
NMR-Based Metabolomic Analysis for the Effects of Trimethylamine N-Oxide Treatment on C2C12 Myoblasts under Oxidative Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture and Experimental Design
2.3. MTS Cell Viability Assay and Cell Proliferation Determination
2.4. Measurement of Cellular T-SOD Capacity
2.5. Measurement of Cellular MDA Level
2.6. Western blotting
2.7. Extraction of Intracellular Aqueous Metabolites
2.8. Sample Preparation and NMR Measurements
2.9. NMR Data Preprocessing
2.10. NMR Data Analysis
3. Results
3.1. Establishment of the Oxidative Stress Model of C2C12 Cells
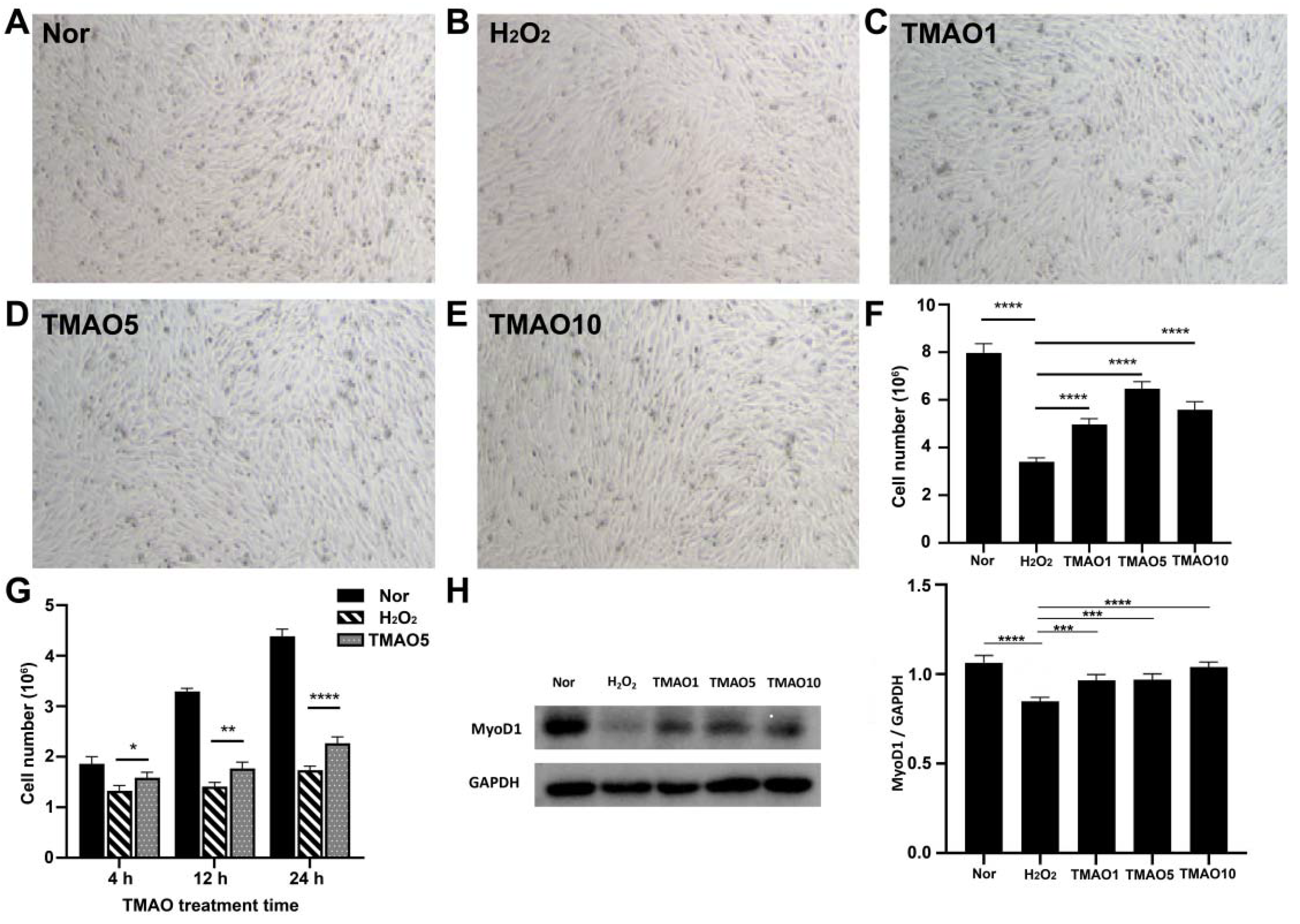
3.2. Effect of TMAO Treatment on the Proliferation Ability of C2C12 Cells
3.3. TMAO Treatment Partly Enhanced the Proliferation and Differentiation Potential of H2O2-Impaired C2C12 Cells
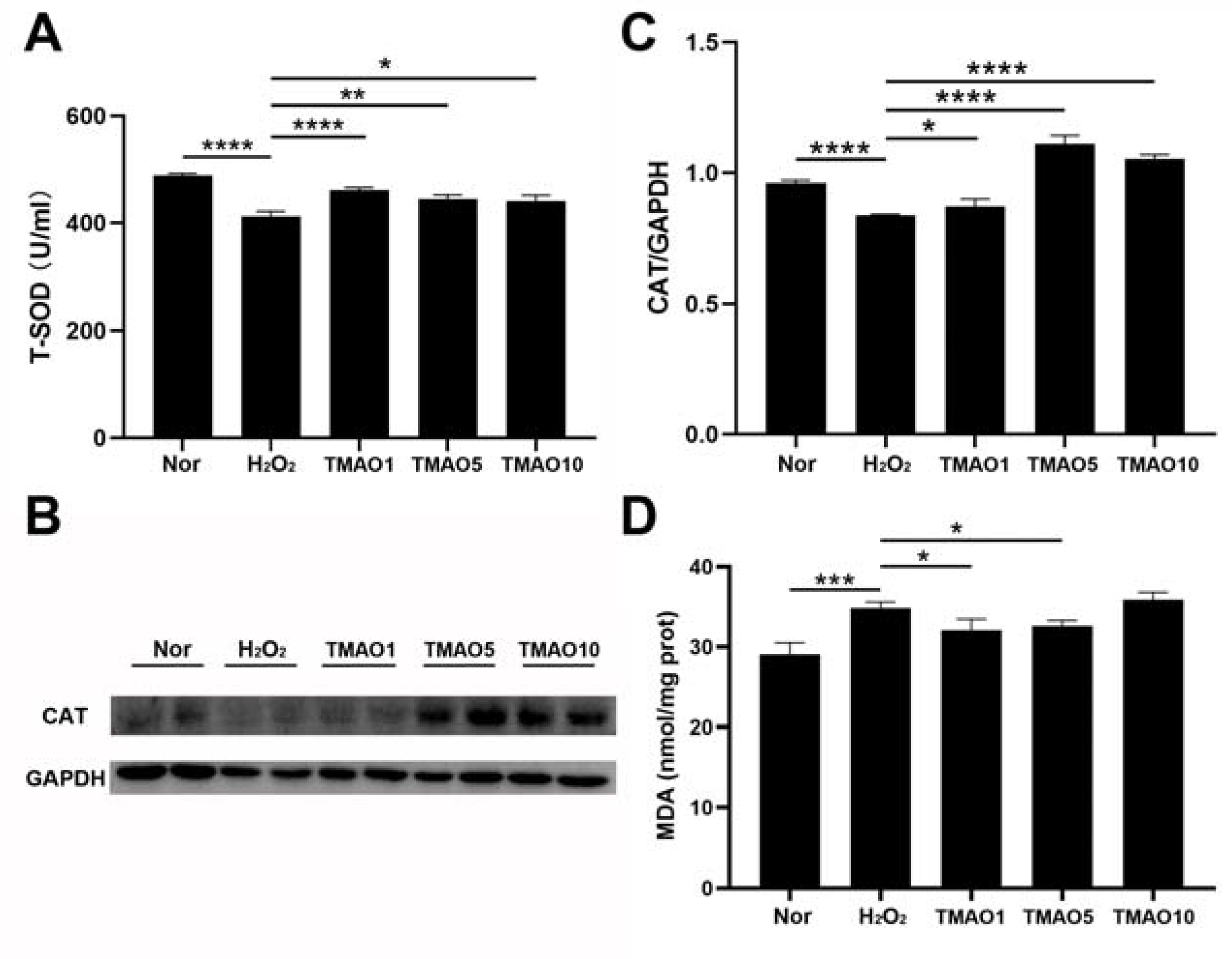
3.4. TMAO Treatment Enhanced Antioxidant Activities in H2O2-Impaired C2C12 Cells
3.5. TMAO Treatment Decreased the MDA Level of H2O2-Impaired C2C12 Cells
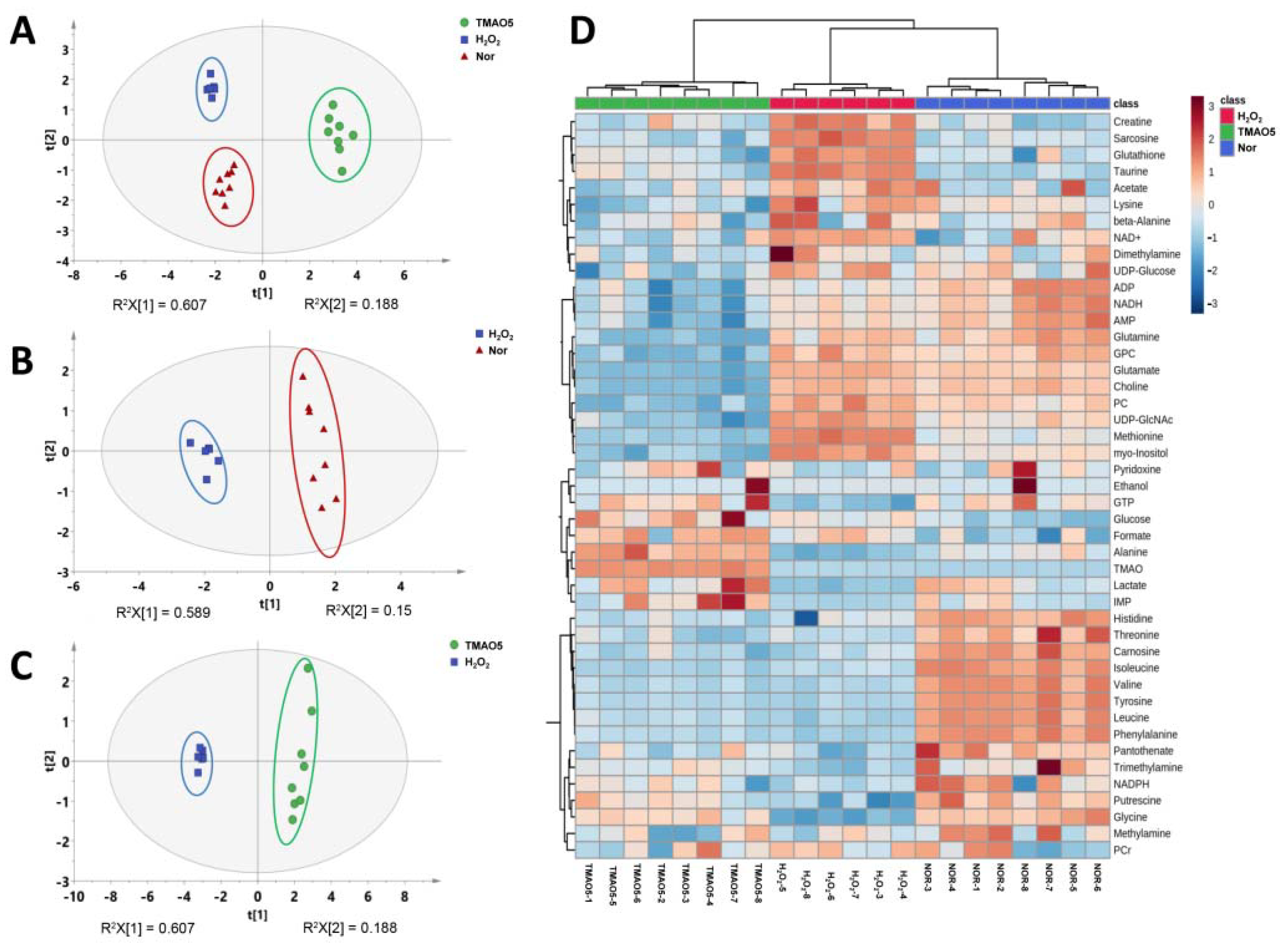
3.6. NMR Spectra of Aqueous Extracts Derived from C2C12 Myoblasts
3.7. Multivariate Data Analysis for NMR Data of C2C12 Myoblasts
3.8. Identifications of Differential Metabolites
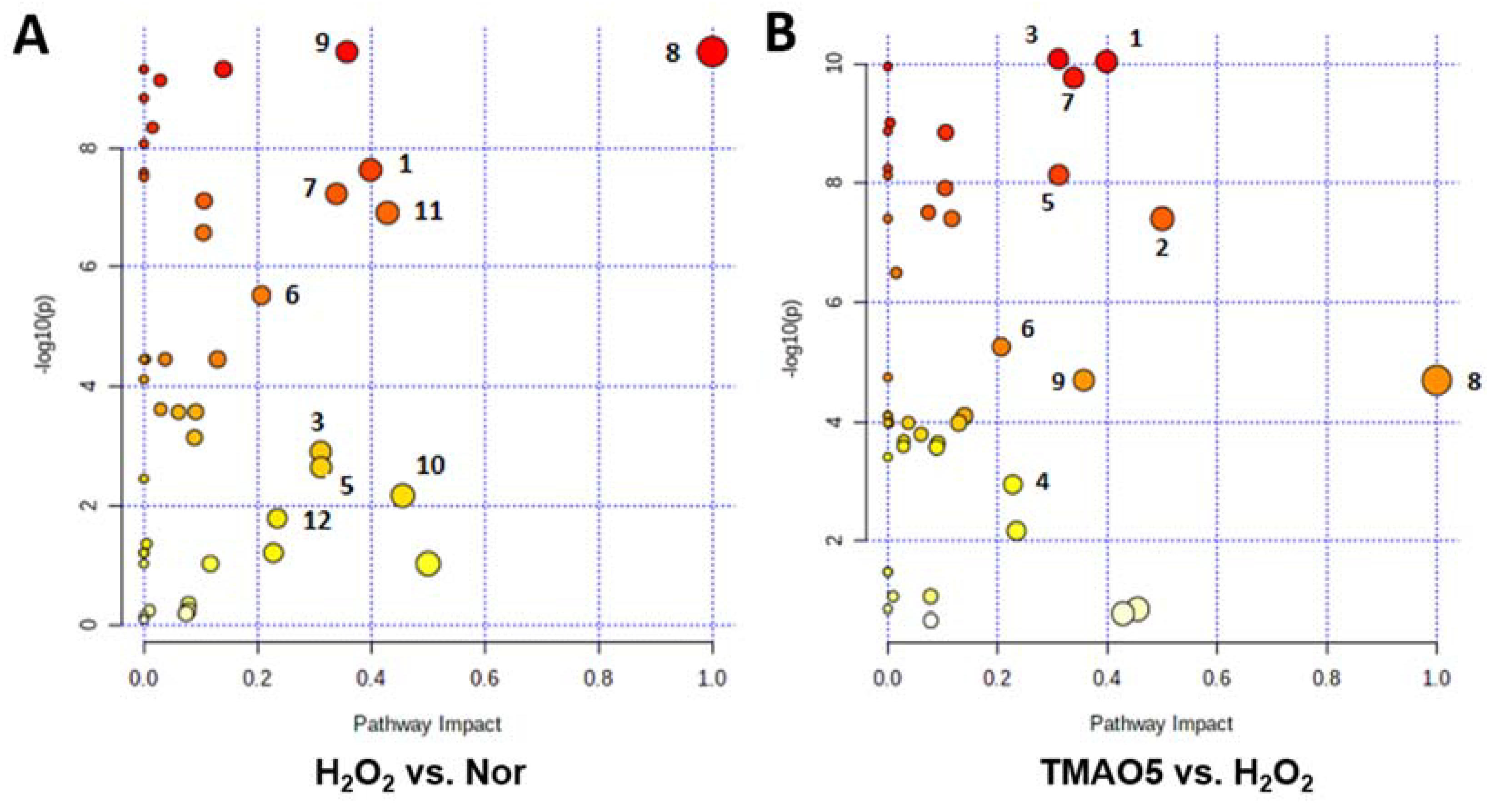
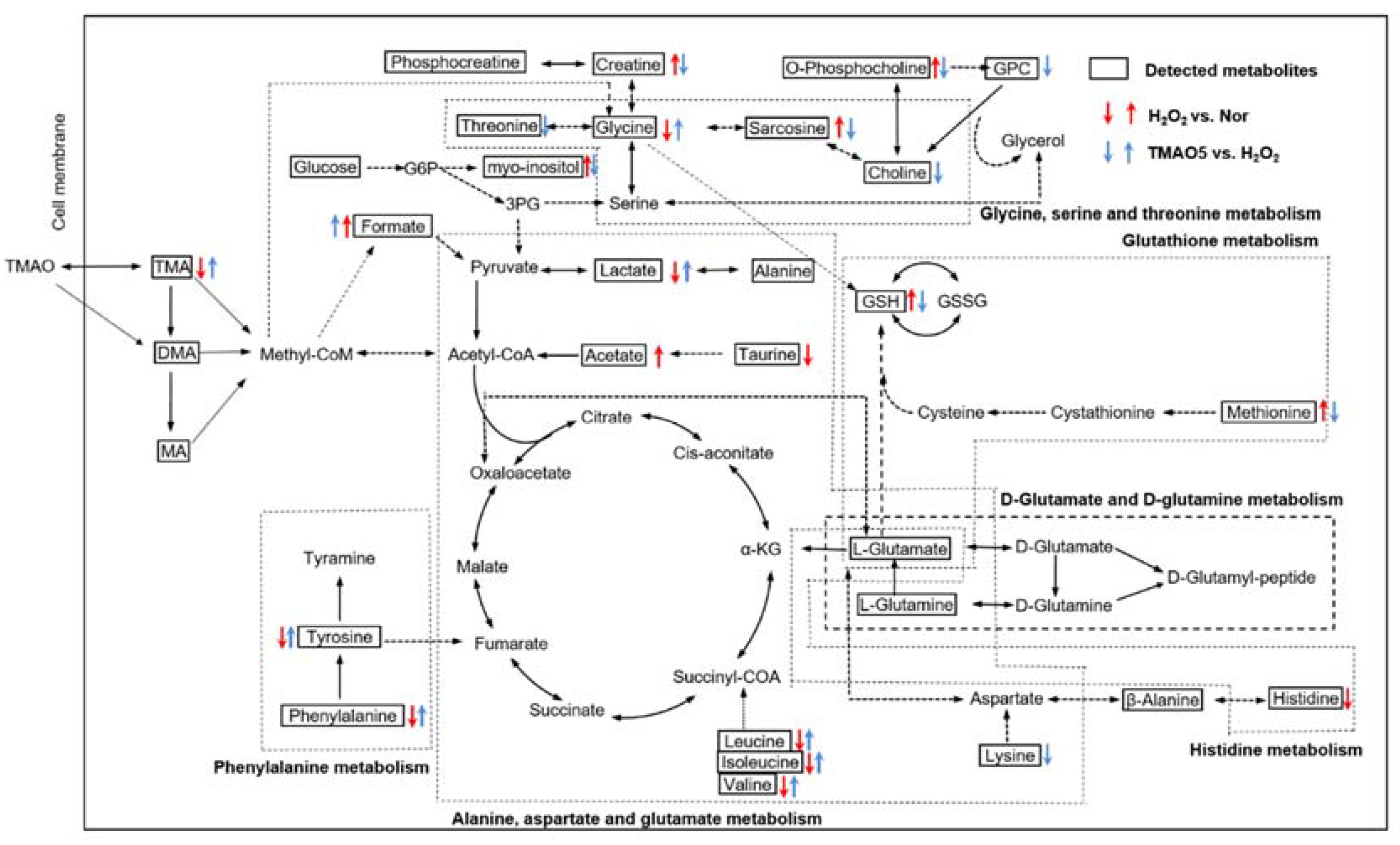
3.9. Identification of Significantly Altered Metabolic Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frontera, W.R.; Ochala, J. Skeletal muscle: A brief review of structure and function. Calcif. Tissue Int. 2015, 96, 183–195. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Lin, T.C.; Wang, S.H.; Huang, C.C.; Lai, Y.C.; Song, T.Y.; Tsai, M.S. Anti-Fatigue, Antioxidation, and Anti-Inflammatory Effects of Eucalyptus Oil Aromatherapy in Swimming-Exercised Rats. Chin. J. Physiol. 2018, 61, 257–265. [Google Scholar] [CrossRef]
