
Antizyme Inhibitor 2-Deficient Mice Exhibit Altered Brain Polyamine Levels and Reduced Locomotor Activity
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Quantitative Real-Time RT-qPCR
2.3. Ornithine Decarboxylase Activity Analysis
2.4. Generation and Genotyping of Transgenic Mice
2.5. Analysis of β-D-Galactosidase Activity
2.6. β-D-Galactosidase Reporter Staining and Immunocytochemistry
2.7. Polyamine Analysis
2.8. Evaluation of Motor Skills
2.9. Statistical Analysis
3. Results
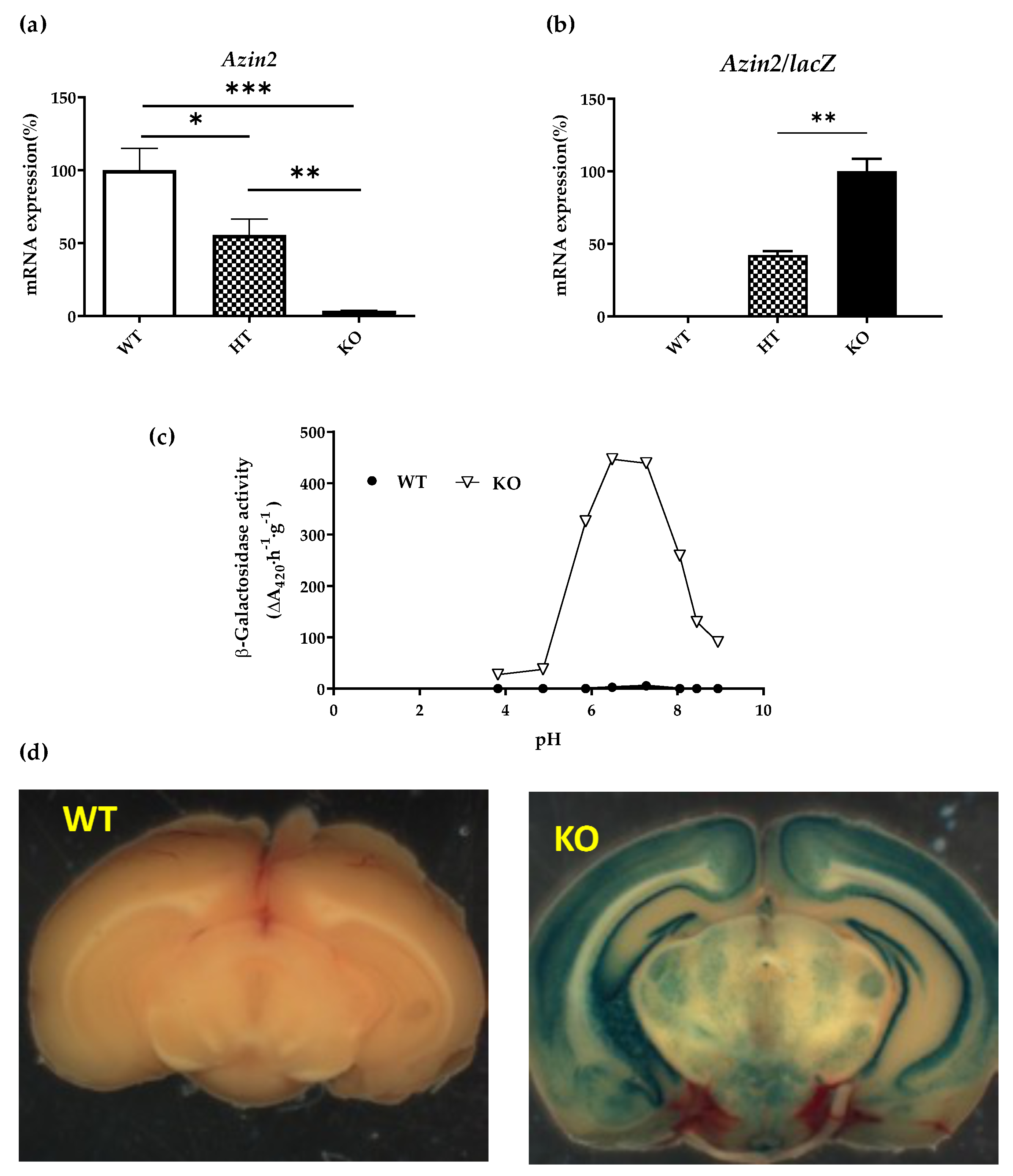
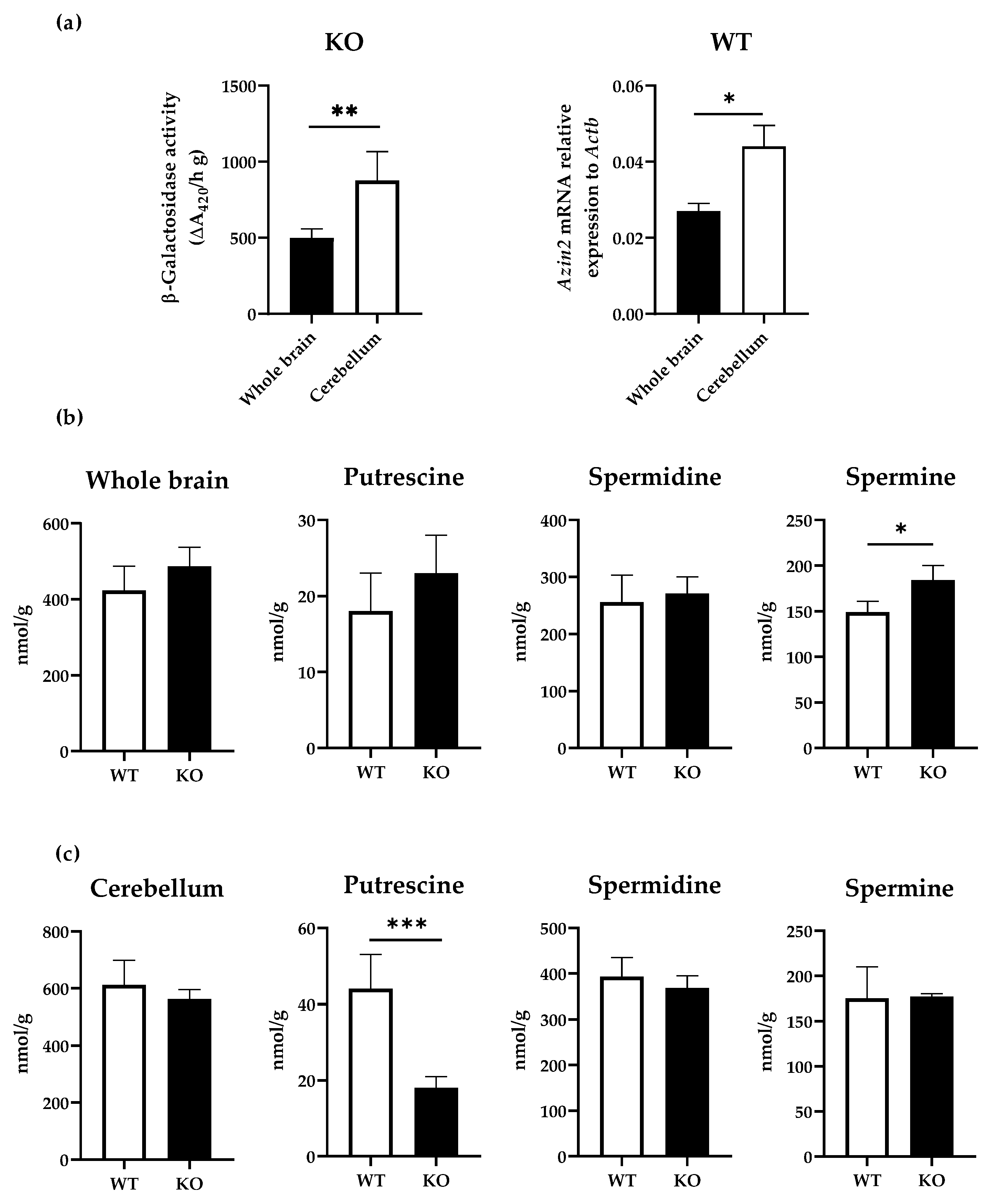
3.1. mRNA Expression Levels of Azin2 and Related Genes in the Mouse Brain
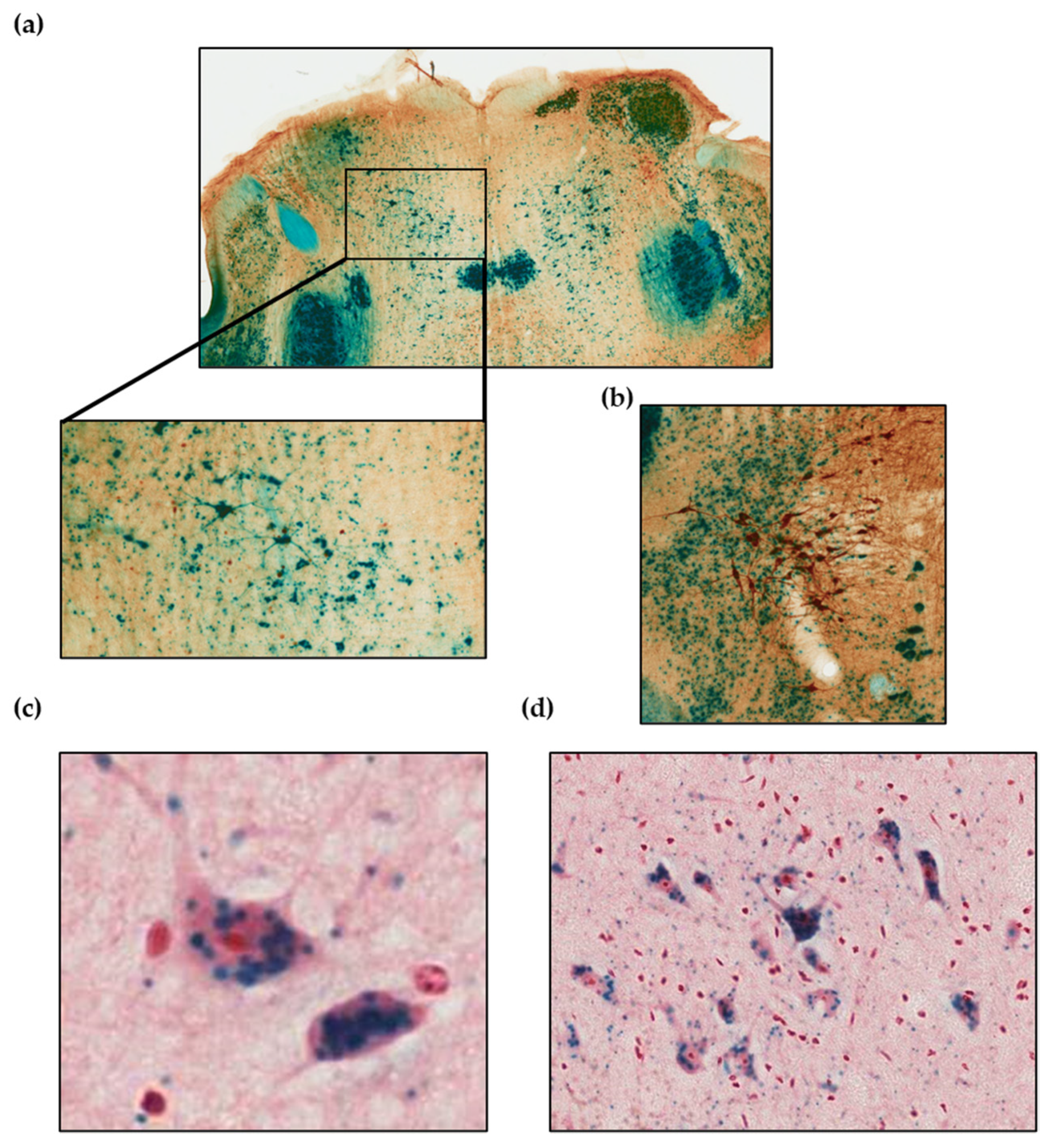
3.2. Azin2-lacZ Expression Pattern in the Mouse Brain
3.3. Influence of AZIN2 Ablation on Polyamine Metabolism in Brain Tissues
3.4. Functional Analysis of AZIN2 in Brain: Influence on Motor Activity
4. Discussions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef] [Green Version]
- Masuko, T.; Kusama-Eguchi, K.; Sakata, K.; Kusama, T.; Chaki, S.; Okuyama, S.; Williams, K.; Kashiwagi, K.; Igarashi, K. Polyamine transport, accumulation, and release in brain. J. Neurochem. 2003, 84, 610–617. [Google Scholar] [CrossRef]
- Bowie, D. Polyamine-mediated channel block of ionotropic glutamate receptors and its regulation by auxiliary proteins. J. Biol. Chem. 2018, 293, 18789–18802. [Google Scholar] [CrossRef] [Green Version]
- Dhara, M.; Matta, J.A.; Lei, M.; Knowland, D.; Yu, H.; Gu, S.; Bredt, D.S. Polyamine regulation of ion channel assembly and implications for nicotinic acetylcholine receptor pharmacology. Nat. Commun. 2020, 11, 2799. [Google Scholar] [CrossRef]
- Baronas, V.A.; Kurata, H.T. Inward rectifiers and their regulation by endogenous polyamines. Front. Physiol. 2014, 5, 325. [Google Scholar] [CrossRef] [Green Version]
