

Role of Insect and Mammal Glutathione Transferases in Chemoperception

 , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. The Involvement of GSTs in Mammalian Chemoperception
2.1. Chemoperception in Mammals
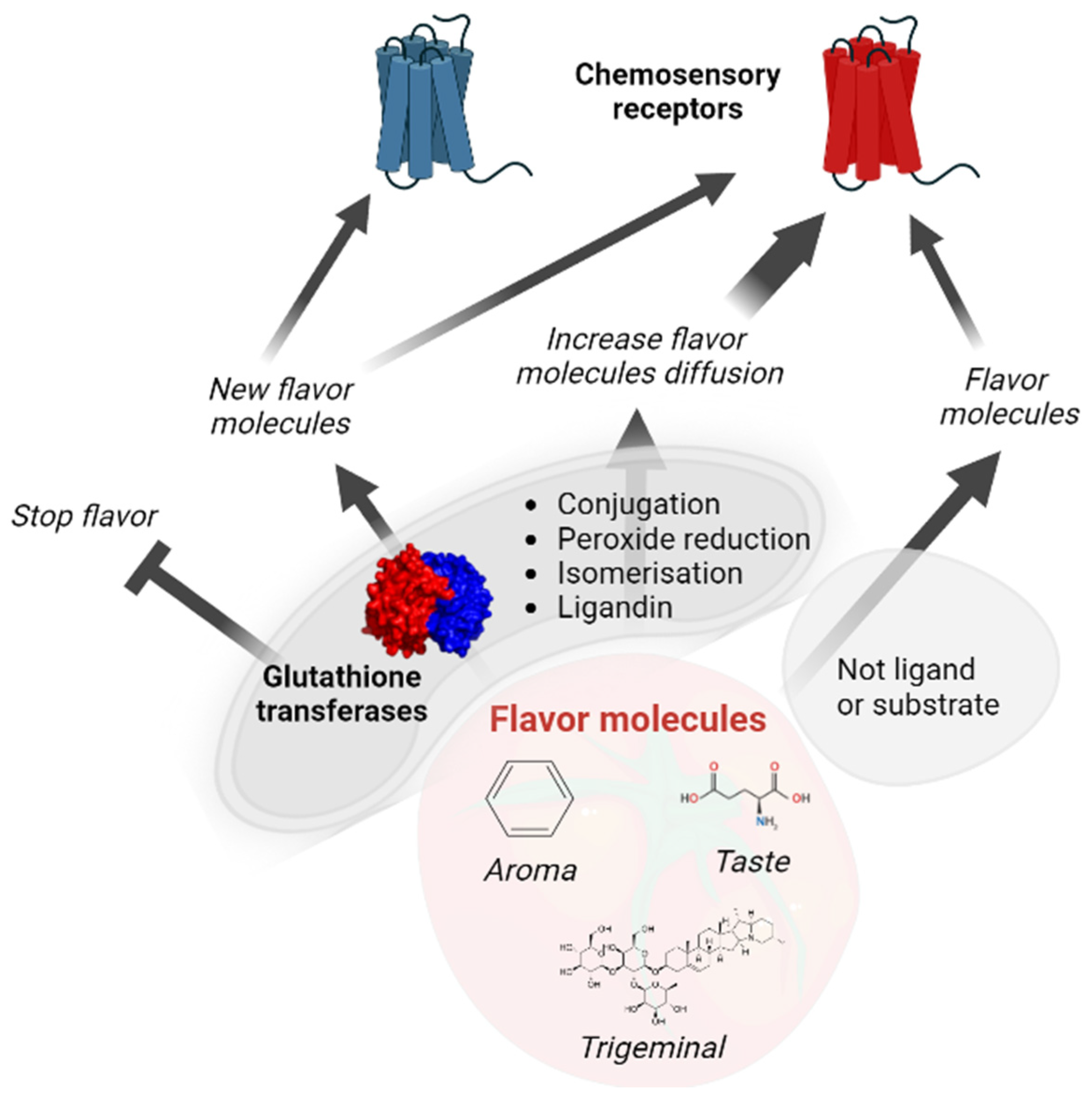
2.2. Roles of GSTs in Mammalian Chemoperception

{kind=link}
| Mammal Species | Location | GST Classes | Ref. |
|---|---|---|---|
| Canis lupus familiaris | Saliva | Alpha, Mu, and Omega | [56] |
| Equus caballus | Saliva | Pi | [57] |
| Homo sapiens | Olfactory mucus | Alpha and Pi | [58,59] |
| Olfactory epithelium | Alpha, Mu, and Pi | [43] | |
| Saliva | Alpha, Kappa, Mu Omega, Theta, and Pi | [60,61] | |
| Mus musculus | Sensory cilia | Alpha, Kappa, Mu, Omega, Tau, and Zeta | [62] |
| Saliva | Omega | [63] | |
| Oryctolagus cuniculus | Olfactory mucus | Alpha, Mu, and Pi | [49] |
| Ovis aries | Saliva | Alpha | [57] |
| Rattus norvegicus | Sensory cilia | Alpha and Mu | [64] |
| Olfactory epithelium | Alpha, Mu, and Pi | [4,41] | |
| Olfactory mucus | Alpha, Mu, and Pi | [4] |
3. The Involvement of GSTs in Insect Chemoperception
3.1. Chemoperception in Insects
3.2. Roles of GSTs in Insect Chemoperception
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Valen, L. A new evolutionary law. Evol. Theory 1973, 1, 1–30. [Google Scholar]
- Booth, J.; Boyland, E.; Sims, P. An enzyme from rat liver catalysing conjugations with glutathione. Biochem. J. 1961, 79, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Mannervik, B. Five decades with glutathione and the GSTome. J. Biol. Chem. 2012, 287, 6072–6083. [Google Scholar] [CrossRef]
- Heydel, J.M.; Menetrier, F.; Belloir, C.; Canon, F.; Faure, P.; Lirussi, F.; Chavanne, E.; Saliou, J.M.; Artur, Y.; Canivenc-Lavier, M.C.; et al. Characterization of rat glutathione transferases in olfactory epithelium and mucus. PLoS ONE 2019, 14, e0220259. [Google Scholar] [CrossRef]
- Gonzalez, D.; Fraichard, S.; Grassein, P.; Delarue, P.; Senet, P.; Nicolai, A.; Chavanne, E.; Mucher, E.; Artur, Y.; Ferveur, J.F.; et al. Characterization of a Drosophila glutathione transferase involved in isothiocyanate detoxification. Insect Biochem. Mol. Biol. 2018, 95, 33–43. [Google Scholar] [CrossRef]
- Hayes, J.D.; Strange, R.C. Glutathione S-transferase polymorphisms and their biological consequences. Pharmacology 2000, 61, 154–166. [Google Scholar] [CrossRef]
- Morel, F.; Aninat, C. The glutathione transferase kappa family. Drug Metab. Rev. 2011, 43, 281–291. [Google Scholar] [CrossRef]
- Ladner, J.E.; Parsons, J.F.; Rife, C.L.; Gilliland, G.L.; Armstrong, R.N. Parallel evolutionary pathways for glutathione transferases: Structure and mechanism of the mitochondrial class kappa enzyme rGSTK1-1. Biochemistry 2004, 43, 352–361. [Google Scholar] [CrossRef]
- Roncalli, V.; Cieslak, M.C.; Passamaneck, Y.; Christie, A.E.; Lenz, P.H. Glutathione S-Transferase (GST) Gene Diversity in the Crustacean Calanus finmarchicus--Contributors to Cellular Detoxification. PLoS ONE 2015, 10, e0123322. [Google Scholar] [CrossRef]
- Hurst, R.; Bao, Y.; Jemth, P.; Mannervik, B.; Williamson, G. Phospholipid hydroperoxide glutathione peroxidase activity of human glutathione transferases. Biochem. J. 1998, 332 Pt 1, 97–100. [Google Scholar] [CrossRef]
- Clark, A.G.; Shamaan, N.A. Evidence that DDT-dehydrochlorinase from the house fly is a glutathione S-transferase. Pestic. Biochem. Physiol. 1984, 22, 249–261. [Google Scholar] [CrossRef]
- Johansson, A.S.; Mannervik, B. Human glutathione transferase A3-3, a highly efficient catalyst of double-bond isomerization in the biosynthetic pathway of steroid hormones. J. Biol. Chem. 2001, 276, 33061–33065. [Google Scholar] [CrossRef]
- Škerlová, J.; Lindström, H.; Gonis, E.; Sjödin, B.; Neiers, F.; Stenmark, P.; Mannervik, B. Structure and steroid isomerase activity of Drosophila glutathione transferase E14 essential for ecdysteroid biosynthesis. FEBS Lett. 2019, 594, 1187–1195. [Google Scholar] [CrossRef]
- Kamisaka, K.; Gatmaitan, Z.; Moore, C.L.; Arias, I.M. Ligandin Reverses Bilirubin Inhibition of Liver Mitochondrial Respiration In Vitro. Pediatr. Res. 1975, 9, 903–905. [Google Scholar]
- Mannervik, B.; Board, P.G.; Hayes, J.D.; Listowsky, I.; Pearson, W.R. Nomenclature for mammalian soluble glutathione transferases. Methods Enzymol. 2005, 401, 1–8. [Google Scholar] [CrossRef]
- Sherratt, P.J.; Hayes, J.D. 9 Glutathione S-Transferases; Costas, I., Ed.; John Wiley and Sons: Hoboken, NJ, USA, 2001. [Google Scholar] [CrossRef]
- Gonis, E.; Fraichard, S.; Chertemps, T.; Hecker, A.; Schwartz, M.; Canon, F.; Neiers, F. Expression Patterns of Drosophila Melanogaster Glutathione Transferases. Insects 2022, 13, 612. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, Z.; Lin, X.; Feng, Q.; Zheng, S. Structure and expression of glutathione S-transferase genes from the midgut of the Common cutworm, Spodoptera litura (Noctuidae) and their response to xenobiotic compounds and bacteria. J. Insect Physiol. 2011, 57, 1033–1044. [Google Scholar] [CrossRef]