- Barrea, L.; Annunziata, G.; Muscogiuri, G.; Laudisio, D.; Di Somma, C.; Maisto, M.; Tenore, G.C.; Colao, A.; Savastano, S. Trimethylamine N-oxide, Mediterranean diet, and nutrition in healthy, normal-weight adults: Also a matter of sex? Nutrition 2019, 62, 7–17. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J.; American Heart Association; Nutrition, Committee. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef]
- Asatoor, A.M.; Simenhoff, M.L. The origin of urinary dimethylamine. Biochim. Biophys. Acta. 1965, 111, 384–392. [Google Scholar] [CrossRef]
- Al-Waiz, M.; Mitchell, S.C.; Idle, J.R.; Smith, R.L. The relative importance of N-oxidation and N-demethylation in the metabolism of trimethylamine in man. Toxicology 1987, 43, 117–121. [Google Scholar] [CrossRef]
- Krueger, S.K.; Williams, D.E. Mammalian flavin-containing monooxygenases: Structure/function, genetic polymorphisms and role in drug metabolism. Pharmacol. Ther. 2005, 106, 357–387. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.H.; Hazen, S.L. The contributory role of gut microbiota in cardiovascular disease. J. Clin. Investig. 2014, 124, 4204–4211. [Google Scholar] [CrossRef]
- Zhuang, R.; Ge, X.; Han, L.; Yu, P.; Gong, X.; Meng, Q.; Zhang, Y.; Fan, H.; Zheng, L.; Liu, Z.; et al. Gut microbe-generated metabolite trimethylamine N-oxide and the risk of diabetes: A systematic review and dose-response meta-analysis. Obes. Rev. 2019, 20, 883–894. [Google Scholar] [CrossRef]
- Oellgaard, J.; Winther, S.A.; Hansen, T.S.; Rossing, P.; von Scholten, B.J. Trimethylamine N-oxide (TMAO) as a New Potential Therapeutic Target for Insulin Resistance and Cancer. Curr. Pharm. Des. 2017, 23, 3699–3712. [Google Scholar] [CrossRef]
- Barrea, L.; Annunziata, G.; Muscogiuri, G.; Di Somma, C.; Laudisio, D.; Maisto, M.; de Alteriis, G.; Tenore, G.C.; Colao, A.; Savastano, S. Trimethylamine-N-oxide (TMAO) as Novel Potential Biomarker of Early Predictors of Metabolic Syndrome. Nutrients 2018, 10, 1971. [Google Scholar] [CrossRef]