- Duan, B.; Wang, Y.Z.; Yang, T.; Chu, X.P.; Yu, Y.; Huang, Y.; Cao, H.; Hansen, J.; Simon, R.P.; Zhu, M.X.; et al. Extracellular spermine exacerbates ischemic neuronal injury through sensitization of ASIC1a channels to extracellular acidosis. J. Neurosci. 2011, 31, 2101–2112. [Google Scholar] [CrossRef] [Green Version]
- Morrison, L.D.; Kish, S.J. Brain polyamine levels are altered in Alzheimer’s disease. Neurosci. Lett. 1995, 197, 5–8. [Google Scholar] [CrossRef]
- Ernestus, R.I.; Rohn, G.; Schroder, R.; Els, T.; Klekner, A.; Paschen, W.; Klug, N. Polyamine metabolism in brain tumours: Diagnostic relevance of quantitative biochemistry. J. Neurol. Neurosurg. Psychiatry 2001, 71, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, S.; Batliwalla, F.; Mocco, J.; Kiss, S.; Huang, J.; Mack, W.; Coon, A.; Eaton, J.W.; Al-Abed, Y.; Gregersen, P.K.; et al. Neuroprotection in cerebral ischemia by neutralization of 3-aminopropanal. Proc. Natl. Acad. Sci. USA 2002, 99, 5579–5584. [Google Scholar] [CrossRef] [Green Version]
- Baroli, G.; Sanchez, J.R.; Agostinelli, E.; Mariottini, P.; Cervelli, M. Polyamines: The possible missing link between mental disorders and epilepsy (Review). Int. J. Mol. Med. 2020, 45, 3–9. [Google Scholar] [CrossRef]
- Cervelli, M.; Averna, M.; Vergani, L.; Pedrazzi, M.; Amato, S.; Fiorucci, C.; Rossi, M.N.; Maura, G.; Mariottini, P.; Cervetto, C.; et al. The Involvement of Polyamines Catabolism in the Crosstalk between Neurons and Astrocytes in Neurodegeneration. Biomedicines 2022, 10, 1756. [Google Scholar] [CrossRef]
- Pegg, A.E. Regulation of ornithine decarboxylase. J. Biol. Chem. 2006, 281, 14529–14532. [Google Scholar] [CrossRef] [Green Version]
- Kahana, C. The antizyme family for regulating polyamines. J. Biol. Chem. 2018, 293, 18730–18735. [Google Scholar] [CrossRef] [Green Version]
