
GABA-A Alpha 2/3 but Not Alpha 1 Receptor Subunit Ligand Inhibits Harmaline and Pimozide-Induced Tremor in Rats

 , ,
, ,  , and
, and 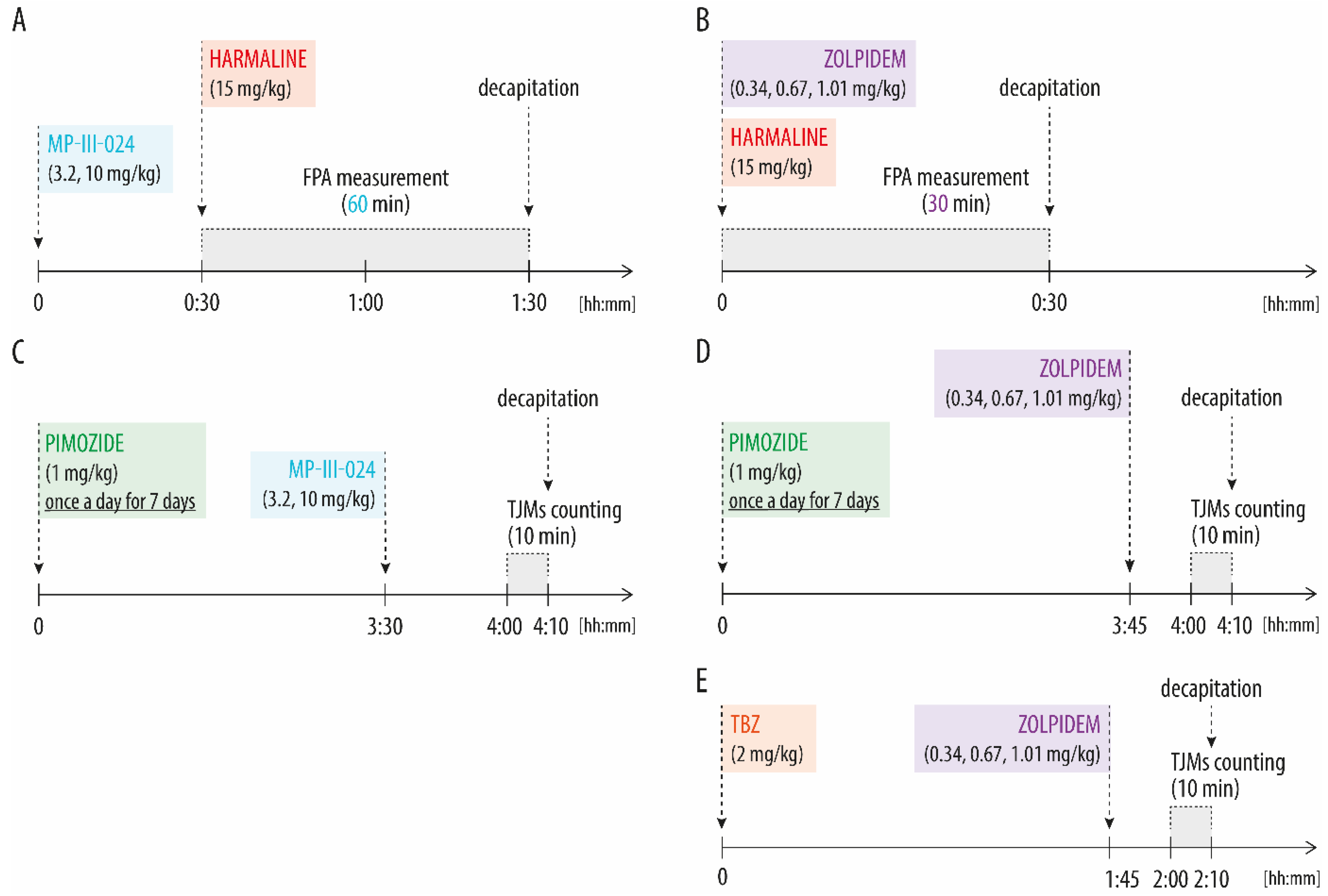{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Essential Tremor