- Papandreou, C.; More, M.; Bellamine, A. Trimethylamine N-Oxide in Relation to Cardiometabolic Health-Cause or Effect? Nutrients 2020, 12, 1330. [Google Scholar] [CrossRef]
- Lupachyk, S.; Watcho, P.; Stavniichuk, R.; Shevalye, H.; Obrosova, I.G. Endoplasmic reticulum stress plays a key role in the pathogenesis of diabetic peripheral neuropathy. Diabetes 2013, 62, 944–952. [Google Scholar] [CrossRef]
- Woltjer, R.L.; McMahan, W.; Milatovic, D.; Kjerulf, J.D.; Shie, F.S.; Rung, L.G.; Montine, K.S.; Montine, T.J. Effects of chemical chaperones on oxidative stress and detergent-insoluble species formation following conditional expression of amyloid precursor protein carboxy-terminal fragment. Neurobiol. Dis. 2007, 25, 427–437. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. ‘Metabonomics’: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Everett, J.R.; Lindon, J.C. Longitudinal pharmacometabonomics for predicting patient responses to therapy: Drug metabolism, toxicity and efficacy. Expert Opin. Drug Metab. Toxicol. 2012, 8, 135–139. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Gao, Y.; Huang, C.; Lin, D. NMR-Based Metabolomic Analysis for the Effects of alpha-Ketoglutarate Supplementation on C2C12 Myoblasts in Different Energy States. Molecules 2021, 26, 1841. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Wilson, I.D. Opinion: Understanding ‘global’ systems biology: Metabonomics and the continuum of metabolism. Nat. Rev. Drug Discov. 2003, 2, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Han, M.H.; Lee, D.S.; Park, C.; Hong, S.H.; Kim, G.Y.; Hong, S.H.; Song, K.S.; Choi, I.W.; Cha, H.J.; et al. Morin exerts cytoprotective effects against oxidative stress in C2C12 myoblasts via the upregulation of Nrf2-dependent HO-1 expression and the activation of the ERK pathway. Int. J. Mol. Med. 2017, 39, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H. Berberine Hydrochloride Protects C2C12 Myoblast Cells Against Oxidative Stress-Induced Damage via Induction of Nrf-2-Mediated HO-1 Expression. Drug Dev. Res. 2016, 77, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Lu, R.; Huang, C.; Lin, D. Taurine Protects C2C12 Myoblasts From Impaired Cell Proliferation and Myotube Differentiation Under Cisplatin-Induced ROS Exposure. Front. Mol. Biosci. 2021, 8, 685362. [Google Scholar] [CrossRef]
- Alway, S.E.; Myers, M.J.; Mohamed, J.S. Regulation of satellite cell function in sarcopenia. Front. Aging Neurosci. 2014, 6, 246. [Google Scholar] [CrossRef]
- Kurosaka, M.; Ogura, Y.; Funabashi, T.; Akema, T. Early Growth Response 3 (Egr3) Contributes a Maintenance of C2C12 Myoblast Proliferation. J. Cell Physiol. 2017, 232, 1114–1122. [Google Scholar] [CrossRef]
- Pownall, M.E.; Gustafsson, M.K.; Emerson, C.P., Jr. Myogenic regulatory factors and the specification of muscle progenitors in vertebrate embryos. Annu. Rev. Cell Dev. Biol. 2002, 18, 747–783. [Google Scholar] [CrossRef]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building muscle: Molecular regulation of myogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008342. [Google Scholar] [CrossRef]
- Weintraub, H.; Davis, R.; Tapscott, S.; Thayer, M.; Krause, M.; Benezra, R.; Blackwell, T.K.; Turner, D.; Rupp, R.; Hollenberg, S.; et al. The myoD gene family: Nodal point during specification of the muscle cell lineage. Science 1991, 251, 761–766. [Google Scholar] [CrossRef]
- Zhang, A.Q.; Mitchell, S.C.; Smith, R.L. Dietary precursors of trimethylamine in man: A pilot study. Food Chem. Toxicol. 1999, 37, 515–520. [Google Scholar] [CrossRef]
- Cho, C.E.; Taesuwan, S.; Malysheva, O.V.; Bender, E.; Tulchinsky, N.F.; Yan, J.; Sutter, J.L.; Caudill, M.A. Trimethylamine-N-oxide (TMAO) response to animal source foods varies among healthy young men and is influenced by their gut microbiota composition: A randomized controlled trial. Mol. Nutr. Food Res. 2017, 61, 1600324. [Google Scholar] [CrossRef] [PubMed]