- Knight, T.R.; Choudhuri, S.; Klaassen, C.D. Constitutive mRNA expression of various glutathione S-transferase isoforms in different tissues of mice. Toxicol. Sci. 2007, 100, 513–524. [Google Scholar] [CrossRef]
- Sullivan, S.L. Mammalian chemosensory receptors. Neuroreport 2002, 13, A9–A17. [Google Scholar] [CrossRef]
- Filiou, R.P.; Lepore, F.; Bryant, B.; Lundstrom, J.N.; Frasnelli, J. Perception of trigeminal mixtures. Chem. Senses 2015, 40, 61–69. [Google Scholar] [CrossRef]
- Su, C.Y.; Menuz, K.; Carlson, J.R. Olfactory perception: Receptors, cells, and circuits. Cell 2009, 139, 45–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malnic, B.; Hirono, J.; Sato, T.; Buck, L.B. Combinatorial receptor codes for odors. Cell 1999, 96, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Yarmolinsky, D.A.; Zuker, C.S.; Ryba, N.J. Common sense about taste: From mammals to insects. Cell 2009, 139, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Dalziel, J.E. G Protein-Coupled Receptors in Taste Physiology and Pharmacology. Front. Pharmacol. 2020, 11, 587664. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, L.; Lametsch, R. Current progress in kokumi-active peptides, evaluation and preparation methods: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 1230–1241. [Google Scholar] [CrossRef]
- Toelstede, S.; Hofmann, T. Kokumi-active glutamyl peptides in cheeses and their biogeneration by Penicillium roquefortii. J. Agric. Food Chem. 2009, 57, 3738–3748. [Google Scholar] [CrossRef]
- Maruyama, Y.; Yasuda, R.; Kuroda, M.; Eto, Y. Kokumi substances, enhancers of basic tastes, induce responses in calcium-sensing receptor expressing taste cells. PLoS ONE 2012, 7, e34489. [Google Scholar] [CrossRef]
- Canon, F.; Belloir, C.; Bourillot, E.; Brignot, H.; Briand, L.; Feron, G.; Lesniewska, E.; Nivet, C.; Septier, C.; Schwartz, M.; et al. Perspectives on Astringency Sensation: An Alternative Hypothesis on the Molecular Origin of Astringency. J. Agric. Food Chem. 2021, 69, 3822–3826. [Google Scholar] [CrossRef]
- Boichot, V.; Muradova, M.; Nivet, C.; Proskura, A.; Heydel, J.M.; Canivenc-Lavier, M.C.; Canon, F.; Neiers, F.; Schwartz, M. The role of perirecetor events in flavor perception. Front. Food Sci. Technol. 2022, 2, 989291. [Google Scholar] [CrossRef]
- Sarkar, M.A. Drug metabolism in the nasal mucosa. Pharm. Res. 1992, 9, 1–9. [Google Scholar] [CrossRef]
- Heydel, J.M.; Faure, P.; Neiers, F. Nasal odorant metabolism: Enzymes, activity and function in olfaction. Drug Metab. Rev. 2019, 51, 224–245. [Google Scholar] [CrossRef]
- Mallery, S.R.; Budendorf, D.E.; Larsen, M.P.; Pei, P.; Tong, M.; Holpuch, A.S.; Larsen, P.E.; Stoner, G.D.; Fields, H.W.; Chan, K.K.; et al. Effects of human oral mucosal tissue, saliva, and oral microflora on intraoral metabolism and bioactivation of black raspberry anthocyanins. Cancer Prev. Res. 2011, 4, 1209–1221. [Google Scholar] [CrossRef]
- Schwartz, M.; Neiers, F.; Charles, J.P.; Heydel, J.M.; Munoz-Gonzalez, C.; Feron, G.; Canon, F. Oral enzymatic detoxification system: Insights obtained from proteome analysis to understand its potential impact on aroma metabolization. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5516–5547. [Google Scholar] [CrossRef]
- Canon, F.; Neiers, F.; Guichard, E. Saliva and Flavor Perception: Perspectives. J. Agric. Food Chem. 2018, 66, 7873–7879. [Google Scholar] [CrossRef]
- Munoz-Gonzalez, C.; Brule, M.; Martin, C.; Feron, G.; Canon, F. Molecular mechanisms of aroma persistence: From noncovalent interactions between aroma compounds and the oral mucosa to metabolization of aroma compounds by saliva and oral cells. Food Chem. 2022, 373, 131467. [Google Scholar] [CrossRef]
- Robert-Hazotte, A.; Faure, P.; Menetrier, F.; Folia, M.; Schwartz, M.; Le Quere, J.L.; Neiers, F.; Thomas-Danguin, T.; Heydel, J.M. Nasal Odorant Competitive Metabolism Is Involved in the Human Olfactory Process. J. Agric. Food Chem. 2022, 70, 8385–8394. [Google Scholar] [CrossRef]
- Ployon, S.; Brule, M.; Andriot, I.; Morzel, M.; Canon, F. Understanding retention and metabolization of aroma compounds using an in vitro model of oral mucosa. Food Chem. 2020, 318, 126468. [Google Scholar] [CrossRef]
- Mannervik, B.; Alin, P.; Guthenberg, C.; Jensson, H.; Tahir, M.K.; Warholm, M.; Jornvall, H. Identification of three classes of cytosolic glutathione transferase common to several mammalian species: Correlation between structural data and enzymatic properties. Proc. Natl. Acad. Sci. USA 1985, 82, 7202–7206. [Google Scholar] [CrossRef]
- Sheehan, D.; Meade, G.; Foley, V.M.; Dowd, C.A. Structure, function and evolution of glutathione transferases: Implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem. J. 2001, 360, 1–16. [Google Scholar] [CrossRef]