1.2. PD Tremor
1.3. GABA-A α2/3 Subunit Specific Receptors as Novel Drug Targets against Tremor
2. Materials and Methods
2.1. Animals
2.2. Compounds
2.3. Experimental Procedures
2.3.1. Harmaline-Induced Tremor and Locomotor Activity Analysis
2.3.2. Tremulous Jaw Movements (TJMs) Analysis
2.4. Statistical Analysis
3. Results
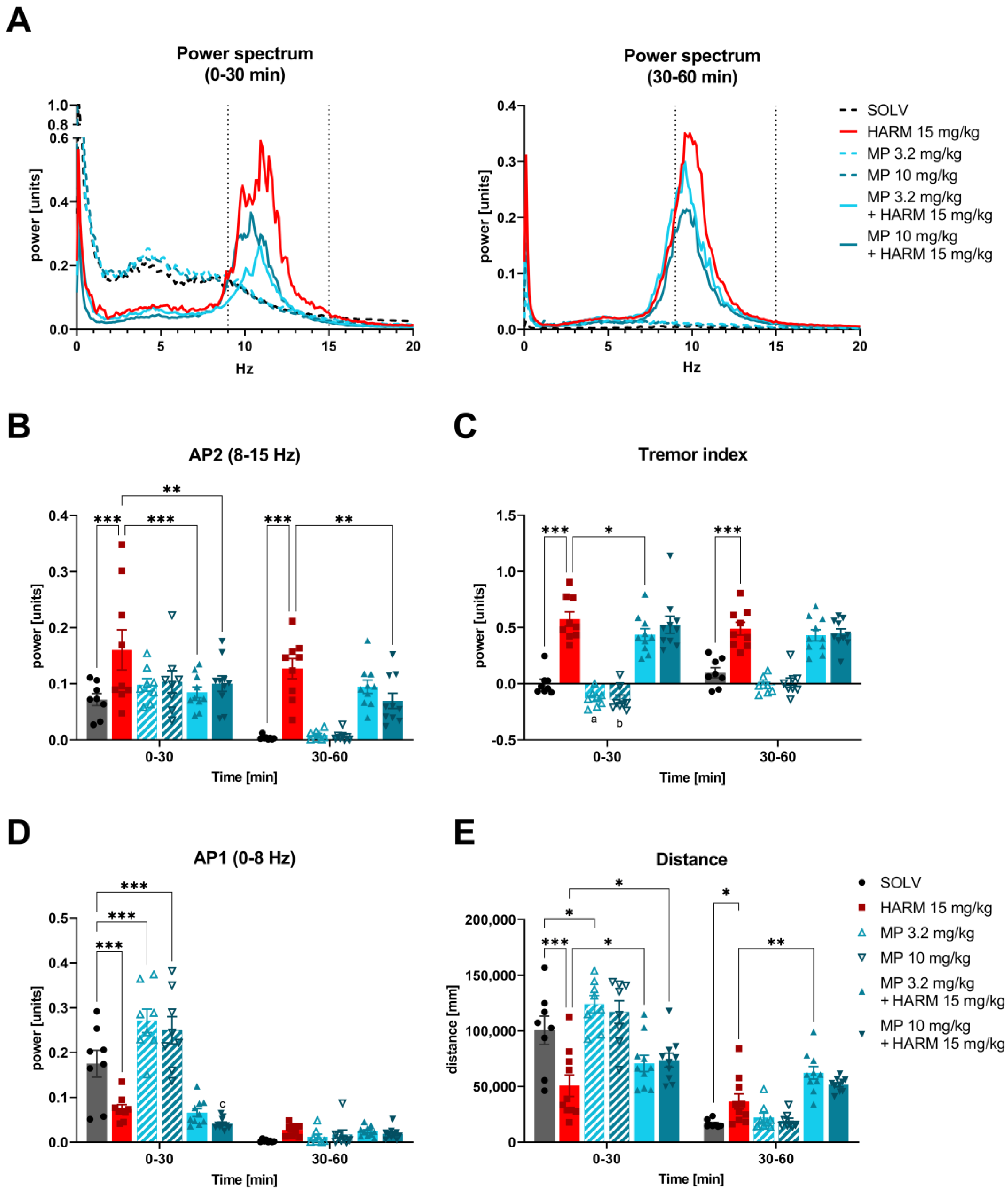
3.1. MP-III-024 Inhibits Harmaline-Induced Tremor and Increases the Locomotor Activity
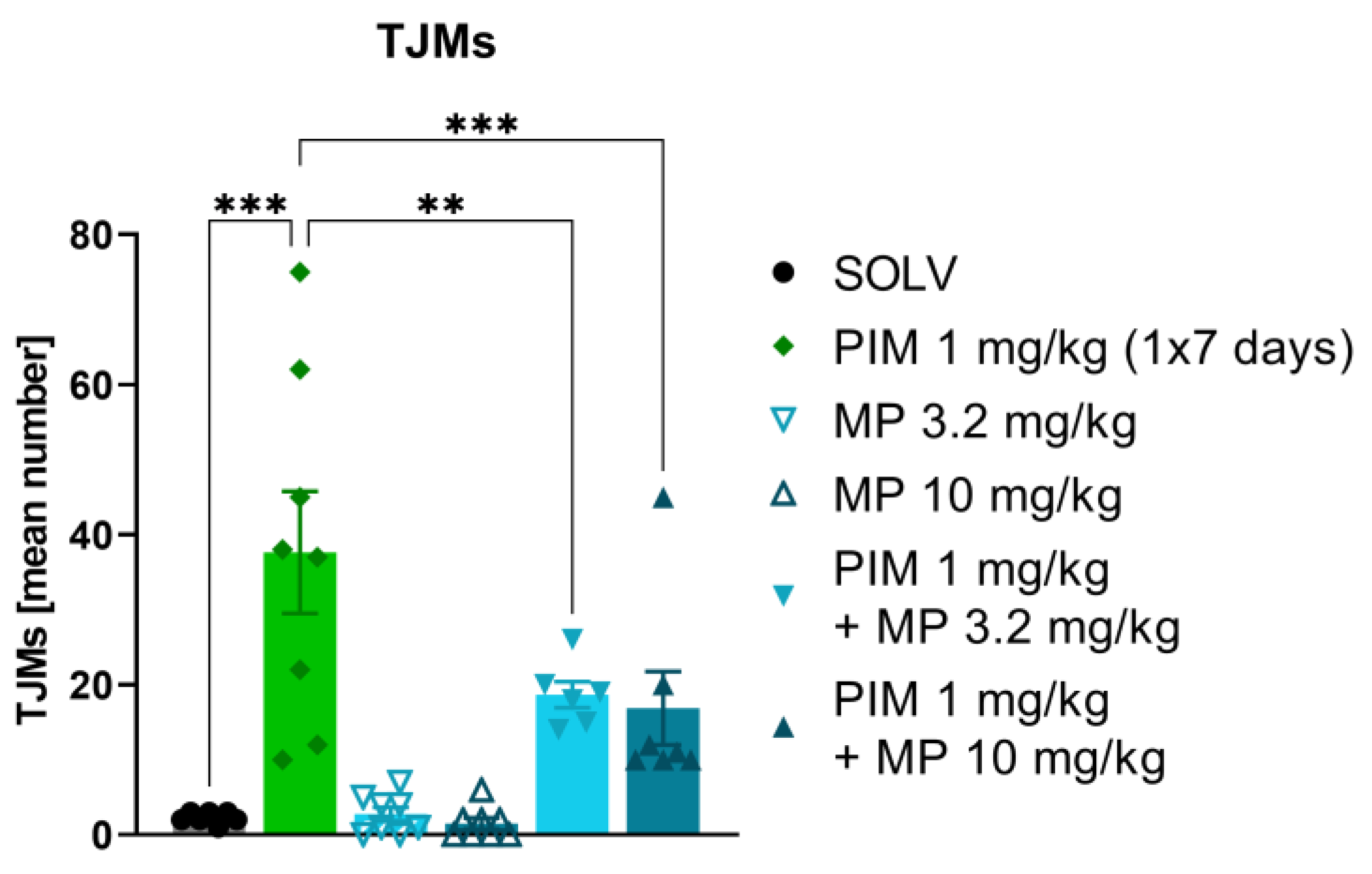
3.2. MP-III-024 Inhibits the Pimozide-Induced TJMs
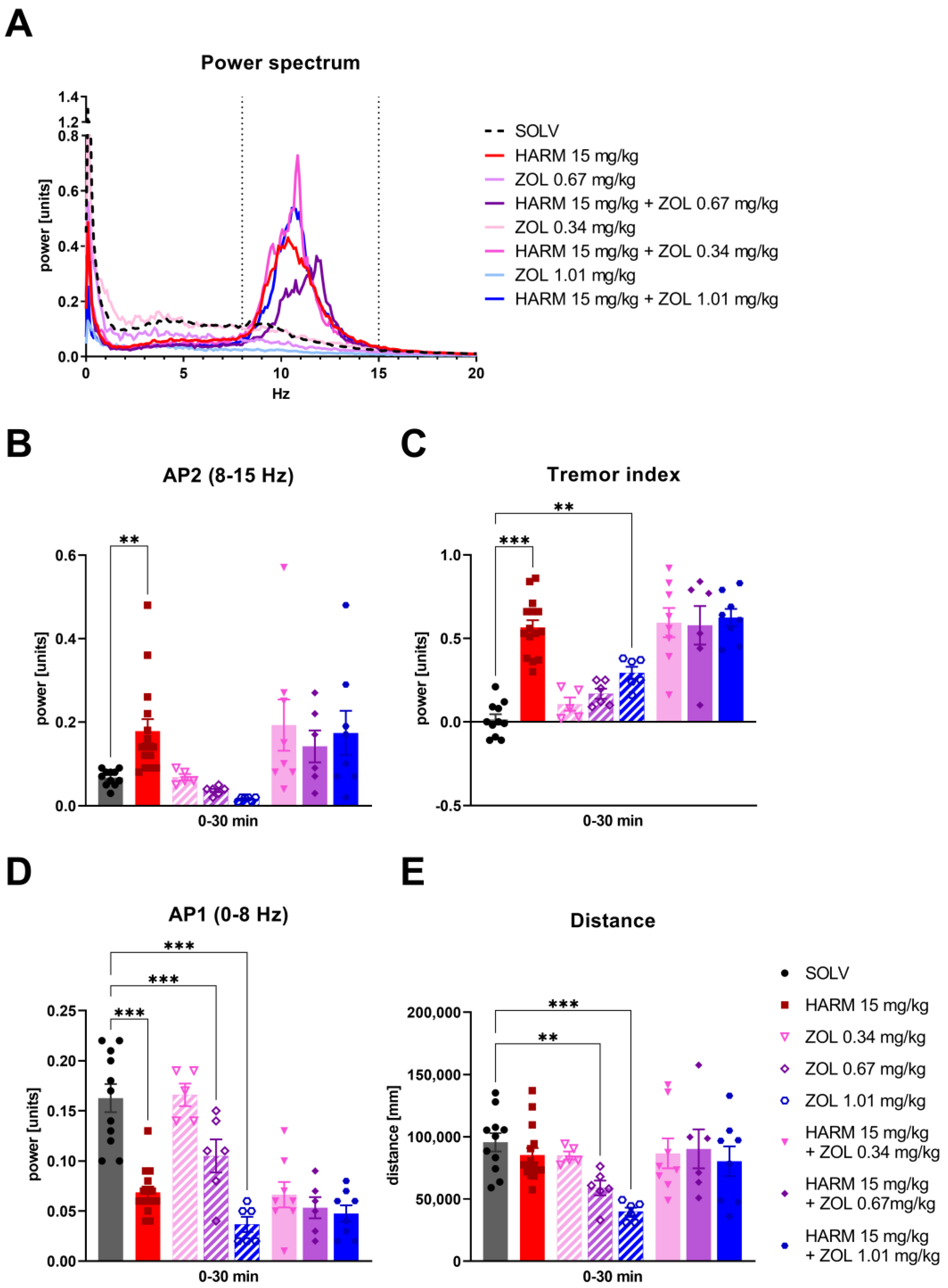
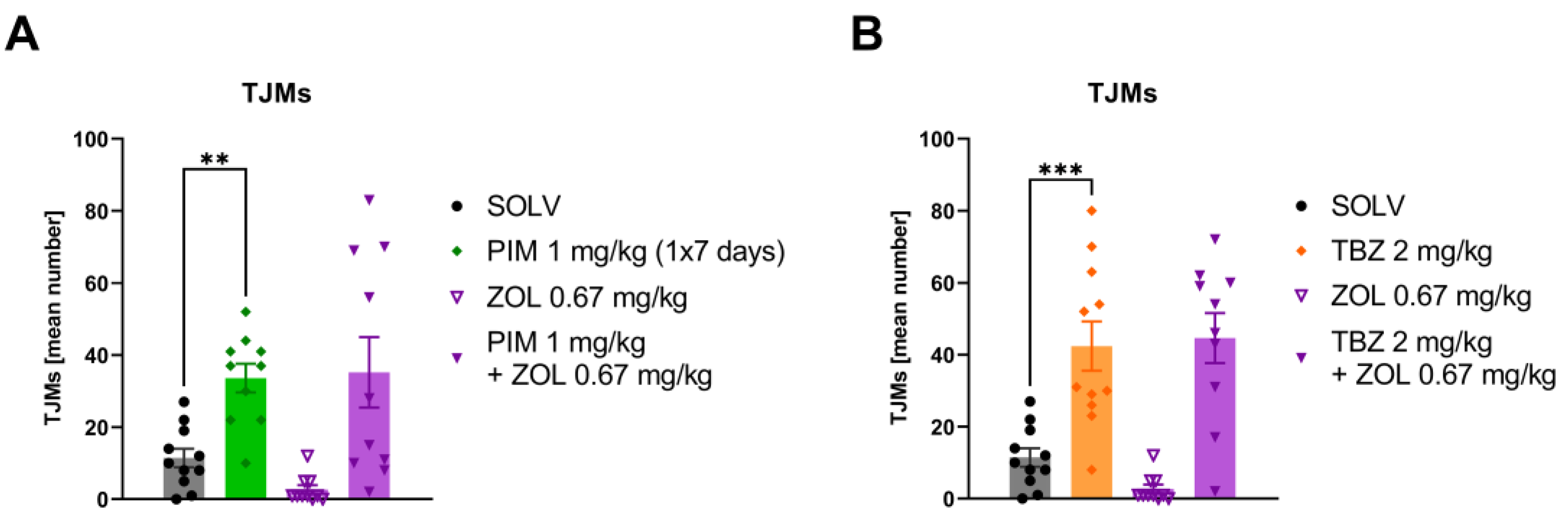
3.3. Zolpidem Has No Effect on Either Harmaline-Induced Tremor or TJMs after Pimozide or TBZ
4. Discussion
4.1. The Tremorolytic Effect of GABA-A α2/3 Subtype Selective Positive Allosteric Modulator MP-III-024
4.2. Different Tremor Models, Same Effect—Why?
4.3. Lack of Tremorolytic Effect of Zolpidem in Experimental Studies
4.4. Lack of Tremorolytic Effect of Zolpidem in ET Model
4.5. GABA-A α1 vs. α2/3 Subunit Localisation
4.6. Effect on Locomotor Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elble, R.J. What Is Essential Tremor? Curr. Neurol. Neurosci. Rep. 2013, 13, 353. [Google Scholar] [CrossRef] [PubMed]
- Kosmowska, B.; Wardas, J. The Pathophysiology and Treatment of Essential Tremor: The Role of Adenosine and Dopamine Receptors in Animal Models. Biomolecules 2021, 11, 1813. [Google Scholar] [CrossRef] [PubMed]
- Rajput, A.H.; Adler, C.H.; Shill, H.A.; Rajput, A. Essential Tremor Is Not a Neurodegenerative Disease. Neurodegener. Dis. Manag. 2012, 2, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axelrad, J.E.; Louis, E.D.; Honig, L.S.; Flores, I.; Ross, G.W.; Pahwa, R.; Lyons, K.E.; Faust, P.L.; Vonsattel, J.P.G. Reduced Purkinje Cell Number in Essential Tremor. Arch. Neurol. 2008, 65, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Choe, M.; Cortés, E.; Vonsattel, J.P.G.; Kuo, S.H.; Faust, P.L.; Louis, E.D. Purkinje Cell Loss in Essential Tremor: Random Sampling Quantification and Nearest Neighbor Analysis. Mov. Disord. 2016, 31, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Louis, E.D.; Faust, P.L.; Vonsattel, J.P.G.; Honig, L.S.; Rajput, A.; Robinson, C.A.; Rajput, A.; Pahwa, R.; Lyons, K.E.; Ross, G.W.; et al. Neuropathological Changes in Essential Tremor: 33 Cases Compared with 21 Controls. Brain 2007, 130, 3297–3307. [Google Scholar] [CrossRef] [Green Version]
- Symanski, C.; Shill, H.A.; Dugger, B.; Hentz, J.G.; Adler, C.H.; Jacobson, S.A.; Driver-Dunckley, E.; Beach, T.G. Essential Tremor Is Not Associated with Cerebellar Purkinje Cell Loss. Mov. Disord. 2014, 29, 496–500. [Google Scholar] [CrossRef]
- Sharifi, S.; Nederveen, A.J.; Booij, J.; van Rootselaar, A.F. Neuroimaging Essentials in Essential Tremor: A Systematic Review. Neuroimage Clin. 2014, 5, 217–231. [Google Scholar] [CrossRef] [Green Version]
- Uusisaari, M.Y.; Knöpfel, T. GABAergic Synaptic Communication in the GABAergic and Non-GABAergic Cells in the Deep Cerebellar Nuclei. Neuroscience 2008, 156, 537–549. [Google Scholar] [CrossRef]
- Uusisaari, M.Y.; Knöpfel, T. Diversity of Neuronal Elements and Circuitry in the Cerebellar Nuclei. Cerebellum 2012, 11, 420–421. [Google Scholar] [CrossRef]
- Helmich, R.C.; Toni, I.; Deuschl, G.; Bloem, B.R. The Pathophysiology of Essential Tremor and Parkinson’s Tremor. Curr. Neurol. Neurosci. Rep. 2013, 13, 378. [Google Scholar] [CrossRef] [PubMed]