- Fennema, D.; Phillips, I.R.; Shephard, E.A. Trimethylamine and Trimethylamine N-Oxide, a Flavin-Containing Monooxygenase 3 (FMO3)-Mediated Host-Microbiome Metabolic Axis Implicated in Health and Disease. Drug Metab. Dispos. 2016, 44, 1839–1850. [Google Scholar] [CrossRef] [PubMed]
- Krueger, E.S.; Beales, J.L.; Russon, K.B.; Elison, W.S.; Davis, J.R.; Hansen, J.M.; Neilson, A.P.; Hansen, J.M.; Tessem, J.S. Gut Metabolite Trimethylamine N-Oxide Protects INS-1 beta-Cell and Rat Islet Function under Diabetic Glucolipotoxic Conditions. Biomolecules 2021, 11, 1892. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Huang, C.; Liu, Y.; Lin, D.; Zhao, Y. NMR-based metabolomic analysis of the effects of alanyl-glutamine supplementation on C2C12 myoblasts injured by energy deprivation. RSC Adv. 2018, 8, 16114–16125. [Google Scholar] [CrossRef] [PubMed]
- Eley, H.L.; Russell, S.T.; Tisdale, M.J. Effect of branched-chain amino acids on muscle atrophy in cancer cachexia. Biochem. J. 2007, 407, 113–120. [Google Scholar] [CrossRef]
- Berti Zanella, P.; Donner Alves, F.; Guerini de Souza, C. Effects of beta-alanine supplementation on performance and muscle fatigue in athletes and non-athletes of different sports: A systematic review. J. Sports Med. Phys. Fit. 2017, 57, 1132–1141. [Google Scholar] [CrossRef]
- Derave, W.; Everaert, I.; Beeckman, S.; Baguet, A. Muscle carnosine metabolism and beta-alanine supplementation in relation to exercise and training. Sports Med. 2010, 40, 247–263. [Google Scholar] [CrossRef]
- Vendelbo, M.H.; Moller, A.B.; Christensen, B.; Nellemann, B.; Clasen, B.F.; Nair, K.S.; Jorgensen, J.O.; Jessen, N.; Moller, N. Fasting increases human skeletal muscle net phenylalanine release and this is associated with decreased mTOR signaling. PLoS ONE 2014, 9, e102031. [Google Scholar] [CrossRef]