- Ben-Arie, N.; Khen, M.; Lancet, D. Glutathione S-transferases in rat olfactory epithelium: Purification, molecular properties and odorant biotransformation. Biochem. J. 1993, 292 Pt 2, 379–384. [Google Scholar] [CrossRef]
- Krishna, N.S.; Getchell, T.V.; Getchell, M.L. Differential expression of alpha, mu, and pi classes of glutathione S-transferases in chemosensory mucosae of rats during development. Cell Tissue Res. 1994, 275, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.; Menetrier, F.; Heydel, J.M.; Chavanne, E.; Faure, P.; Labrousse, M.; Lirussi, F.; Canon, F.; Mannervik, B.; Briand, L.; et al. Interactions Between Odorants and Glutathione Transferases in the Human Olfactory Cleft. Chem. Senses 2020, 45, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Toyoshima, K.; Tandler, B. Dividing type II cell in rabbit taste bud. Anat Rec. 1986, 214, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.M.; Cox, M.E.; Solomons, N. Zinc and the special senses. Ann. Intern. Med. 1983, 99, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Kudo, H.; Doi, Y.; Nishino, T.; Nara, S.; Hamasaki, K.; Fujimoto, S. Dietary zinc deficiency decreases glutathione S-transferase expression in the rat olfactory epithelium. J. Nutr. 2000, 130, 38–44. [Google Scholar] [CrossRef]
- Schaal, B.; Coureaud, G.; Langlois, D.; Ginies, C.; Semon, E.; Perrier, G. Chemical and behavioural characterization of the rabbit mammary pheromone. Nature 2003, 424, 68–72. [Google Scholar] [CrossRef]
- Legendre, A.; Faure, P.; Tiesset, H.; Potin, C.; Jakob, I.; Sicard, G.; Schaal, B.; Artur, Y.; Coureaud, G.; Heydel, J.M. When the nose must remain responsive: Glutathione conjugation of the mammary pheromone in the newborn rabbit. Chem. Senses 2014, 39, 425–437. [Google Scholar] [CrossRef]
- Robert-Hazotte, A.; Faure, P.; Neiers, F.; Potin, C.; Artur, Y.; Coureaud, G.; Heydel, J.M. Nasal mucus glutathione transferase activity and impact on olfactory perception and neonatal behavior. Sci. Rep. 2019, 9, 3104. [Google Scholar] [CrossRef]
- Hanser, H.I.; Faure, P.; Robert-Hazotte, A.; Artur, Y.; Duchamp-Viret, P.; Coureaud, G.; Heydel, J.M. Odorant-odorant metabolic interaction, a novel actor in olfactory perception and behavioral responsiveness. Sci. Rep. 2017, 7, 10219. [Google Scholar] [CrossRef]
- Nishino, T.; Kudo, H.; Doi, Y.; Maeda, M.; Hamasaki, K.; Morita, M.; Fujimoto, S. Immunocytochemistry of glutathione S-transferase in taste bud cells of rat circumvallate and foliate papillae. Chem. Senses 2001, 26, 179–188. [Google Scholar] [CrossRef]
- Fabrini, R.; Bocedi, A.; Camerini, S.; Fusetti, M.; Ottaviani, F.; Passali, F.M.; Topazio, D.; Iavarone, F.; Francia, I.; Castagnola, M.; et al. Inactivation of human salivary glutathione transferase P1-1 by hypothiocyanite: A post-translational control system in search of a role. PLoS ONE 2014, 9, e112797. [Google Scholar] [CrossRef] [Green Version]
- Derindag, G.; Akgul, H.M.; Kiziltunc, A.; Ozkan, H.I.; Kiziltunc Ozmen, H.; Akgul, N. Evaluation of saliva glutathione, glutathione peroxidase, and malondialdehyde levels in head-neck radiotherapy patients. Turk J. Med. Sci. 2021, 51, 644–649. [Google Scholar] [CrossRef]
- Sreerama, L.; Hedge, M.W.; Sladek, N.E. Identification of a class 3 aldehyde dehydrogenase in human saliva and increased levels of this enzyme, glutathione S-transferases, and DT-diaphorase in the saliva of subjects who continually ingest large quantities of coffee or broccoli. Clin. Cancer Res. 1995, 1, 1153–1163. [Google Scholar]
- Schwartz, M.; Brignot, H.; Feron, G.; Hummel, T.; Zhu, Y.; von Koskull, D.; Heydel, J.M.; Lirussi, F.; Canon, F.; Neiers, F. Role of human salivary enzymes in bitter taste perception. Food Chem. 2022, 386, 132798. [Google Scholar] [CrossRef]
- Franco-Martinez, L.; Gelemanovic, A.; Horvatic, A.; Contreras-Aguilar, M.D.; Dabrowski, R.; Mrljak, V.; Ceron, J.J.; Martinez-Subiela, S.; Tvarijonaviciute, A. Changes in Serum and Salivary Proteins in Canine Mammary Tumors. Animals 2020, 10, 741. [Google Scholar] [CrossRef]
- de Sousa-Pereira, P.; Cova, M.; Abrantes, J.; Ferreira, R.; Trindade, F.; Barros, A.; Gomes, P.; Colaco, B.; Amado, F.; Esteves, P.J.; et al. Cross-species comparison of mammalian saliva using an LC-MALDI based proteomic approach. Proteomics 2015, 15, 1598–1607. [Google Scholar] [CrossRef]
- Debat, H.; Eloit, C.; Blon, F.; Sarazin, B.; Henry, C.; Huet, J.C.; Trotier, D.; Pernollet, J.C. Identification of human olfactory cleft mucus proteins using proteomic analysis. J. Proteome Res. 2007, 6, 1985–1996. [Google Scholar] [CrossRef]
- Yoshikawa, K.; Wang, H.; Jaen, C.; Haneoka, M.; Saito, N.; Nakamura, J.; Adappa, N.D.; Cohen, N.A.; Dalton, P. The human olfactory cleft mucus proteome and its age-related changes. Sci. Rep. 2018, 8, 17170. [Google Scholar] [CrossRef]