- Hedera, P.; Cibulčík, F.; Davis, T.L. Pharmacotherapy of Essential Tremor. J. Cent. Nerv. Syst. Dis. 2013, 5, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Ondo, W.G. Current and Emerging Treatments of Essential Tremor. Neurol. Clin. 2020, 38, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Zesiewicz, T.A.; Shaw, J.D.; Allison, K.G.; Staffetti, J.S.; Okun, M.S.; Sullivan, K.L. Update on Treatment of Essential Tremor. Curr. Treat. Options Neurol. 2013, 15, 410–423. [Google Scholar] [CrossRef] [PubMed]
- Kralic, J.E.; Criswell, H.E.; Osterman, J.L.; O’Buckley, T.K.; Wilkie, M.E.; Matthews, D.B.; Hamre, K.; Breese, G.R.; Homanics, G.E.; Morrow, A.L. Genetic Essential Tremor in Gamma-Aminobutyric AcidA Receptor Alpha1 Subunit Knockout Mice. J. Clin. Investiig. 2005, 115, 774–779. [Google Scholar] [CrossRef] [Green Version]
- Nietz, A.; Krook-Magnuson, C.; Gutierrez, H.; Klein, J.; Sauve, C.; Hoff, I.; Wick, Z.C.; Krook-Magnuson, E. Selective Loss of the GABAAα1 Subunit from Purkinje Cells Is Sufficient to Induce a Tremor Phenotype. J. Neurophysiol. 2020, 124, 1183–1197. [Google Scholar] [CrossRef]
- Handforth, A. Harmaline Tremor: Underlying Mechanisms in a Potential Animal Model of Essential Tremor. Tremor Other Hyperkinetic Mov. 2012, 2, 02-92-769–1. [Google Scholar] [CrossRef]
- Martin, F.C.; Le, A.T.; Handforth, A. Harmaline-Induced Tremor as a Potential Preclinical Screening Method for Essential Tremor Medications. Mov. Disord. 2005, 20, 298–305. [Google Scholar] [CrossRef]
- Miwa, H. Rodent Models of Tremor. Cerebellum 2007, 6, 66–72. [Google Scholar] [CrossRef]
- Helmich, R.C.; Dirkx, M. Pathophysiology and Management of Parkinsonian Tremor. Semin. Neurol. 2017, 37, 127–134. [Google Scholar] [CrossRef]
- Deuschl, G.; Raethjen, J.; Baron, R.; Lindemann, M.; Wilms, H.; Krack, P. The Pathophysiology of Parkinsonian Tremor: A Review. J. Neurol. 2000, 247 (Suppl. S5), V33–V48. [Google Scholar] [CrossRef] [PubMed]
- DeLong, M.R.; Wichmann, T. Basal Ganglia Circuits as Targets for Neuromodulation in Parkinson Disease. JAMA Neurol. 2015, 72, 1354. [Google Scholar] [CrossRef] [PubMed]
- Gerfen, C.R. The Neostriatal Mosaic: Multiple Levels of Compartmental Organization. Trends Neurosci. 1992, 15, 133–139. [Google Scholar] [CrossRef]
- Castela, I.; Hernandez, L.F. Shedding Light on Dyskinesias. Eur. J. Neurosci. 2021, 53, 2398–2413. [Google Scholar] [CrossRef] [PubMed]
- Helmich, R.C. The Cerebral Basis of Parkinsonian Tremor: A Network Perspective. Mov. Disord. 2018, 33, 219–231. [Google Scholar] [CrossRef]
- Hallett, M. Parkinson’s Disease Tremor: Pathophysiology. Park. Relat. Disord. 2012, 18, S85–S86. [Google Scholar] [CrossRef]
- Salamone, J.D.; Mayorga, A.J.; Trevitt, J.T.; Cousins, M.S.; Conlan, A.; Nawab, A. Tremulous Jaw Movements in Rats: A Model of Parkinsonian Tremor. Prog. Neurobiol. 1998, 56, 591–611. [Google Scholar] [CrossRef]
- Ionov, I.D.; Pushinskaya, I.I.; Frenkel, D.D.; Gorev, N.P.; Shpilevaya, L.A. Neuroanatomical Correlates of the Inhibition of Tremulous Jaw Movements in Rats by a Combination of Memantine and Δ 9-tetrahydrocannabinol. Br. J. Pharmacol. 2020, 177, 1514–1524. [Google Scholar] [CrossRef]
- Rabey, J.M.; Nissipeanu, P.; Korczyn, A.D. Efficacy of Memantine, an NMDA Receptor Antagonist, in the Treatment of Parkinson’s Disease. J. Neural. Transm. Park. Dis. Dement. Sect. 1992, 4, 277–282. [Google Scholar] [CrossRef]