- Rousseau, A.S.; Hininger, I.; Palazzetti, S.; Faure, H.; Roussel, A.M.; Margaritis, I. Antioxidant vitamin status in high exposure to oxidative stress in competitive athletes. Br. J. Nutr. 2004, 92, 461–468. [Google Scholar] [CrossRef]
- Lee, H.; Ko, E.H.; Lai, M.; Wei, N.; Balroop, J.; Kashem, Z.; Zhang, M. Delineating the relationships among the formation of reactive oxygen species, cell membrane instability and innate autoimmunity in intestinal reperfusion injury. Mol. Immunol. 2014, 58, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Lin, D.; Huang, C. NMR-based metabolomic analysis for the effects of creatine supplementation on mouse myoblast cell line C2C12. Acta Biochim. Biophys. Sin. 2017, 49, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Mean ± Standard | Multiple Comparisons | One-Way ANOVA | |||
|---|---|---|---|---|---|---|
| Nor | H2O2 | TMAO5 | H2O2 vs. Nor | TMAO5 vs. H2O2 | p | |
| Pantothenate | 0.016 ± 0.002 | 0.009 ± 0.002 | 0.012 ± 0.002 | ↓↓↓↓ | ↑ | <0.0001 |
| Isoleucine | 0.769 ± 0.026 | 0.538 ± 0.013 | 0.573 ± 0.018 | ↓↓↓↓ | ↑↑ | <0.0001 |
| Leucine | 0.705 ± 0.027 | 0.476 ± 0.019 | 0.571 ± 0.027 | ↓↓↓↓ | ↑↑↑↑ | <0.0001 |
| Valine | 0.653 ± 0.027 | 0.435 ± 0.017 | 0.496 ± 0.024 | ↓↓↓↓ | ↑↑↑↑ | <0.0001 |
| Ethanol | 0.062 ± 0.017 | 0.078 ± 0.016 | 0.057 ± 0.011 | ns | ↓ | 0.7696 |
| Alanine | 1.172 ± 0.110 | 0.890 ± 0.047 | 1.617 ± 0.118 | ↓↓↓↓ | ↑↑↑↑ | <0.0001 |
| Putrescine | 0.092 ± 0.010 | 0.051 ± 0.012 | 0.086 ± 0.013 | ↓↓↓↓ | ↑↑↑ | <0.0001 |
| Acetate | 0.552 ± 0.393 | 0.849 ± 0.172 | 0.437 ± 0.177 | ↑↑↑ | ↓↓↓ | 0.0370 |
| Glutamate | 3.015 ± 0.076 | 3.150 ± 0.058 | 2.203 ± 0.151 | ↑↑ | ↓↓↓↓ | <0.0001 |
| Glutamine | 2.246 ± 0.193 | 2.148 ± 0.142 | 1.803 ± 0.148 | ns | ↓↓↓ | <0.0001 |
| Methylamine | 0.004 ± 0.002 | 0.002 ± 0.001 | 0.002 ± 0.002 | ns | ns | 0.1214 |