- Pappa, E.; Vastardis, H.; Mermelekas, G.; Gerasimidi-Vazeou, A.; Zoidakis, J.; Vougas, K. Saliva Proteomics Analysis Offers Insights on Type 1 Diabetes Pathology in a Pediatric Population. Front. Physiol. 2018, 9, 444. [Google Scholar] [CrossRef]
- Grassl, N.; Kulak, N.A.; Pichler, G.; Geyer, P.E.; Jung, J.; Schubert, S.; Sinitcyn, P.; Cox, J.; Mann, M. Ultra-deep and quantitative saliva proteome reveals dynamics of the oral microbiome. Genome Med. 2016, 8, 44. [Google Scholar] [CrossRef]
- Kuhlmann, K.; Tschapek, A.; Wiese, H.; Eisenacher, M.; Meyer, H.E.; Hatt, H.H.; Oeljeklaus, S.; Warscheid, B. The membrane proteome of sensory cilia to the depth of olfactory receptors. Mol. Cell Proteom. 2014, 13, 1828–1843. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.; Zweyer, M.; Mundegar, R.R.; Swandulla, D.; Ohlendieck, K. Dataset on the comparative proteomic profiling of mouse saliva and serum from wild type versus the dystrophic mdx-4cv mouse model of dystrophinopathy. Data Brief 2018, 21, 1236–1245. [Google Scholar] [CrossRef] [PubMed]
- Mayer, U.; Ungerer, N.; Klimmeck, D.; Warnken, U.; Schnolzer, M.; Frings, S.; Mohrlen, F. Proteomic analysis of a membrane preparation from rat olfactory sensory cilia. Chem. Senses 2008, 33, 145–162. [Google Scholar] [CrossRef] [PubMed]
- Nauen, R.; Bass, C.; Feyereisen, R.; Vontas, J. The Role of Cytochrome P450s in Insect Toxicology and Resistance. Annu. Rev. Entomol. 2022, 67, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Pavlidi, N.; Vontas, J.; Van Leeuwen, T. The role of glutathione S-transferases (GSTs) in insecticide resistance in crop pests and disease vectors. Curr. Opin. Insect Sci. 2018, 27, 97–102. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Furlong, M.J. Behavior as a mechanism of insecticide resistance: Evaluation of the evidence. Curr. Opin. Insect Sci. 2017, 21, 19–25. [Google Scholar] [CrossRef]
- Fischler, W.; Kong, P.; Marella, S.; Scott, K. The detection of carbonation by the Drosophila gustatory system. Nature 2007, 448, 1054–1057. [Google Scholar] [CrossRef]
- Scott, K. Out of thin air: Sensory detection of oxygen and carbon dioxide. Neuron 2011, 69, 194–202. [Google Scholar] [CrossRef]
- Cameron, P.; Hiroi, M.; Ngai, J.; Scott, K. The molecular basis for water taste in Drosophila. Nature 2010, 465, 91–95. [Google Scholar] [CrossRef]
- Falk, R.; Bleiser-Avivi, N.; Atidia, J. Labellar taste organs of Drosophila melanogaster. J. Morphol. 1976, 150, 327–341. [Google Scholar] [CrossRef]
- Steinbrecht, R.A. Structure and function of insect olfactory sensilla. Ciba Found Symp. 1996, 200, 158–174; discussion 157–174. [Google Scholar] [CrossRef]
- Benton, R.; Sachse, S.; Michnick, S.W.; Vosshall, L.B. Atypical membrane topology and heteromeric function of Drosophila odorant receptors in vivo. PLoS Biol. 2006, 4, e20. [Google Scholar] [CrossRef] [Green Version]
- Strausfeld, N.J.; Hildebrand, J.G. Olfactory systems: Common design, uncommon origins? Curr. Opin. Neurobiol. 1999, 9, 634–639. [Google Scholar] [CrossRef]
- Clyne, P.J.; Warr, C.G.; Carlson, J.R. Candidate taste receptors in Drosophila. Science 2000, 287, 1830–1834. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Amrein, H.; Morozov, P.S.; Rzhetsky, A.; Axel, R. A spatial map of olfactory receptor expression in the Drosophila antenna. Cell 1999, 96, 725–736. [Google Scholar] [CrossRef]
- Isono, K.; Morita, H. Molecular and cellular designs of insect taste receptor system. Front. Cell Neurosci. 2010, 4, 20. [Google Scholar] [CrossRef]
- Sanchez-Gracia, A.; Vieira, F.G.; Rozas, J. Molecular evolution of the major chemosensory gene families in insects. Heredity 2009, 103, 208–216. [Google Scholar] [CrossRef]
- Shi, H.; Pei, L.; Gu, S.; Zhu, S.; Wang, Y.; Zhang, Y.; Li, B. Glutathione S-transferase (GST) genes in the red flour beetle, Tribolium castaneum, and comparative analysis with five additional insects. Genomics 2012, 100, 327–335. [Google Scholar] [CrossRef]
- Ohno, S. Evolution by Gene Duplication; Allen & Unwin; Springer: London, UK; New York, NY, USA, 1970; p. xv. 160p. [Google Scholar]
- Mitter, C.; Farrell, B.; Wiegmann, B. The phylogenetic study of adaptive zones: Has phytophagy promoted insect diversification? Am. Nat. 1988, 132, 107–128. [Google Scholar] [CrossRef]
- Saruta, F.; Yamada, N.; Yamamoto, K. Functional Analysis of an Epsilon-Class Glutathione S-Transferase From Nilaparvata lugens (Hemiptera: Delphacidae). J. Insect Sci. 2019, 19, 14. [Google Scholar] [CrossRef]
- Yang, J.; Kong, X.D.; Zhu-Salzman, K.; Qin, Q.M.; Cai, Q.N. The Key Glutathione S-Transferase Family Genes Involved in the Detoxification of Rice Gramine in Brown Planthopper Nilaparvata lugens. Insects 2021, 12, 1055. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Xu, Z.; Zou, H.; Liu, J.; Chen, S.; Feng, Q.; Zheng, S. Glutathione S-transferase SlGSTE1 in Spodoptera litura may be associated with feeding adaptation of host plants. Insect Biochem. Mol. Biol. 2016, 70, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Gloss, A.D.; Vassao, D.G.; Hailey, A.L.; Nelson Dittrich, A.C.; Schramm, K.; Reichelt, M.; Rast, T.J.; Weichsel, A.; Cravens, M.G.; Gershenzon, J.; et al. Evolution in an ancient detoxification pathway is coupled with a transition to herbivory in the drosophilidae. Mol. Biol. Evol. 2014, 31, 2441–2456. [Google Scholar] [CrossRef] [PubMed]
