/biom12050730