- Matsufuji, S.; Matsufuji, T.; Miyazaki, Y.; Murakami, Y.; Atkins, J.F.; Gesteland, R.F.; Hayashi, S. Autoregulatory frameshifting in decoding mammalian ornithine decarboxylase antizyme. Cell 1995, 80, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Molina, B.; Lambertos, A.; Penafiel, R. Antizyme Inhibitors in Polyamine Metabolism and Beyond: Physiopathological Implications. Med. Sci. 2018, 6, 89. [Google Scholar] [CrossRef] [Green Version]
- Onoue, H.; Matsufuji, S.; Nishiyama, M.; Murakami, Y.; Hayashi, S. Changes in ornithine decarboxylase and antizyme activities in developing mouse brain. Biochem. J. 1988, 250, 797–803. [Google Scholar] [CrossRef] [Green Version]
- Pujic, Z.; Matsumoto, I.; Wilce, P.A. Expression of the genes coding for ornithine decarboxylase and its regulatory protein antizyme in the developing rat brain. Dev. Neurosci. 1995, 17, 286–291. [Google Scholar] [CrossRef]
- Kilpelainen, P.; Rybnikova, E.; Hietala, O.; Pelto-Huikko, M. Expression of ODC and its regulatory protein antizyme in the adult rat brain. J. Neurosci. Res. 2000, 62, 675–685. [Google Scholar] [CrossRef]
- Ramos-Molina, B.; Lopez-Contreras, A.J.; Cremades, A.; Penafiel, R. Differential expression of ornithine decarboxylase antizyme inhibitors and antizymes in rodent tissues and human cell lines. Amino Acids 2012, 42, 539–547. [Google Scholar] [CrossRef]
- Junttila, T.; Rechardt, L.; Hietala, O.A.; Pelto-Huikko, M. The expression of ornithine decarboxylase antizyme mRNA and protein in rat motoneurons. Neurosci. Lett. 1995, 197, 187–190. [Google Scholar] [CrossRef]
- Fernandes, J.R.D.; Jain, S.; Banerjee, A. Expression of ODC1, SPD, SPM and AZIN1 in the hypothalamus, ovary and uterus during rat estrous cycle. Gen. Comp. Endocrinol. 2017, 246, 9–22. [Google Scholar] [CrossRef]
- Greenwood, M.P.; Greenwood, M.; Paton, J.F.; Murphy, D. Control of Polyamine Biosynthesis by Antizyme Inhibitor 1 Is Important for Transcriptional Regulation of Arginine Vasopressin in the Male Rat Hypothalamus. Endocrinology 2015, 156, 2905–2917. [Google Scholar] [CrossRef] [Green Version]