- Bostan, A.C.; Dum, R.P.; Strick, P.L. The Basal Ganglia Communicate with the Cerebellum. Proc. Natl. Acad. Sci. USA 2010, 107, 8452–8456. [Google Scholar] [CrossRef]
- Helmich, R.C.; Hallett, M.; Deuschl, G.; Toni, I.; Bloem, B.R. Cerebral Causes and Consequences of Parkinsonian Resting Tremor: A Tale of Two Circuits? Brain 2012, 135, 3206–3226. [Google Scholar] [CrossRef] [Green Version]
- Antonini, A.; Moeller, J.R.; Nakamura, T.; Spetsieris, P.; Dhawan, V.; Eidelberg, D. The Metabolic Anatomy of Tremor in Parkinson’s Disease. Neurology 1998, 51, 803–810. [Google Scholar] [CrossRef]
- Helmich, R.C.; Janssen, M.J.R.; Oyen, W.J.G.; Bloem, B.R.; Toni, I. Pallidal Dysfunction Drives a Cerebellothalamic Circuit into Parkinson Tremor. Ann. Neurol. 2011, 69, 269–281. [Google Scholar] [CrossRef]
- van den Berg, K.R.E.; Helmich, R.C. The Role of the Cerebellum in Tremor—Evidence from Neuroimaging. Tremor Other Hyperkinet. Mov. 2021, 11, 49. [Google Scholar] [CrossRef]
- Finn, M.; Mayorga, A.J.; Conlan, A.; Salamone, J.D. Involvement of Pallidal and Nigral Gaba Mechanisms in the Generation of Tremulous Jaw Movements in Rats. Neuroscience 1997, 80, 535–544. [Google Scholar] [CrossRef]
- Zhang, X.; Santaniello, S. Role of Cerebellar GABAergic Dysfunctions in the Origins of Essential Tremor. Proc. Natl. Acad. Sci. USA 2019, 116, 13592–13601. [Google Scholar] [CrossRef] [Green Version]
- Bullock, A.; Kaul, I.; Li, S.; Silber, C.; Doherty, J.; Kanes, S.J. Zuranolone as an Oral Adjunct to Treatment of Parkinsonian Tremor: A Phase 2, Open-Label Study. J. Neurol. Sci. 2021, 421, 117277. [Google Scholar] [CrossRef]
- Daniele, A.; Panza, F.; Greco, A.; Logroscino, G.; Seripa, D. Can a Positive Allosteric Modulation of GABAergic Receptors Improve Motor Symptoms in Patients with Parkinson’s Disease? The Potential Role of Zolpidem in the Treatment of Parkinson’s Disease. Park. Dis. 2016, 2016, 2531812. [Google Scholar] [CrossRef]
- Collins-Praino, L.E.; Paul, N.E.; Rychalsky, K.L.; Hinman, J.R.; Chrobak, J.J.; Senatus, P.B.; Salamone, J.D. Pharmacological and Physiological Characterization of the Tremulous Jaw Movement Model of Parkinsonian Tremor: Potential Insights into the Pathophysiology of Tremor. Front. Syst. Neurosci. 2011, 5, 49. [Google Scholar] [CrossRef] [Green Version]
- Podurgiel, S.J.; Yohn, S.E.; Dortche, K.; Correa, M.; Salamone, J.D. The MAO-B Inhibitor Deprenyl Reduces the Oral Tremor and the Dopamine Depletion Induced by the VMAT-2 Inhibitor Tetrabenazine. Behav. Brain Res. 2016, 298, 188–191. [Google Scholar] [CrossRef]
- Collins-Praino, L.E.; Paul, N.E.; Ledgard, F.; Podurgiel, S.J.; Kovner, R.; Baqi, Y.; Müller, C.E.; Senatus, P.B.; Salamone, J.D. Deep Brain Stimulation of the Subthalamic Nucleus Reverses Oral Tremor in Pharmacological Models of Parkinsonism: Interaction with the Effects of Adenosine A2A Antagonism. Eur. J. Neurosci. 2013, 38, 2183–2191. [Google Scholar] [CrossRef] [PubMed]
- Cousins, M.S.; Carriero, D.L.; Salamone, J.D. Tremulous Jaw Movements Induced by the Acetylcholinesterase Inhibitor Tacrine: Effects of Antiparkinsonian Drugs. Eur. J. Pharmacol. 1997, 322, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Salamone, J.D.; Carlson, B.B.; Rios, C.; Lentini, E.; Correa, M.; Wisniecki, A.; Betz, A. Dopamine Agonists Suppress Cholinomimetic-Induced Tremulous Jaw Movements in an Animal Model of Parkinsonism: Tremorolytic Effects of Pergolide, Ropinirole and CY 208–243. Behav. Brain Res. 2005, 156, 173–179. [Google Scholar] [CrossRef]