- Labandeira, C.C. Early history of Arthropod and vascular plant associations. Annu. Rev. Earth Planet. Sci. 1998, 26, 329–377. [Google Scholar] [CrossRef]
- Syvanen, M.; Zhou, Z.; Wharton, J.; Goldsbury, C.; Clark, A. Heterogeneity of the glutathione transferase genes encoding enzymes responsible for insecticide degradation in the housefly. J. Mol. Evol. 1996, 43, 236–240. [Google Scholar] [CrossRef]
- Wang, J.Y.; McCommas, S.; Syvanen, M. Molecular cloning of a glutathione S-transferase overproduced in an insecticide-resistant strain of the housefly (Musca domestica). Mol. Gen. Genet. MGG 1991, 227, 260–266. [Google Scholar] [CrossRef]
- Paton, M.G.; Karunaratne, S.H.; Giakoumaki, E.; Roberts, N.; Hemingway, J. Quantitative analysis of gene amplification in insecticide-resistant Culex mosquitoes. Biochem. J. 2000, 346 Pt 1, 17–24. [Google Scholar] [CrossRef]
- Yamamoto, K.; Higashiura, A.; Hirowatari, A.; Yamada, N.; Tsubota, T.; Sezutsu, H.; Nakagawa, A. Characterisation of a diazinon-metabolising glutathione S-transferase in the silkworm Bombyx mori by X-ray crystallography and genome editing analysis. Sci. Rep. 2018, 8, 16835. [Google Scholar] [CrossRef]
- Yamamoto, K.; Ichinose, H.; Aso, Y.; Banno, Y.; Kimura, M.; Nakashima, T. Molecular characterization of an insecticide-induced novel glutathione transferase in silkworm. Biochim. Biophys. Acta 2011, 1810, 420–426. [Google Scholar] [CrossRef]
- Ortelli, F.; Rossiter, L.C.; Vontas, J.; Ranson, H.; Hemingway, J. Heterologous expression of four glutathione transferase genes genetically linked to a major insecticide-resistance locus from the malaria vector Anopheles gambiae. Biochem. J. 2003, 373, 957–963. [Google Scholar] [CrossRef]
- Yu, X.; Killiny, N. RNA interference of two glutathione S-transferase genes, Diaphorina citri DcGSTe2 and DcGSTd1, increases the susceptibility of Asian citrus psyllid (Hemiptera: Liviidae) to the pesticides fenpropathrin and thiamethoxam. Pest Manag. Sci. 2018, 74, 638–647. [Google Scholar] [CrossRef]
- Bamidele, O.S.; Ajele, J.O.; Olajuyigbe, F.M.; Gaber, Y. An evaluation of glutathione transferase associated with Dichlorvos degradation in African palm weevil (Rynchophorus phoenicis) larva. Cogent Biol. 2017, 3, 1286764. [Google Scholar] [CrossRef]
- Chanut-Delalande, H.; Hashimoto, Y.; Pelissier-Monier, A.; Spokony, R.; Dib, A.; Kondo, T.; Bohere, J.; Niimi, K.; Latapie, Y.; Inagaki, S.; et al. Pri peptides are mediators of ecdysone for the temporal control of development. Nat. Cell Biol. 2014, 16, 1035–1044. [Google Scholar] [CrossRef]
- Enya, S.; Ameku, T.; Igarashi, F.; Iga, M.; Kataoka, H.; Shinoda, T.; Niwa, R. A Halloween gene noppera-bo encodes a glutathione S-transferase essential for ecdysteroid biosynthesis via regulating the behaviour of cholesterol in Drosophila. Sci. Rep. 2014, 4, 6586. [Google Scholar] [CrossRef]
- Ma, J.; Sun, L.; Zhao, H.; Wang, Z.; Zou, L.; Cao, C. Functional identification and characterization of GST genes in the Asian gypsy moth in response to poplar secondary metabolites. Pestic. Biochem. Physiol. 2021, 176, 104860. [Google Scholar] [CrossRef]
- Zhao, Y.J.; Wang, Z.Q.; Zhu, J.Y.; Liu, N.Y. Identification and characterization of detoxification genes in two cerambycid beetles, Rhaphuma horsfieldi and Xylotrechus quadripes (Coleoptera: Cerambycidae: Clytini). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2020, 243–244, 110431. [Google Scholar] [CrossRef]
- McKenna, D.D.; Scully, E.D.; Pauchet, Y.; Hoover, K.; Kirsch, R.; Geib, S.M.; Mitchell, R.F.; Waterhouse, R.M.; Ahn, S.J.; Arsala, D.; et al. Genome of the Asian longhorned beetle (Anoplophora glabripennis), a globally significant invasive species, reveals key functional and evolutionary innovations at the beetle-plant interface. Genome Biol. 2016, 17, 227. [Google Scholar] [CrossRef]
- Schoville, S.D.; Chen, Y.H.; Andersson, M.N.; Benoit, J.B.; Bhandari, A.; Bowsher, J.H.; Brevik, K.; Cappelle, K.; Chen, M.M.; Childers, A.K.; et al. A model species for agricultural pest genomics: The genome of the Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Sci. Rep. 2018, 8, 1931. [Google Scholar] [CrossRef]
- Wu, Z.; Bin, S.; He, H.; Wang, Z.; Li, M.; Lin, J. Differential Expression Analysis of Chemoreception Genes in the Striped Flea Beetle Phyllotreta striolata Using a Transcriptomic Approach. PLoS ONE 2016, 11, e0153067. [Google Scholar] [CrossRef]