| Methionine | 0.313 ± 0.017 | 0.400 ± 0.013 | 0.289 ± 0.017 | ↑↑↑↑ | ↓↓↓↓ | <0.0001 |
| DMA | 0.004 ± 0.002 | 0.006 ± 0.004 | 0.002 ± 0.001 | ns | ns | 0.0747 |
| Sarcosine | 0.032 ± 0.002 | 0.042 ± 0.002 | 0.034 ± 0.003 | ↑↑↑↑ | ↓↓↓ | <0.0001 |
| TMA | 0.020 ± 0.010 | 0.005 ± 0.003 | 0.014 ± 0.003 | ↓↓↓ | ↑↑ | 0.0022 |
| Glutathione | 0.450 ± 0.064 | 0.599 ± 0.023 | 0.494 ± 0.073 | ↑↑↑ | ↓↓ | 0.0007 |
| Lysine | 0.270 ± 0.011 | 0.290 ± 0.024 | 0.260 ± 0.021 | ns | ↓ | 0.0334 |
| PCr | 1.449 ± 0.172 | 1.497 ± 0.099 | 1.610 ± 0.185 | ns | ns | 0.1536 |
| beta-Alanine | 0.439 ± 0.023 | 0.445 ± 0.032 | 0.472 ± 0.032 | ns | ns | 0.0776 |
| Choline | 5.027 ± 0.275 | 5.092 ± 0.162 | 3.321 ± 0.314 | ns | ↓↓↓↓ | <0.0001 |
| GPC | 0.637 ± 0.050 | 0.644 ± 0.049 | 0.482 ± 0.053 | ns | ↓↓↓↓ | <0.0001 |
| TMAO | 1.272 ± 0.062 | 1.494 ± 0.027 | 8.242 ± 0.291 | ns | ↑↑↑↑ | <0.0001 |
| Taurine | 2.255 ± 0.087 | 2.730 ± 0.068 | 2.615 ± 0.179 | ↑↑↑↑ | ns | <0.0001 |
| myo-Inositol | 2.478 ± 0.078 | 2.750 ± 0.081 | 2.322 ± 0.170 | ↑↑↑↑ | ↓↓↓ | <0.0001 |
| Glycine | 3.639 ± 0.205 | 2.487 ± 0.121 | 3.768 ± 0.299 | ↓↓↓↓ | ↑↑↑↑ | <0.0001 |
| Threonine | 0.708 ± 0.082 | 0.520 ± 0.020 | 0.569 ± 0.067 | ↓↓↓ | ns | <0.0001 |
| Creatine | 2.339 ± 0.280 | 3.404 ± 0.192 | 2.789 ± 0.428 | ↑↑↑↑ | ↓↓↓ | <0.0001 |
| Lactate | 1.254 ± 0.353 | 0.546 ± 0.069 | 1.676 ± 0.578 | ↓↓↓ | ↑↑↑ | 0.0003 |
| PC | 1.332 ± 0.082 | 1.517 ± 0.102 | 0.821 ± 0.106 | ↑↑ | ↓↓↓↓ | <0.0001 |
| Glucose | 0.077 ± 0.024 | 0.127 ± 0.016 | 0.191 ± 0.062 | ↑↑↑ | ↑ | 0.0001 |
| UDPG | 0.046 ± 0.005 | 0.047 ± 0.005 | 0.042 ± 0.006 | ns | ns | 0.1539 |
| UDP-GlcNAc | 0.250 ± 0.013 | 0.276 ± 0.007 | 0.222 ± 0.025 | ↑↑↑ | ↓↓↓ | <0.0001 |
| NADH | 0.677 ± 0.046 | 0.619 ± 0.025 | 0.605 ± 0.057 | ns | ns | 0.0157 |
| Carnosine | 0.039 ± 0.003 | 0.026 ± 0.003 | 0.028 ± 0.004 | ↓↓↓↓ | ns | <0.0001 |
| Tyrosine | 0.324 ± 0.014 | 0.199 ± 0.010 | 0.231 ± 0.010 | ↓↓↓↓ | ↑↑↑↑ | <0.0001 |