- Makitie, L.T.; Kanerva, K.; Polvikoski, T.; Paetau, A.; Andersson, L.C. Brain neurons express ornithine decarboxylase-activating antizyme inhibitor 2 with accumulation in Alzheimer’s disease. Brain Pathol. 2010, 20, 571–580. [Google Scholar] [CrossRef]
- Sandusky-Beltran, L.A.; Kovalenko, A.; Placides, D.S.; Ratnasamy, K.; Ma, C.; Hunt, J.B., Jr.; Liang, H.; Calahatian, J.I.T.; Michalski, C.; Fahnestock, M.; et al. Aberrant AZIN2 and polyamine metabolism precipitates tau neuropathology. J. Clin. Investig. 2021, 131, e126299. [Google Scholar] [CrossRef]
- Zhu, M.Y.; Iyo, A.; Piletz, J.E.; Regunathan, S. Expression of human arginine decarboxylase, the biosynthetic enzyme for agmatine. Biochim. Biophys. Acta 2004, 1670, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Coleman, C.S.; Hu, G.; Pegg, A.E. Putrescine biosynthesis in mammalian tissues. Biochem. J. 2004, 379, 849–855. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Contreras, A.J.; Lopez-Garcia, C.; Jimenez-Cervantes, C.; Cremades, A.; Penafiel, R. Mouse ornithine decarboxylase-like gene encodes an antizyme inhibitor devoid of ornithine and arginine decarboxylating activity. J. Biol. Chem. 2006, 281, 30896–30906. [Google Scholar] [CrossRef] [Green Version]
- Kanerva, K.; Makitie, L.T.; Pelander, A.; Heiskala, M.; Andersson, L.C. Human ornithine decarboxylase paralogue (ODCp) is an antizyme inhibitor but not an arginine decarboxylase. Biochem. J. 2008, 409, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Garcia, C.; Ramos-Molina, B.; Lambertos, A.; Lopez-Contreras, A.J.; Cremades, A.; Penafiel, R. Antizyme inhibitor 2 hypomorphic mice. New patterns of expression in pancreas and adrenal glands suggest a role in secretory processes. PLoS ONE 2013, 8, e69188. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; McCarter, J.D.; Okamura-Oho, Y.; Yaghi, F.; Hinek, A.; Withers, S.G.; Callahan, J.W. Kinetic mechanism and characterization of human beta-galactosidase precursor secreted by permanently transfected Chinese hamster ovary cells. Biochem. J. 1994, 304 Pt 1, 281–288. [Google Scholar] [CrossRef]