- Cousins, M.S.; Finn, M.; Trevitt, J.; Carriero, D.L.; Conlan, A.; Salamone, J.D. The Role of Ventrolateral Striatal Acetylcholine in the Production of Tacrine-Induced Jaw Movements. Pharmacol. Biochem. Behav. 1999, 62, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Salamone, J.D.; Betz, A.J.; Ishiwari, K.; Felsted, J.; Madson, L.; Mirante, B.; Clark, K.; Font, L.; Korbey, S.; Sager, T.N.; et al. Tremorolytic Effects of Adenosine A2A Antagonists: Implications for Parkinsonism. Front. Biosci. 2008, 13, 3594–3605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisniecki, A.; Correa, M.; Arizzi, M.N.; Ishiwari, K.; Salamone, J.D. Motor Effects of GABA(A) Antagonism in Globus Pallidus: Studies of Locomotion and Tremulous Jaw Movements in Rats. Psychopharmacology 2003, 170, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Kosmowska, B.; Ossowska, K.; Wardas, J. Blockade of Adenosine A2A Receptors Inhibits Tremulous Jaw Movements as Well as Expression of Zif-268 and GAD65 MRNAs in Brain Motor Structures. Behav. Brain Res. 2022, 417, 113585. [Google Scholar] [CrossRef]
- Shahed, J.; Jankovic, J. Exploring the Relationship between Essential Tremor and Parkinson’s Disease. Park. Relat. Disord. 2007, 13, 67–76. [Google Scholar] [CrossRef]
- Lenka, A.; Benito-León, J.; Louis, E.D. Is There a Premotor Phase of Essential Tremor? Tremor. Other Hyperkinet. Mov. 2017, 7, 498. [Google Scholar] [CrossRef]
- Labiano-Fontcuberta, A.; Benito-León, J. [Essential Tremor and Parkinson’s Disease: Are They Associated?]. Rev. Neurol. 2012, 55, 479–489. [Google Scholar]
- Amrutkar, D.V.; Dyhring, T.; Jacobsen, T.A.; Larsen, J.S.; Sandager-Nielsen, K. Anti-Tremor Action of Subtype Selective Positive Allosteric Modulators of GABAA Receptors in a Rat Model of Essential Tremors. Cerebellum 2020, 19, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Assini, R.; Abercrombie, E.D. Zolpidem Ameliorates Motor Impairments in the Unilaterally 6-Hydroxydopamine-Lesioned Rat. Eur. J. Neurosci. 2018, 48, 1896–1905. [Google Scholar] [CrossRef] [PubMed]
- Collins-Praino, L.E.; Podurgiel, S.J.; Kovner, R.; Randall, P.A.; Salamone, J.D. Extracellular GABA in Globus Pallidus Increases during the Induction of Oral Tremor by Haloperidol but Not by Muscarinic Receptor Stimulation. Behav. Brain Res. 2012, 234, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Kosmowska, B.; Ossowska, K.; Wardas, J. Pramipexole Reduces Zif-268 MRNA Expression in Brain Structures Involved in the Generation of Harmaline-Induced Tremor. Neurochem. Res. 2020, 45, 1518–1525. [Google Scholar] [CrossRef] [Green Version]
- Kralic, J.E. Molecular and Pharmacological Characterization of GABAA Receptor Alpha 1 Subunit Knockout Mice. J. Pharmacol. Exp. Ther. 2002, 302, 1037–1045. [Google Scholar] [CrossRef] [Green Version]
- Cerne, R.; Lippa, A.; Poe, M.M.; Smith, J.L.; Jin, X.; Ping, X.; Golani, L.K.; Cook, J.M.; Witkin, J.M. GABAkines–Advances in the Discovery, Development, and Commercialization of Positive Allosteric Modulators of GABAA Receptors. Pharmacol. Ther. 2022, 234, 108035. [Google Scholar] [CrossRef]
- Sieghart, W.; Savic, M.M. International Union of Basic and Clinical Pharmacology. CVI: GABAA Receptor Subtype-and Function-Selective Ligands: Key Issues in Translation to Humans. Pharmacol. Rev. 2018, 70, 836–878. [Google Scholar] [CrossRef] [Green Version]
