
N-Palmitoyl-D-Glucosamine Inhibits TLR-4/NLRP3 and Improves DNBS-Induced Colon Inflammation through a PPAR-α-Dependent Mechanism

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Disease Activity Index (DAI)
2.3. Histopathological Analyses
2.4. Immunofluorescence Analysis on Colonic Sections
2.5. Enzyme-Linked Immunosorbent Assay (ELISA) for IL-1β and PGE2
2.6. Statistical Analyses
3. Results
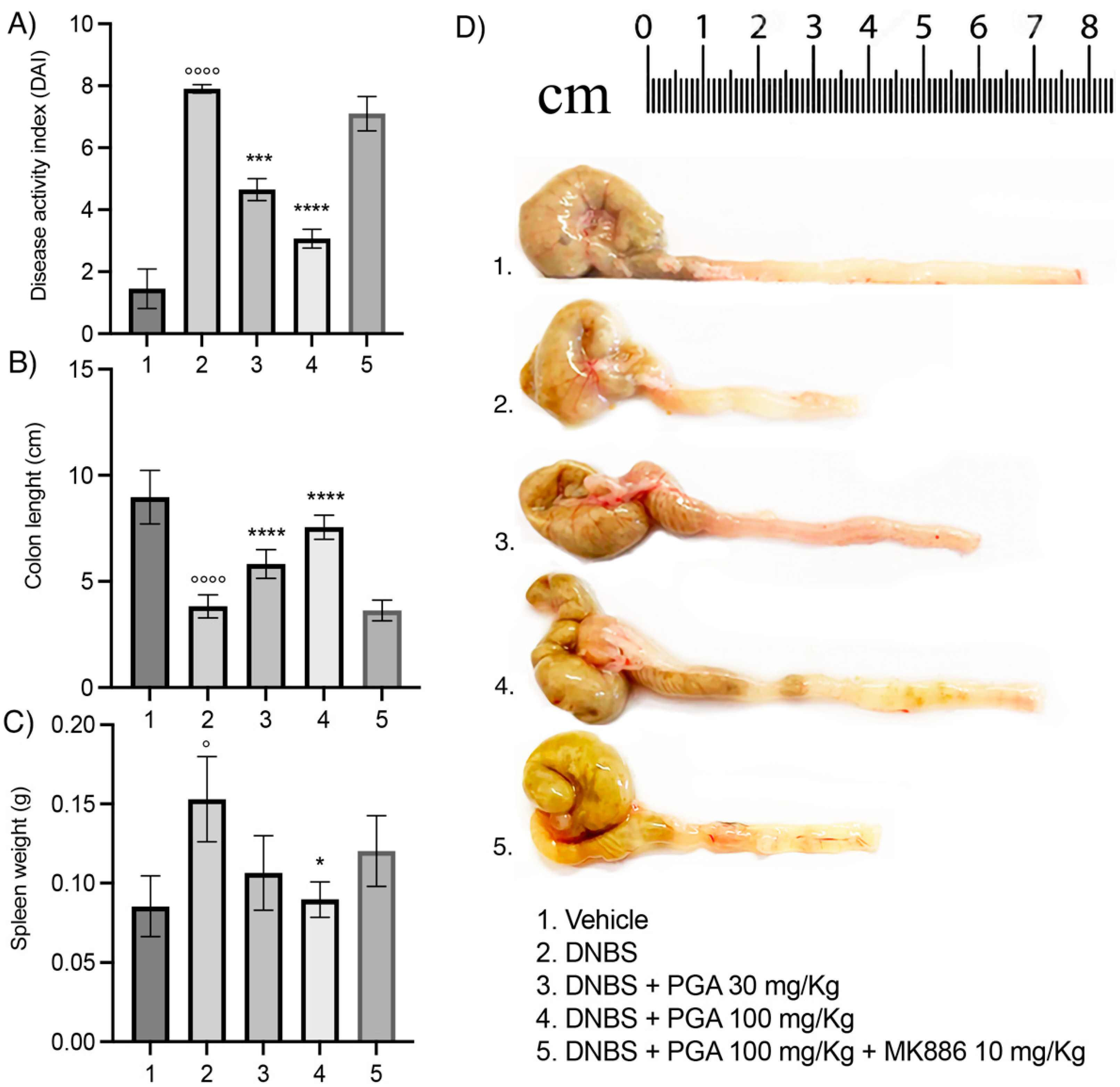
3.1. Micronized PGA Improves the Disease Spectrum and Macroscopic Signs of Colitis in a Dose-Dependent Manner through PPAR-α Involvement
3.2. Micronized PGA Ameliorates Mucosal Integrity and Prevents Colonic Histological Damage in DNBS-Treated Mice
3.3. Micronized PGA Downregulates the TLR-4/NLRP3/iNOS Expression and Decreases the Release of Inflammatory Mediators in DNBS-Treated Mice via PPAR-α Receptors
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.-Z.; Li, Y.-Y. Inflammatory bowel disease: Pathogenesis. World J. Gastroenterol. 2014, 20, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Siel, D.; Beltrán, C.J.; Martínez, E.; Pino, M.; Vargas, N.; Salinas, A.; Pérez, O.; Pereira, I.; Ramírez-Toloza, G. Elucidating the Role of Innate and Adaptive Immune Responses in the Pathogenesis of Canine Chronic Inflammatory Enteropathy—A Search for Potential Biomarkers. Animals 2022, 12, 1645. [Google Scholar] [CrossRef] [PubMed]
- Kucharzik, T.; Koletzko, S.; Kannengießer, K.; Dignaß, A. Ulcerative Colitis—Diagnostic and Therapeutic Algorithms. Dtsch. Ärzteblatt Int. 2020, 117, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Ha, C. Epidemiology and Pathogenesis of Ulcerative Colitis. Gastroenterol. Clin. N. Am. 2020, 49, 643–654. [Google Scholar] [CrossRef]
- Gearry, R.B.; Barclay, M.L.; Burt, M.J.; Collett, J.A.; Chapman, B.A. Thiopurine drug adverse effects in a population of New Zealand patients with inflammatory bowel disease. Pharmacoepidemiol. Drug Saf. 2004, 13, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Siegmund, B.; Le Berre, C.; Wei, S.C.; Ferrante, M.; Shen, B.; Bernstein, C.N.; Danese, S.; Peyrin-Biroulet, L.; Hibi, T. Ulcerative colitis. Nat. Rev. Dis. Prim. 2020, 6, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Siracusa, R.; Impellizzeri, D.; D’Amico, R.; Peritore, A.F.; Crupi, R.; Gugliandolo, E.; Fusco, R.; Di Paola, R.; Schievano, C.; et al. Safety and efficacy of a new micronized formulation of the ALIAmide palmitoylglucosamine in preclinical models of inflammation and osteoarthritis pain. Arthritis Res. Ther. 2019, 21, 254. [Google Scholar] [CrossRef] [Green Version]
- Gugliandolo, E.; Peritore, A.F.; Piras, C.; Cuzzocrea, S.; Crupi, R. Palmitoylethanolamide and Related ALIAmides: Prohomeostatic Lipid Compounds for Animal Health and Wellbeing. Veter.-Sci. 2020, 7, 78. [Google Scholar] [CrossRef]