- Stiles, J.; Jernigan, T.L. The basics of brain development. Neuropsychol. Rev. 2010, 20, 327–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanes, J.R.; Zipursky, S.L. Synaptic Specificity, Recognition Molecules, and Assembly of Neural Circuits. Cell 2020, 181, 536–556. [Google Scholar] [CrossRef]
- Suorsa, A.; Hietala, O.; Pajunen, A. Developmental expression of ornithine and S-adenosylmethionine decarboxylases in mouse brain. Biochem. Biophys. Res. Commun. 1992, 184, 1114–1118. [Google Scholar] [CrossRef]
- Fiori, L.M.; Turecki, G. Implication of the polyamine system in mental disorders. J. Psychiatry Neurosci. 2008, 33, 102–110. [Google Scholar]
- Wan, T.; Hu, Y.; Zhang, W.; Huang, A.; Yamamura, K.; Tang, H. Changes in liver gene expression of Azin1 knock-out mice. Z. Für Nat. C J. Biosci. 2010, 65, 519–527. [Google Scholar] [CrossRef]
- Tang, H.; Ariki, K.; Ohkido, M.; Murakami, Y.; Matsufuji, S.; Li, Z.; Yamamura, K. Role of ornithine decarboxylase antizyme inhibitor in vivo. Genes Cells 2009, 14, 79–87. [Google Scholar] [CrossRef]
- Martinez-de-la-Torre, M.; Lambertos, A.; Penafiel, R.; Puelles, L. An exercise in brain genoarchitectonics: Analysis of AZIN2-Lacz expressing neuronal populations in the mouse hindbrain. J. Neurosci. Res. 2018, 96, 1490–1517. [Google Scholar] [CrossRef]
- Rasila, T.; Lehtonen, A.; Kanerva, K.; Makitie, L.T.; Haglund, C.; Andersson, L.C. Expression of ODC Antizyme Inhibitor 2 (AZIN2) in Human Secretory Cells and Tissues. PLoS ONE 2016, 11, e0151175. [Google Scholar] [CrossRef] [Green Version]
- Kanerva, K.; Makitie, L.T.; Back, N.; Andersson, L.C. Ornithine decarboxylase antizyme inhibitor 2 regulates intracellular vesicle trafficking. Exp. Cell Res. 2010, 316, 1896–1906. [Google Scholar] [CrossRef]