- Dai, L.; Ma, J.; Ma, M.; Zhang, H.; Shi, Q.; Zhang, R.; Chen, H. Characterisation of GST genes from the Chinese white pine beetle Dendroctonus armandi (Curculionidae: Scolytinae) and their response to host chemical defence. Pest Manag. Sci. 2016, 72, 816–827. [Google Scholar] [CrossRef]
- Keeling, C.I.; Yuen, M.M.; Liao, N.Y.; Docking, T.R.; Chan, S.K.; Taylor, G.A.; Palmquist, D.L.; Jackman, S.D.; Nguyen, A.; Li, M.; et al. Draft genome of the mountain pine beetle, Dendroctonus ponderosae Hopkins, a major forest pest. Genome Biol. 2013, 14, R27. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Ye, K.; Tu, X.F.; Lu, Y.J.; Thakur, K.; Jiang, L.; Wei, Z.J. Identification and expression profiles of twenty-six glutathione S-transferase genes from rice weevil, Sitophilus oryzae (Coleoptera: Curculionidae). Int. J. Biol. Macromol. 2018, 120, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; McKenna, D.; Scully, E.; Cook, S.C.; Dainat, B.; Egekwu, N.; Grubbs, N.; Lopez, D.; Lorenzen, M.D.; Reyna, S.M.; et al. Genome of the small hive beetle (Aethina tumida, Coleoptera: Nitidulidae), a worldwide parasite of social bee colonies, provides insights into detoxification and herbivory. Gigascience 2018, 7, giy138. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.M.; Markov, G.V.; Baskaran, P.; Herrmann, M.; Sommer, R.J.; Rodelsperger, C. Draft Genome of the Scarab Beetle Oryctes borbonicus on La Reunion Island. Genome Biol. Evol. 2016, 8, 2093–2105. [Google Scholar] [CrossRef]
- Liu, S.; Rao, X.J.; Li, M.Y.; Feng, M.F.; He, M.Z.; Li, S.G. Glutathione s-transferase genes in the rice leaffolder, cnaphalocrocis medinalis (Lepidoptera: Pyralidae): Identification and expression profiles. Arch Insect Biochem. Physiol. 2015, 90, 1–13. [Google Scholar] [CrossRef]
- Nair, P.M.; Choi, J. Identification, characterization and expression profiles of Chironomus riparius glutathione S-transferase (GST) genes in response to cadmium and silver nanoparticles exposure. Aquat. Toxicol. 2011, 101, 550–560. [Google Scholar] [CrossRef]
- Strode, C.; Wondji, C.S.; David, J.P.; Hawkes, N.J.; Lumjuan, N.; Nelson, D.R.; Drane, D.R.; Karunaratne, S.H.; Hemingway, J.; Black, W.C.t.; et al. Genomic analysis of detoxification genes in the mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 2008, 38, 113–123. [Google Scholar] [CrossRef]
- Yan, L.; Yang, P.; Jiang, F.; Cui, N.; Ma, E.; Qiao, C.; Cui, F. Transcriptomic and phylogenetic analysis of Culex pipiens quinquefasciatus for three detoxification gene families. BMC Genom. 2012, 13, 609. [Google Scholar] [CrossRef]
- Niranjan Reddy, B.P.; Prasad, G.B.; Raghavendra, K. In silico characterization and comparative genomic analysis of the Culex quinquefasciatus glutathione S-transferase (GST) supergene family. Parasitol. Res. 2011, 109, 1165–1177. [Google Scholar] [CrossRef]
- Hu, F.; Dou, W.; Wang, J.J.; Jia, F.X.; Wang, J.J. Multiple glutathione S-transferase genes: Identification and expression in oriental fruit fly, Bactrocera dorsalis. Pest Manag. Sci. 2014, 70, 295–303. [Google Scholar] [CrossRef]
- Papanicolaou, A.; Schetelig, M.F.; Arensburger, P.; Atkinson, P.W.; Benoit, J.B.; Bourtzis, K.; Castanera, P.; Cavanaugh, J.P.; Chao, H.; Childers, C.; et al. The whole genome sequence of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann), reveals insights into the biology and adaptive evolution of a highly invasive pest species. Genome Biol. 2016, 17, 192. [Google Scholar] [CrossRef]
- Aidlin Harari, O.; Santos-Garcia, D.; Musseri, M.; Moshitzky, P.; Patel, M.; Visendi, P.; Seal, S.; Sertchook, R.; Malka, O.; Morin, S. Molecular Evolution of the Glutathione S-Transferase Family in the Bemisia tabaci Species Complex. Genome Biol. Evol. 2020, 12, 3857–3872. [Google Scholar] [CrossRef]
- Bailey, E.; Field, L.; Rawlings, C.; King, R.; Mohareb, F.; Pak, K.H.; Hughes, D.; Williamson, M.; Ganko, E.; Buer, B.; et al. A scaffold-level genome assembly of a minute pirate bug, Orius laevigatus (Hemiptera: Anthocoridae), and a comparative analysis of insecticide resistance-related gene families with hemipteran crop pests. BMC Genom. 2022, 23, 45. [Google Scholar] [CrossRef]
- Ramsey, J.S.; Rider, D.S.; Walsh, T.K.; De Vos, M.; Gordon, K.H.; Ponnala, L.; Macmil, S.L.; Roe, B.A.; Jander, G. Comparative analysis of detoxification enzymes in Acyrthosiphon pisum and Myzus persicae. Insect Mol. Biol. 2010, 19 (Suppl. S2), 155–164. [Google Scholar] [CrossRef]
- Zhou, W.W.; Li, X.W.; Quan, Y.H.; Cheng, J.; Zhang, C.X.; Gurr, G.; Zhu, Z.R. Identification and expression profiles of nine glutathione S-transferase genes from the important rice phloem sap-sucker and virus vector Laodelphax striatellus (Fallen) (Hemiptera: Delphacidae). Pest Manag. Sci. 2012, 68, 1296–1305. [Google Scholar] [CrossRef]
- Xue, J.; Zhou, X.; Zhang, C.X.; Yu, L.L.; Fan, H.W.; Wang, Z.; Xu, H.J.; Xi, Y.; Zhu, Z.R.; Zhou, W.W.; et al. Genomes of the rice pest brown planthopper and its endosymbionts reveal complex complementary contributions for host adaptation. Genome Biol. 2014, 15, 521. [Google Scholar] [CrossRef]