- de Lucas, A.G.; Ahring, P.K.; Larsen, J.S.; Rivera-Arconada, I.; Lopez-Garcia, J.A.; Mirza, N.R.; Munro, G. GABAA A5 Subunit-Containing Receptors Do Not Contribute to Reversal of Inflammatory-Induced Spinal Sensitization as Indicated by the Unique Selectivity Profile of the GABAA Receptor Allosteric Modulator NS16085. Biochem. Pharmacol. 2015, 93, 370–379. [Google Scholar] [CrossRef]
- Witkin, J.M.; Lippa, A.; Smith, J.L.; Jin, X.; Ping, X.; Biggerstaff, A.; Kivell, B.M.; Knutson, D.E.; Sharmin, D.; Pandey, K.P.; et al. The Imidazodiazepine, KRM-II-81: An Example of a Newly Emerging Generation of GABAkines for Neurological and Psychiatric Disorders. Pharmacol. Biochem. Behav. 2022, 213, 173321. [Google Scholar] [CrossRef]
- Fischer, B.; Schlitt, R.; Hamade, B.; Rehman, S.; Ernst, M.; Poe, M.; Li, G.; Kodali, R.; Arnold, L.; Cook, J. Pharmacological and Antihyperalgesic Properties of the Novel A2/3 Preferring GABAA Receptor Ligand MP-III-024. Brain Res. Bull. 2017, 131, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Podurgiel, S.J.; Nunes, E.J.; Yohn, S.E.; Barber, J.; Thompson, A.; Milligan, M.; Lee, C.A.; López-Cruz, L.; Pardo, M.; Valverde, O.; et al. The Vesicular Monoamine Transporter (VMAT-2) Inhibitor Tetrabenazine Induces Tremulous Jaw Movements in Rodents: Implications for Pharmacological Models of Parkinsonian Tremor. Neuroscience 2013, 250, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Yohn, S.E.; Thompson, C.; Randall, P.A.; Lee, C.A.; Müller, C.E.; Baqi, Y.; Correa, M.; Salamone, J.D. The VMAT-2 Inhibitor Tetrabenazine Alters Effort-Related Decision Making as Measured by the T-Maze Barrier Choice Task: Reversal with the Adenosine A2A Antagonist MSX-3 and the Catecholamine Uptake Blocker Bupropion. Psychopharmacology 2015, 232, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Ishiwari, K.; Betz, A.; Weber, S.; Felsted, J.; Salamone, J.D. Validation of the Tremulous Jaw Movement Model for Assessment of the Motor Effects of Typical and Atypical Antipychotics: Effects of Pimozide (Orap) in Rats. Pharmacol. Biochem. Behav. 2005, 80, 351–362. [Google Scholar] [CrossRef]
- Fajkis, N.; Marcinkowska, M.; Gryzło, B.; Krupa, A.; Kolaczkowski, M. Study on a Three-Step Rapid Assembly of Zolpidem and Its Fluorinated Analogues Employing Microwave-Assisted Chemistry. Molecules 2020, 25, 3161. [Google Scholar] [CrossRef]
- Murphy, H.M.; Ihekoronze, C.; Wideman, C.H. Zolpidem-Induced Changes in Activity, Metabolism, and Anxiety in Rats. Pharmacol. Biochem. Behav. 2011, 98, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Mierzejewski, P.; Kolaczkowski, M.; Marcinkowska, M.; Wesolowska, A.; Samochowiec, J.; Pawlowski, M.; Bienkowski, P. Antipsychotic-like Effects of Zolpidem in Wistar Rats. Eur. J. Pharmacol. 2016, 773, 51–58. [Google Scholar] [CrossRef]
- Poe, M.M. Synthesis of Subtype Selective Bz/GABAA Receptor Ligands for the Treatment of Anxiety, Epilepsy and Neuropathic Pain, As Well As Schizophrenia and Asthma. Ph.D. Thesis, University of Wisconsin-Milwaukee, Milwaukee, WI, USA, 2016. [Google Scholar]
- Rahman, M.A.; Keck, T.M.; Poe, M.M.; Sharmin, D.; Cook, J.M.; Fischer, B.D. Synergistic Antihyperalgesic and Antinociceptive Effects of Morphine and Methyl 8-Ethynyl-6-(Pyridin-2-Yl)-4H-Benzo[f]Imidazo[1,5-a][1,4]Diazepine-3-Carboxylate (MP-III-024): A Positive Allosteric Modulator at A2GABAA and A3GABAA Receptors. Psychopharmacology 2021, 238, 1585–1592. [Google Scholar] [CrossRef]