- Capasso, R.; Orlando, P.; Pagano, E.; Aveta, T.; Buono, L.; Borrelli, F.; Di Marzo, V.; Izzo, A.A. Palmitoylethanolamide normalizes intestinal motility in a model of post-inflammatory accelerated transit: Involvement of CB1 receptors and TRPV1 channels. J. Cereb. Blood Flow Metab. 2014, 171, 4026–4037. [Google Scholar] [CrossRef] [Green Version]
- Borrelli, F.; Romano, B.; Petrosino, S.; Pagano, E.; Capasso, R.; Coppola, D.; Battista, G.; Orlando, P.; Di Marzo, V.; Izzo, A.A. Palmitoylethanolamide, a naturally occurring lipid, is an orally effective intestinal anti-inflammatory agent. Br. J. Pharmacol. 2014, 172, 142–158. [Google Scholar] [CrossRef] [Green Version]
- Yomogida, S.; Kojima, Y.; Tsutsumi-Ishii, Y.; Hua, J.; Sakamoto, K.; Nagaoka, I. Glucosamine, a naturally oc-curring amino monosaccharide, suppresses dextran sulfate sodium-induced colitis in rats. Int. J. Mol. Med. 2008, 22, 317–323. [Google Scholar] [PubMed]
- Bak, Y.-K.; Lampe, J.W.; Sung, M.-K. Effects of dietary supplementation of glucosamine sulfate on intestinal inflammation in a mouse model of experimental colitis. J. Gastroenterol. Hepatol. 2014, 29, 957–963. [Google Scholar] [CrossRef]
- Navarro, S.L.; Levy, L.; Curtis, K.R.; Lampe, J.W.; Hullar, M.A.J. Modulation of Gut Microbiota by Glucosamine and Chondroitin in a Randomized, Double-Blind Pilot Trial in Humans. Microorganisms 2019, 7, 610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannotta, M.; Belardo, C.; Trotta, M.C.; Iannotti, F.A.; Vitale, R.M.; Maisto, R.; Boccella, S.; Infantino, R.; Ricciardi, F.; Mirto, B.F.; et al. N-palmitoyl-D-glucosamine, a Natural Monosaccharide-Based Glycolipid, Inhibits TLR4 and Prevents LPS-Induced Inflammation and Neuropathic Pain in Mice. Int. J. Mol. Sci. 2021, 22, 1491. [Google Scholar] [CrossRef] [PubMed]
- Del Re, A.; Corpetti, C.; Pesce, M.; Seguella, L.; Steardo, L.; Palenca, I.; Rurgo, S.; De Conno, B.; Sarnelli, G.; Esposito, G. Ultramicronized Palmitoylethanolamide Inhibits NLRP3 Inflammasome Expression and Pro-Inflammatory Response Activated by SARS-CoV-2 Spike Protein in Cultured Murine Alveolar Macrophages. Metabolites 2021, 11, 592. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Di Paola, R.; Cordaro, M.; Gugliandolo, E.; Casili, G.; Morittu, V.M.; Britti, D.; Esposito, E.; Cuzzocrea, S. Adelmidrol, a palmitoylethanolamide analogue, as a new pharmacological treatment for the management of acute and chronic inflammation. Biochem. Pharmacol. 2016, 119, 27–41. [Google Scholar] [CrossRef]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Laboratory investigation. J. Tech. Methods Pathol. 1993, 69, 238–249. [Google Scholar]