- Zhou, W.W.; Liang, Q.M.; Xu, Y.; Gurr, G.M.; Bao, Y.Y.; Zhou, X.P.; Zhang, C.X.; Cheng, J.; Zhu, Z.R. Genomic insights into the glutathione S-transferase gene family of two rice planthoppers, Nilaparvata lugens (Stal) and Sogatella furcifera (Horvath) (Hemiptera: Delphacidae). PLoS ONE 2013, 8, e56604. [Google Scholar] [CrossRef]
- Schama, R.; Pedrini, N.; Juarez, M.P.; Nelson, D.R.; Torres, A.Q.; Valle, D.; Mesquita, R.D. Rhodnius prolixus supergene families of enzymes potentially associated with insecticide resistance. Insect Biochem. Mol. Biol. 2016, 69, 91–104. [Google Scholar] [CrossRef]
- Sadd, B.M.; Barribeau, S.M.; Bloch, G.; de Graaf, D.C.; Dearden, P.; Elsik, C.G.; Gadau, J.; Grimmelikhuijzen, C.J.; Hasselmann, M.; Lozier, J.D.; et al. The genomes of two key bumblebee species with primitive eusocial organization. Genome Biol. 2015, 16, 76. [Google Scholar] [CrossRef]
- Zhang, X.R.; Zhang, J.Q.; Shao, Y.Y.; Xing, X.R.; Wang, J.; Liu, Z.X.; Li, Y.J.; Ofori, A.D.; Tu, Q.B.; Wang, J.; et al. Identification of glutathione-S-transferase genes by transcriptome analysis in Meteorus pulchricornis (Hymenoptera: Braconidae) and their expression patterns under stress of phoxim and cypermethrin. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 31, 100607. [Google Scholar] [CrossRef]
- Oakeshott, J.G.; Johnson, R.M.; Berenbaum, M.R.; Ranson, H.; Cristino, A.S.; Claudianos, C. Metabolic enzymes associated with xenobiotic and chemosensory responses in Nasonia vitripennis. Insect Mol. Biol. 2010, 19 (Suppl. S1), 147–163. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Teng, Z.W.; Gu, G.X.; Guo, L.; Wang, F.; Xiao, S.; Wang, J.L.; Wang, B.B.; Fang, Q.; Wang, F.; et al. Genomic and transcriptomic analyses of glutathione S-transferases in an endoparasitoid wasp, Pteromalus puparum. Arch. Insect Biochem. Physiol. 2020, 103, e21634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Lu, C.; Li, B.; Fang, S.; Zuo, W.; Dai, F.; Zhang, Z.; Xiang, Z. Identification, genomic organization and expression pattern of glutathione S-transferase in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1158–1164. [Google Scholar] [CrossRef]
- Cheng, J.; Wang, C.Y.; Lyu, Z.H.; Lin, T. Multiple Glutathione S-Transferase Genes in Heortia vitessoides (Lepidoptera: Crambidae): Identification and Expression Patterns. J. Insect Sci. 2018, 18, 23. [Google Scholar] [CrossRef]
- Cheng, T.; Wu, J.; Wu, Y.; Chilukuri, R.V.; Huang, L.; Yamamoto, K.; Feng, L.; Li, W.; Chen, Z.; Guo, H.; et al. Genomic adaptation to polyphagy and insecticides in a major East Asian noctuid pest. Nat. Ecol. Evol. 2017, 1, 1747–1756. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, Y.X.; Wang, W.L.; Zhang, B.X.; Li, S.G. Identification and characterisation of seventeen glutathione S-transferase genes from the cabbage white butterfly Pieris rapae. Pestic. Biochem. Physiol. 2017, 143, 102–110. [Google Scholar] [CrossRef]
- You, Y.; Xie, M.; Ren, N.; Cheng, X.; Li, J.; Ma, X.; Zou, M.; Vasseur, L.; Gurr, G.M.; You, M. Characterization and expression profiling of glutathione S-transferases in the diamondback moth, Plutella xylostella (L.). BMC Genom. 2015, 16, 152. [Google Scholar] [CrossRef]
- Huang, X.L.; Fan, D.S.; Liu, L.; Feng, J.N. Identification and characterization of glutathione S-transferase genes in the antennae of codling moth (Lepidoptera: Tortricidae). Ann. Entomol. Soc. Am. 2017, 110, 409–416. [Google Scholar] [CrossRef]
- Wang, X.; Fang, X.; Yang, P.; Jiang, X.; Jiang, F.; Zhao, D.; Li, B.; Cui, F.; Wei, J.; Ma, C.; et al. The locust genome provides insight into swarm formation and long-distance flight. Nat. Commun. 2014, 5, 2957. [Google Scholar] [CrossRef]
- Kirkness, E.F.; Haas, B.J.; Sun, W.; Braig, H.R.; Perotti, M.A.; Clark, J.M.; Lee, S.H.; Robertson, H.M.; Kennedy, R.C.; Elhaik, E.; et al. Genome sequences of the human body louse and its primary endosymbiont provide insights into the permanent parasitic lifestyle. Proc. Natl. Acad. Sci. USA 2010, 107, 12168–12173. [Google Scholar] [CrossRef]
- Wei, D.D.; Chen, E.H.; Ding, T.B.; Chen, S.C.; Dou, W.; Wang, J.J. De novo assembly, gene annotation, and marker discovery in stored-product pest Liposcelis entomophila (Enderlein) using transcriptome sequences. PLoS ONE 2013, 8, e80046. [Google Scholar] [CrossRef]
- Younus, F.; Chertemps, T.; Pearce, S.L.; Pandey, G.; Bozzolan, F.; Coppin, C.W.; Russell, R.J.; Maibeche-Coisne, M.; Oakeshott, J.G. Identification of candidate odorant degrading gene/enzyme systems in the antennal transcriptome of Drosophila melanogaster. Insect Biochem. Mol. Biol. 2014, 53, 30–43. [Google Scholar] [CrossRef] [Green Version]