- Kosmowska, B.; Ossowska, K.; Konieczny, J.; Lenda, T.; Berghauzen-Maciejewska, K.; Wardas, J. Inhibition of Excessive Glutamatergic Transmission in the Ventral Thalamic Nuclei by a Selective Adenosine A1 Receptor Agonist, 5′-Chloro-5′-Deoxy-(±)-ENBA Underlies Its Tremorolytic Effect in the Harmaline-Induced Model of Essential Tremor. Neuroscience 2020, 429, 106–118. [Google Scholar] [CrossRef]
- Ossowska, K.; Głowacka, U.; Kosmowska, B.; Wardas, J. Apomorphine Enhances Harmaline-Induced Tremor in Rats. Pharmacol. Rep. 2015, 67, 435–441. [Google Scholar] [CrossRef]
- Sequeira, A.; Shen, K.; Gottlieb, A.; Limon, A. Human Brain Transcriptome Analysis Finds Region- and Subject-Specific Expression Signatures of GABAAR Subunits. Commun. Biol. 2019, 2, 153. [Google Scholar] [CrossRef] [Green Version]
- de Haas, S.L.; Zoethout, R.W.M.; van Dyck, K.; de Smet, M.; Rosen, L.B.; Murphy, M.G.; Gottesdiener, K.M.; Schoemaker, R.C.; Cohen, A.F.; van Gerven, J.M.A. The Effects of TPA023, a GABA Aα2,3 Subtype-Selective Partial Agonist, on Essential Tremor in Comparison to Alcohol. J. Psychopharmacol. 2012, 26, 282–291. [Google Scholar] [CrossRef]
- Atack, J.R. GABAA Receptor Subtype-Selective Modulators. I. A2/A3-Selective Agonists as Non-Sedating Anxiolytics. Curr. Top. Med. Chem. 2011, 11, 1176–1202. [Google Scholar] [CrossRef] [PubMed]
- Gołembiowska, K.; Berghauzen-Maciejewska, K.; Górska, A.; Kamińska, K.; Ossowska, K. A Partial Lesion of the Substantia Nigra Pars Compacta and Retrorubral Field Decreases the Harmaline-Induced Glutamate Release in the Rat Cerebellum. Brain Res. 2013, 1537, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Beitz, A.J.; Saxon, D. Harmaline-Induced Climbing Fiber Activation Causes Amino Acid and Peptide Release in the Rodent Cerebellar Cortex and a Unique Temporal Pattern of Fos Expression in the Olivo-Cerebellar Pathway. J. Neurocytol. 2004, 33, 49–74. [Google Scholar] [CrossRef] [PubMed]
- Daniele, A.; Albanese, A.; Gainotti, G.; Gregori, B.; Bartolomeo, P. Zolpidem in Parkinson’s Disease. Lancet 1997, 349, 1222–1223. [Google Scholar] [CrossRef]
- Farver, D.K.; Khan, M.H. Zolpidem for Antipsychotic-Induced Parkinsonism. Ann. Pharmacother. 2001, 35, 435–437. [Google Scholar] [CrossRef]
- Růzicka, E.; Roth, J.; Jech, R.; Busek, P. Subhypnotic Doses of Zolpidem Oppose Dopaminergic-Induced Dyskinesia in Parkinson’s Disease. Mov. Disord. 2000, 15, 734–735. [Google Scholar] [CrossRef]
- Hall, S.D.; Prokic, E.J.; McAllister, C.J.; Ronnqvist, K.C.; Williams, A.C.; Yamawaki, N.; Witton, C.; Woodhall, G.L.; Stanford, I.M. GABA-Mediated Changes in Inter-Hemispheric Beta Frequency Activity in Early-Stage Parkinson’s Disease. Neuroscience 2014, 281, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Diamond, J.M.; Lopes, M.B.; Elias, W.J.; Jansen, L.A. Gamma-Aminobutyric Acid A Receptor Subunit Expression and Cellular Localization in the Human Parkinsonian Globus Pallidus. World Neurosurg. 2022, 165, e159–e168. [Google Scholar] [CrossRef]
- Laifenfeld, D.; Yanover, C.; Ozery-Flato, M.; Shaham, O.; Rosen-Zvi, M.; Lev, N.; Goldschmidt, Y.; Grossman, I. Emulated Clinical Trials from Longitudinal Real-World Data Efficiently Identify Candidates for Neurological Disease Modification: Examples from Parkinson’s Disease. Front. Pharmacol. 2021, 12, 631584. [Google Scholar] [CrossRef]