- Li, R.; Kim, M.-H.; Sandhu, A.K.; Gao, C.; Gu, L. Muscadine Grape (Vitis rotundifolia) or Wine Phytochemicals Reduce Intestinal Inflammation in Mice with Dextran Sulfate Sodium-Induced Colitis. J. Agric. Food Chem. 2017, 65, 769–776. [Google Scholar] [CrossRef]
- Cushing, K.; Higgins, P.D.R. Management of Crohn Disease. JAMA 2021, 325, 69–80. [Google Scholar] [CrossRef]
- Bruscoli, S.; Febo, M.; Riccardi, C.; Migliorati, G. Glucocorticoid Therapy in Inflammatory Bowel Disease: Mechanisms and Clinical Practice. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Shen, W.; Gao, Y.; Lu, B.; Zhang, Q.; Hu, Y.; Chen, Y. Negatively regulating TLR4/NF-κB signaling via PPARα in endotoxin-induced uveitis. Biochim. Et Biophys. Acta (BBA)—Mol. Basis Dis. 2014, 1842, 1109–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couch, D.G.; Cook, H.; Ortori, C.; Barrett, D.; Lund, J.N.; O’Sullivan, S.E. Palmitoylethanolamide and Cannabidiol Prevent Inflammation-induced Hyperpermeability of the Human Gut In Vitro and In Vivo—A Randomized, Placebo-controlled, Double-blind Controlled Trial. Inflamm. Bowel Dis. 2019, 25, 1006–1018. [Google Scholar] [CrossRef] [PubMed]
- Matalon, S.T.; Azar, S.; Meiri, D.; Hadar, R.; Nemirovski, A.; Abu Jabal, N.; Konikoff, F.M.; Drucker, L.; Tam, J.; Naftali, T. Endocannabinoid Levels in Ulcerative Colitis Patients Correlate with Clinical Parameters and Are Affected by Cannabis Consumption. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef]
- Esposito, G.; Corpetti, C.; Pesce, M.; Seguella, L.; Annunziata, G.; Del Re, A.; Vincenzi, M.; Lattanzi, R.; Lu, J.; Sanseverino, W.; et al. A Palmitoylethanolamide Producing Lactobacillus paracasei Improves Clostridium difficile Toxin A-Induced Colitis. Front. Pharmacol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Pesce, M.; Seguella, L.; Lu, J.; Corpetti, C.; Del Re, A.; De Palma, F.; Esposito, G.; Sanseverino, W.; Sarnelli, G. Engineered Lactobacillus paracasei Producing Palmitoylethanolamide (PEA) Prevents Colitis in Mice. Int. J. Mol. Sci. 2021, 22, 2945. [Google Scholar] [CrossRef]
- Goodman, M.J.; Kent, P.W.; Truelove, S.C. Glucosamine synthetase activity of the colonic mucosa in ulcerative colitis and Crohn’s disease. Gut 1977, 18, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Goodman, M.J.; Kent, P.W.; Truelove, S.C. Glucosamine synthetase activity of the colonic mucosa in membranous colitis. Gut 1977, 18, 229–231. [Google Scholar] [CrossRef] [Green Version]
- Salvatore, S.; Heuschkel, R.; Tomlin, S.; Davies, S.E.; Edwards, S.; Walker-Smith, J.A.; French, I.; Murch, S.H. A pilot study of N-acetyl glucosamine, a nutritional substrate for glycosaminoglycan synthesis, in paediatric chronic inflammatory bowel disease. Aliment. Pharmacol. Ther. 2000, 14, 1567–1579. [Google Scholar] [CrossRef]