- Silva, T.M.; Cirenajwis, H.; Wallace, H.M.; Oredsson, S.; Persson, L. A role for antizyme inhibitor in cell proliferation. Amino Acids 2015, 47, 1341–1352. [Google Scholar] [CrossRef] [Green Version]
- Lewandowski, N.M.; Ju, S.; Verbitsky, M.; Ross, B.; Geddie, M.L.; Rockenstein, E.; Adame, A.; Muhammad, A.; Vonsattel, J.P.; Ringe, D.; et al. Polyamine pathway contributes to the pathogenesis of Parkinson disease. Proc. Natl. Acad. Sci. USA 2010, 107, 16970–16975. [Google Scholar] [CrossRef]
- Polis, B.; Karasik, D.; Samson, A.O. Alzheimer’s disease as a chronic maladaptive polyamine stress response. Aging 2021, 13, 10770–10795. [Google Scholar] [CrossRef] [PubMed]
- Prokop, J.W.; Bupp, C.P.; Frisch, A.; Bilinovich, S.M.; Campbell, D.B.; Vogt, D.; Schultz, C.R.; Uhl, K.L.; VanSickle, E.; Rajasekaran, S.; et al. Emerging Role of ODC1 in Neurodevelopmental Disorders and Brain Development. Genes 2021, 12, 470. [Google Scholar] [CrossRef] [PubMed]
- Halmekyto, M.; Alhonen, L.; Wahlfors, J.; Sinervirta, R.; Eloranta, T.; Janne, J. Characterization of a transgenic mouse line over-expressing the human ornithine decarboxylase gene. Biochem. J. 1991, 278 Pt 3, 895–898. [Google Scholar] [CrossRef] [Green Version]
- Rieck, J.; Skatchkov, S.N.; Derst, C.; Eaton, M.J.; Veh, R.W. Unique Chemistry, Intake, and Metabolism of Polyamines in the Central Nervous System (CNS) and Its Body. Biomolecules 2022, 12, 501. [Google Scholar] [CrossRef] [PubMed]
- Koziol, L.F.; Budding, D.; Andreasen, N.; D’Arrigo, S.; Bulgheroni, S.; Imamizu, H.; Ito, M.; Manto, M.; Marvel, C.; Parker, K.; et al. Consensus paper: The cerebellum’s role in movement and cognition. Cerebellum 2014, 13, 151–177. [Google Scholar] [CrossRef]
- Kono, M.; Kakegawa, W.; Yoshida, K.; Yuzaki, M. Interneuronal NMDA receptors regulate long-term depression and motor learning in the cerebellum. J. Physiol. 2019, 597, 903–920. [Google Scholar] [CrossRef] [Green Version]