- Prokic, E.J.; Stanford, I.M.; Woodhall, G.L.; Williams, A.C.; Hall, S.D. Bradykinesia Is Driven by Cumulative Beta Power during Continuous Movement and Alleviated by Gabaergic Modulation in Parkinson’s Disease. Front. Neurol. 2019, 10, 1298. [Google Scholar] [CrossRef] [PubMed]
- Fois, A.F.; Chang, F.C.; Barnett, R.; London, K.; Mahant, N.; Ha, A.; Aldaajani, Z.; Cruse, B.; Morales-Briceno, H.; Ma, K.K.; et al. Rest Tremor Correlates with Reduced Contralateral Striatal Dopamine Transporter Binding in Parkinson’s Disease. Park. Relat. Disord. 2021, 85, 102–108. [Google Scholar] [CrossRef]
- Katz, J.; Nielsen, K.M.; Soghomonian, J.-J. Comparative Effects of Acute or Chronic Administration of Levodopa to 6-Hydroxydopamine-Lesioned Rats on the Expression of Glutamic Acid Decarboxylase in the Neostriatum and GABAA Receptors Subunits in the Substantia Nigra, Pars Reticulata. Neuroscience 2005, 132, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Zimprich, F.; Zezula, J.; Sieghart, W.; Lassmann, H. Immunohistochemical Localization of the A1, A2 and A3 Subunit of the GABAA Receptor in the Rat Brain. Neurosci. Lett. 1991, 127, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Hörtnagl, H.; Tasan, R.O.; Wieselthaler, A.; Kirchmair, E.; Sieghart, W.; Sperk, G. Patterns of MRNA and Protein Expression for 12 GABAA Receptor Subunits in the Mouse Brain. Neuroscience 2013, 236, 345–372. [Google Scholar] [CrossRef] [Green Version]
- Laurie, D.; Seeburg, P.; Wisden, W. The Distribution of 13 GABAA Receptor Subunit MRNAs in the Rat Brain. II. Olfactory Bulb and Cerebellum. J. Neurosci. 1992, 12, 1063–1076. [Google Scholar] [CrossRef]
- Pirker, S.; Schwarzer, C.; Wieselthaler, A.; Sieghart, W.; Sperk, G. GABAA Receptors: Immunocytochemical Distribution of 13 Subunits in the Adult Rat Brain. Neuroscience 2000, 101, 815–850. [Google Scholar] [CrossRef]
- Wisden, W.; Laurie, D.J.; Monyer, H.; Seeburg, P.H. The Distribution of 13 GABAA Receptor Subunit MRNAs in the Rat Brain. I. Telencephalon, Diencephalon, Mesencephalon. J. Neurosci. 1992, 12, 1040–1062. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosmowska, B.; Paleczna, M.; Biała, D.; Kadłuczka, J.; Wardas, J.; Witkin, J.M.; Cook, J.M.; Sharmin, D.; Marcinkowska, M.; Kuter, K.Z. GABA-A Alpha 2/3 but Not Alpha 1 Receptor Subunit Ligand Inhibits Harmaline and Pimozide-Induced Tremor in Rats. Biomolecules 2023, 13, 197. https://doi.org/10.3390/biom13020197
Kosmowska B, Paleczna M, Biała D, Kadłuczka J, Wardas J, Witkin JM, Cook JM, Sharmin D, Marcinkowska M, Kuter KZ. GABA-A Alpha 2/3 but Not Alpha 1 Receptor Subunit Ligand Inhibits Harmaline and Pimozide-Induced Tremor in Rats. Biomolecules. 2023; 13(2):197. https://doi.org/10.3390/biom13020197
Chicago/Turabian StyleKosmowska, Barbara, Martyna Paleczna, Dominika Biała, Justyna Kadłuczka, Jadwiga Wardas, Jeffrey M. Witkin, James M. Cook, Dishary Sharmin, Monika Marcinkowska, and Katarzyna Z. Kuter. 2023. "GABA-A Alpha 2/3 but Not Alpha 1 Receptor Subunit Ligand Inhibits Harmaline and Pimozide-Induced Tremor in Rats" Biomolecules 13, no. 2: 197. https://doi.org/10.3390/biom13020197
APA StyleKosmowska, B., Paleczna, M., Biała, D., Kadłuczka, J., Wardas, J., Witkin, J. M., Cook, J. M., Sharmin, D., Marcinkowska, M., & Kuter, K. Z. (2023). GABA-A Alpha 2/3 but Not Alpha 1 Receptor Subunit Ligand Inhibits Harmaline and Pimozide-Induced Tremor in Rats. Biomolecules, 13(2), 197. https://doi.org/10.3390/biom13020197