- Kantor, E.D.; Zhang, X.; Wu, K.; Signorello, L.B.; Chan, A.T.; Fuchs, C.S.; Giovannucci, E.L. Use of glucosamine and chondroitin supplements in relation to risk of colorectal cancer: Results from the Nurses’ Health Study and Health Professionals follow-up study. Int. J. Cancer 2016, 139, 1949–1957. [Google Scholar] [CrossRef]
- Kantor, E.D.; Newton, C.C.; Giovannucci, E.L.; McCullough, M.L.; Campbell, P.T.; Jacobs, E.J. Glucosamine use and risk of colorectal cancer: Results from the Cancer Prevention Study II Nutrition Cohort. Cancer Causes Control 2018, 29, 389–397. [Google Scholar] [CrossRef]
- Lee, D.H.; Cao, C.; Zong, X.; Zhang, X.; O’Connell, K.; Song, M.; Wu, K.; Du, M.; Cao, Y.; Giovannucci, E.L.; et al. Glucosamine and Chondroitin Supplements and Risk of Colorectal Adenoma and Serrated Polyp. Cancer Epidemiol. Biomark. Prev. 2020, 29, 2693–2701. [Google Scholar] [CrossRef] [PubMed]
- Navarro, S.L.; White, E.; Kantor, E.; Zhang, Y.; Rho, J.; Song, X.; Milne, G.; Lampe, P.D.; Lampe, J.W. Randomized Trial of Glucosamine and Chondroitin Supplementation on Inflammation and Oxidative Stress Biomarkers and Plasma Proteomics Profiles in Healthy Humans. PLoS ONE 2015, 10, e0117534. [Google Scholar] [CrossRef] [PubMed]
- Sarnelli, G.; Seguella, L.; Pesce, M.; Lu, J.; Gigli, S.; Bruzzese, E.; Lattanzi, R.; D’Alessandro, A.; Cuomo, R.; Steardo, L.; et al. HIV-1 Tat-induced diarrhea is improved by the PPARalpha agonist, palmitoylethanolamide, by suppressing the activation of enteric glia. J. Neuroinflamm. 2018, 15, 94. [Google Scholar] [CrossRef] [Green Version]
- Darmani, N.A.; Izzo, A.A.; Degenhardt, B.; Valenti, M.; Scaglione, G.; Capasso, R.; Sorrentini, I.; Di Marzo, V. Involvement of the cannabimimetic compound, N-palmitoyl-ethanolamine, in inflammatory and neuropathic conditions: Review of the available pre-clinical data, and first human studies. Neuropharmacology 2005, 48, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Grabacka, M.; Pierzchalska, M.; Płonka, P.M.; Pierzchalski, P. The Role of PPAR Alpha in the Modulation of Innate Immunity. Int. J. Mol. Sci. 2021, 22, 10545. [Google Scholar] [CrossRef] [PubMed]
- Dana, N.; Vaseghi, G.; Haghjooy-Javanmard, S. Crosstalk between Peroxisome Proliferator-Activated Receptors andToll-Like Receptors: A Systematic Review. Adv. Pharm. Bull. 2019, 9, 12–21. [Google Scholar] [CrossRef]
- Takeda, K.; Akira, S. Microbial recognition by Toll-like receptors. J. Dermatol. Sci. 2004, 34, 73–82. [Google Scholar] [CrossRef]
- Zhen, Y.; Zhang, H. NLRP3 Inflammasome and Inflammatory Bowel Disease. Front. Immunol. 2019, 10, 276. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.-Y.; Ye, X.-J.; He, X.-H.; Ouyang, D.-Y. The Signaling Pathways Regulating NLRP3 Inflammasome Activation. Inflammation 2021, 44, 1229–1245. [Google Scholar] [CrossRef]