- Rioult-Pedotti, M.S.; Pekanovic, A.; Atiemo, C.O.; Marshall, J.; Luft, A.R. Dopamine Promotes Motor Cortex Plasticity and Motor Skill Learning via PLC Activation. PLoS ONE 2015, 10, e0124986. [Google Scholar] [CrossRef] [Green Version]
- Lisman, J.; Buzsaki, G.; Eichenbaum, H.; Nadel, L.; Ranganath, C.; Redish, A.D. Viewpoints: How the hippocampus contributes to memory, navigation and cognition. Nat. Neurosci. 2017, 20, 1434–1447. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ROTA-ROD | |||
| WT (n = 17) | KO (n = 17) | p-value | |
| Score (cm/min) | 107 ± 13 | 88 ± 24 | 0.03 |
| Time (s) | 313 ± 27 | 42 ± 36 | < 0.0001 |
| POLE TEST | |||
| WT (n = 16) | KO (n = 16) | ||
| Successful rate | 15/16 | 0/16 | |
| T-turn (s) | 1.55 ± 3.2 | ||
| T-total (s) | 5.6 ± 3.2 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lambertos, A.; Nuñez-Sanchez, M.A.; López-García, C.; López-Contreras, A.J.; Ramos-Molina, B.; Peñafiel, R. Antizyme Inhibitor 2-Deficient Mice Exhibit Altered Brain Polyamine Levels and Reduced Locomotor Activity. Biomolecules 2023, 13, 14. https://doi.org/10.3390/biom13010014
Lambertos A, Nuñez-Sanchez MA, López-García C, López-Contreras AJ, Ramos-Molina B, Peñafiel R. Antizyme Inhibitor 2-Deficient Mice Exhibit Altered Brain Polyamine Levels and Reduced Locomotor Activity. Biomolecules. 2023; 13(1):14. https://doi.org/10.3390/biom13010014
Chicago/Turabian StyleLambertos, Ana, Maria Angeles Nuñez-Sanchez, Carlos López-García, Andrés Joaquín López-Contreras, Bruno Ramos-Molina, and Rafael Peñafiel. 2023. "Antizyme Inhibitor 2-Deficient Mice Exhibit Altered Brain Polyamine Levels and Reduced Locomotor Activity" Biomolecules 13, no. 1: 14. https://doi.org/10.3390/biom13010014
APA StyleLambertos, A., Nuñez-Sanchez, M. A., López-García, C., López-Contreras, A. J., Ramos-Molina, B., & Peñafiel, R. (2023). Antizyme Inhibitor 2-Deficient Mice Exhibit Altered Brain Polyamine Levels and Reduced Locomotor Activity. Biomolecules, 13(1), 14. https://doi.org/10.3390/biom13010014