| Phenylalanine | 0.241 ± 0.009 | 0.148 ± 0.007 | 0.177 ± 0.007 | ↓↓↓↓ | ↑↑↑↑ | <0.0001 |
| Pyridoxine | 0.112 ± 0.075 | 0.086 ± 0.038 | 0.133 ± 0.079 | ns | ns | 0.4676 |
| Histidine | 0.084 ± 0.006 | 0.047 ± 0.021 | 0.055 ± 0.008 | ↓↓↓↓ | ns | <0.0001 |
| GTP | 0.045 ± 0.007 | 0.029 ± 0.003 | 0.052 ± 0.010 | ↓↓↓ | ↑↑↑ | 0.0001 |
| AMP | 0.954 ± 0.070 | 0.865 ± 0.036 | 0.815 ± 0.087 | ↓ | ns | 0.0029 |
| NAD+ | 0.076 ± 0.017 | 0.096 ± 0.002 | 0.079 ± 0.012 | ns | ns | 0.0251 |
| Formate | 0.046 ± 0.012 | 0.061 ± 0.007 | 0.075 ± 0.012 | ↑ | ↑ | 0.0002 |
| NADPH | 0.040 ± 0.015 | 0.024 ± 0.004 | 0.032 ± 0.010 | ↓↓↓ | ns | 0.0518 |
| ADP | 1.176 ± 0.079 | 1.023 ± 0.043 | 1.027 ± 0.123 | ↓↓ | ns | 0.0054 |
| IMP | 0.013 ± 0.011 | 0.001 ± 0.001 | 0.033 ± 0.024 | ↓ | ↑↑ | 0.0042 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, H.; Huang, C.; Zhou, L.; Lu, R.; Zhang, Y.; Lin, D. NMR-Based Metabolomic Analysis for the Effects of Trimethylamine N-Oxide Treatment on C2C12 Myoblasts under Oxidative Stress. Biomolecules 2022, 12, 1288. https://doi.org/10.3390/biom12091288
Zou H, Huang C, Zhou L, Lu R, Zhang Y, Lin D. NMR-Based Metabolomic Analysis for the Effects of Trimethylamine N-Oxide Treatment on C2C12 Myoblasts under Oxidative Stress. Biomolecules. 2022; 12(9):1288. https://doi.org/10.3390/biom12091288
Chicago/Turabian StyleZou, Hong, Caihua Huang, Lin Zhou, Ruohan Lu, Yimin Zhang, and Donghai Lin. 2022. "NMR-Based Metabolomic Analysis for the Effects of Trimethylamine N-Oxide Treatment on C2C12 Myoblasts under Oxidative Stress" Biomolecules 12, no. 9: 1288. https://doi.org/10.3390/biom12091288
APA StyleZou, H., Huang, C., Zhou, L., Lu, R., Zhang, Y., & Lin, D. (2022). NMR-Based Metabolomic Analysis for the Effects of Trimethylamine N-Oxide Treatment on C2C12 Myoblasts under Oxidative Stress. Biomolecules, 12(9), 1288. https://doi.org/10.3390/biom12091288