- Zhuang, Y.; Zhao, F.; Liang, J.; Deng, X.; Zhang, Y.; Ding, G.; Zhang, A.; Jia, Z.; Huang, S. Activation of COX-2/mPGES-1/PGE2 Cascade via NLRP3 Inflammasome Contributes to Albumin-Induced Proximal Tubule Cell Injury. Cell. Physiol. Biochem. 2017, 42, 797–807. [Google Scholar] [CrossRef]
- Luo, H.; Guo, P.; Zhou, Q. Role of TLR4/NF-κB in Damage to Intestinal Mucosa Barrier Function and Bacterial Translocation in Rats Exposed to Hypoxia. PLoS ONE 2012, 7, e46291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Soto, W.; Gulbransen, B.D. Enteric Glia: A New Player in Abdominal Pain. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 433–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, B.; Tang, W.-H.; Lu, L.-J.; Zhou, Y.; Zhu, H.-Y.; Zhou, Y.-L.; Zhang, H.-H.; Hu, C.-Y.; Xu, G.-Y. TLR4 upregulates CBS expression through NF-κB activation in a rat model of irritable bowel syndrome with chronic visceral hypersensitivity. World J. Gastroenterol. 2015, 21, 8615–8628. [Google Scholar] [CrossRef]
- Bettoni, I.; Comelli, F.; Rossini, C.; Granucci, F.; Giagnoni, G.; Peri, F.; Costa, B. Glial TLR4 receptor as new target to treat neuropathic pain: Efficacy of a new receptor antagonist in a model of peripheral nerve injury in mice. Glia 2008, 56, 1312–1319. [Google Scholar] [CrossRef]
- Hutchinson, M.R.; Zhang, Y.; Brown, K.; Coats, B.D.; Shridhar, M.; Sholar, P.W.; Patel, S.J.; Crysdale, N.Y.; Harrison, J.A.; Maier, S.F. Non-stereoselective reversal of neuropathic pain by naloxone and naltrexone: Involvement of toll-like receptor 4 (TLR4). Eur. J. Neurosci. 2008, 28, 20–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lönnfors, S.; Vermeire, S.; Greco, M.; Hommes, D.; Bell, C.; Avedano, L. IBD and health-related quality of life—Discovering the true impact. J. Crohn’s Colitis 2014, 8, 1281–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuesta, C.M.; Pascual, M.; Pérez-Moraga, R.; Rodríguez-Navarro, I.; García-García, F.; Ureña-Peralta, J.R.; Guerri, C. TLR4 Deficiency Affects the Microbiome and Reduces Intestinal Dysfunctions and Inflammation in Chronic Alcohol-Fed Mice. Int. J. Mol. Sci. 2021, 22, 12830. [Google Scholar] [CrossRef] [PubMed]
- Philip-Hollingsworth, S.; Dazzo, F.B.; I Hollingsworth, R. Structural requirements of Rhizobium chitolipooligosaccharides for uptake and bioactivity in legume roots as revealed by synthetic analogs and fluorescent probes. J. Lipid Res. 1997, 38, 1229–1241. [Google Scholar] [CrossRef]
- Das, A.; Srinivasan, M.; Ghosh, T.S.; Mande, S.S. Xenobiotic metabolism and gut microbiomes. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Matuszyk, A.; Ceranowicz, D.; Warzecha, Z.; Ceranowicz, P.; Fyderek, K.; Gałązka, K.; Cieszkowski, J.; Bonior, J.; Jaworek, J.; Pihut, M.; et al. The Influence of Ghrelin on the Development of Dextran Sodium Sul-fate-Induced Colitis in Rats. BioMed Res. Int. 2015, 2015, 718314. [Google Scholar] [CrossRef] [Green Version]
- Dembiński, A.; Warzecha, Z.; Ceranowicz, P.; Dembiński, M.; Cieszkowski, J.; Gosiewski, T.; Bulanda, M.; Kuśnierz-Cabala, B.; Gałązka, K.; Konturek, P.C. Synergic Interaction of Rifaximin and Mutaflor (Escherichia coli Nissle 1917) in the Treatment of Acetic Acid-Induced Colitis in Rats. Gastroenterol. Res. Pract. 2016, 2016, 3126280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matuszyk, A.; Ceranowicz, P.; Warzecha, Z.; Cieszkowski, J.; Gałązka, K.; Bonior, J.; Jaworek, J.; Konturek, P.C.; Gil, K.; Dembiński, A. Pretreatment with obestatin inhibits the development of acetic acid-induced colitis in rats. Arch. Med. Sci. 2018, 14, 920–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Antibody | Host | Clonality | Dilution | Brand |
|---|---|---|---|---|
| ZO-1 | Mouse | Monoclonal | 6 microgram w/v | Invitrogen, Thermo Fisher, Waltham, MA, USA |
| Occludin | Rabbit | Polyclonal | 1:100 v/v | Bioss Antibodies, Boston, MA, USA |
| NLRP3 | Rabbit | Polyclonal | 1:1000 v/v | Invitrogen, Thermo Fisher, Waltham, MA, USA |
| TLR-4 | Rabbit | Polyclonal | 1:150 v/v | Bioss Antibodies, Boston, MA, USA |
| iNOS | Mouse | Monoclonal | 1:1000 v/v | Novusbio, Centennial, CO, USA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palenca, I.; Seguella, L.; Del Re, A.; Franzin, S.B.; Corpetti, C.; Pesce, M.; Rurgo, S.; Steardo, L.; Sarnelli, G.; Esposito, G. N-Palmitoyl-D-Glucosamine Inhibits TLR-4/NLRP3 and Improves DNBS-Induced Colon Inflammation through a PPAR-α-Dependent Mechanism. Biomolecules 2022, 12, 1163. https://doi.org/10.3390/biom12081163
Palenca I, Seguella L, Del Re A, Franzin SB, Corpetti C, Pesce M, Rurgo S, Steardo L, Sarnelli G, Esposito G. N-Palmitoyl-D-Glucosamine Inhibits TLR-4/NLRP3 and Improves DNBS-Induced Colon Inflammation through a PPAR-α-Dependent Mechanism. Biomolecules. 2022; 12(8):1163. https://doi.org/10.3390/biom12081163
Chicago/Turabian StylePalenca, Irene, Luisa Seguella, Alessandro Del Re, Silvia Basili Franzin, Chiara Corpetti, Marcella Pesce, Sara Rurgo, Luca Steardo, Giovanni Sarnelli, and Giuseppe Esposito. 2022. "N-Palmitoyl-D-Glucosamine Inhibits TLR-4/NLRP3 and Improves DNBS-Induced Colon Inflammation through a PPAR-α-Dependent Mechanism" Biomolecules 12, no. 8: 1163. https://doi.org/10.3390/biom12081163
APA StylePalenca, I., Seguella, L., Del Re, A., Franzin, S. B., Corpetti, C., Pesce, M., Rurgo, S., Steardo, L., Sarnelli, G., & Esposito, G. (2022). N-Palmitoyl-D-Glucosamine Inhibits TLR-4/NLRP3 and Improves DNBS-Induced Colon Inflammation through a PPAR-α-Dependent Mechanism. Biomolecules, 12(8), 1163. https://doi.org/10.3390/biom12081163