- Corcoran, J.A.; Jordan, M.D.; Thrimawithana, A.H.; Crowhurst, R.N.; Newcomb, R.D. The Peripheral Olfactory Repertoire of the Lightbrown Apple Moth, Epiphyas postvittana. PLoS ONE 2015, 10, e0128596. [Google Scholar] [CrossRef]
- Liu, S.; Gong, Z.J.; Rao, X.J.; Li, M.Y.; Li, S.G. Identification of Putative Carboxylesterase and Glutathione S-transferase Genes from the Antennae of the Chilo suppressalis (Lepidoptera: Pyralidae). J. Insect Sci. 2015, 15, 103. [Google Scholar] [CrossRef]
- Tan, X.; Hu, X.M.; Zhong, X.W.; Chen, Q.M.; Xia, Q.Y.; Zhao, P. Antenna-specific glutathione S-transferase in male silkmoth Bombyx mori. Int. J. Mol. Sci. 2014, 15, 7429–7443. [Google Scholar] [CrossRef]
- Leal, W.S.; Ishida, Y.; Pelletier, J.; Xu, W.; Rayo, J.; Xu, X.; Ames, J.B. Olfactory proteins mediating chemical communication in the navel orangeworm moth, Amyelois transitella. PLoS ONE 2009, 4, e7235. [Google Scholar] [CrossRef]
- Wang, G.R.; Guo, Y.Y.; Wu, K.M. Cloning of a cDNA fragment of an antenna-specific gene in Helicoverpa armigera. Chin. J. Agric. Biotechnol. 2002, 1, 37–43. [Google Scholar] [CrossRef]
- Rogers, M.E.; Jani, M.K.; Vogt, R.G. An olfactory-specific glutathione-S-transferase in the sphinx moth Manduca sexta. J. Exp. Biol. 1999, 202, 1625–1637. [Google Scholar] [CrossRef]
- Durand, N.; Pottier, M.A.; Siaussat, D.; Bozzolan, F.; Maibeche, M.; Chertemps, T. Glutathione-S-Transferases in the Olfactory Organ of the Noctuid Moth Spodoptera littoralis, Diversity and Conservation of Chemosensory Clades. Front. Physiol. 2018, 9, 1283. [Google Scholar] [CrossRef]
- Mamidala, P.; Wijeratne, A.J.; Wijeratne, S.; Poland, T.; Qazi, S.S.; Doucet, D.; Cusson, M.; Beliveau, C.; Mittapalli, O. Identification of odor-processing genes in the emerald ash borer, Agrilus planipennis. PLoS ONE 2013, 8, e56555. [Google Scholar] [CrossRef]
- Gu, X.C.; Zhang, Y.N.; Kang, K.; Dong, S.L.; Zhang, L.W. Antennal Transcriptome Analysis of Odorant Reception Genes in the Red Turpentine Beetle (RTB), Dendroctonus valens. PLoS ONE 2015, 10, e0125159. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Xu, S.; Xie, C.; Geng, H.; Zhao, Y.; Li, J.; Huang, W.F.; Lin, Y.; Li, Z.; Su, S. Comparative transcriptome analysis of Apis mellifera antennae of workers performing different tasks. Mol. Genet Genom. 2018, 293, 237–248. [Google Scholar] [CrossRef]
- Carr, A.L.; Mitchell, R.D., III; Dhammi, A.; Bissinger, B.W.; Sonenshine, D.E.; Roe, R.M. Tick Haller’s Organ, a New Paradigm for Arthropod Olfaction: How Ticks Differ from Insects. Int. J. Mol. Sci. 2017, 18, 1563. [Google Scholar] [CrossRef] [PubMed]
- Saisawang, C.; Ketterman, A.J. Micro-plasticity of genomes as illustrated by the evolution of glutathione transferases in 12 Drosophila species. PLoS ONE 2014, 9, e109518. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, R.; Singh, S.P.; Mondal, A.K.; Benes, H.; Zimniak, P. Cloning, expression and biochemical characterization of one Epsilon-class (GST-3) and ten Delta-class (GST-1) glutathione S-transferases from Drosophila melanogaster, and identification of additional nine members of the Epsilon class. Biochem. J. 2003, 370, 661–669. [Google Scholar] [CrossRef]
- Li, G.W.; Chen, X.L.; Xu, X.L.; Wu, J.X. Degradation of sex pheromone and plant volatile components by an antennal glutathione S-transferase in the oriental fruit moth, Grapholita molesta Busck (Lepidoptera: Tortricidae). Arch Insect Biochem.Physiol. 2018, 99, e21512. [Google Scholar] [CrossRef]
- Francis, F.; Vanhaelen, N.; Haubruge, E. Glutathione S-transferases in the adaptation to plant secondary metabolites in the Myzus persicae aphid. Arch Insect Biochem. Physiol. 2005, 58, 166–174. [Google Scholar] [CrossRef]
- Dweck, H.K.; Talross, G.J.; Wang, W.; Carlson, J.R. Evolutionary shifts in taste coding in the fruit pest Drosophila suzukii. Elife 2021, 10, e64317. [Google Scholar] [CrossRef]
- Lombardo, F.; Salvemini, M.; Fiorillo, C.; Nolan, T.; Zwiebel, L.J.; Ribeiro, J.M.; Arca, B. Deciphering the olfactory repertoire of the tiger mosquito Aedes albopictus. BMC Genom. 2017, 18, 770. [Google Scholar] [CrossRef]
- Jordan, M.D.; Stanley, D.; Marshall, S.D.; De Silva, D.; Crowhurst, R.N.; Gleave, A.P.; Greenwood, D.R.; Newcomb, R.D. Expressed sequence tags and proteomics of antennae from the tortricid moth, Epiphyas postvittana. Insect Mol. Biol. 2008, 17, 361–373. [Google Scholar] [CrossRef]
- Ono, H.; Ozaki, K.; Yoshikawa, H. Identification of cytochrome P450 and glutathione-S-transferase genes preferentially expressed in chemosensory organs of the swallowtail butterfly, Papilio xuthus L. Insect Biochem. Mol. Biol. 2005, 35, 837–846. [Google Scholar] [CrossRef]
- Liu, H.; Tang, Y.; Wang, Q.; Shi, H.; Yin, J.; Li, C. Identification and Characterization of an Antennae-Specific Glutathione S-Transferase From the Indian Meal Moth. Front. Physiol. 2021, 12, 727619. [Google Scholar] [CrossRef]
- Legeai, F.; Malpel, S.; Montagne, N.; Monsempes, C.; Cousserans, F.; Merlin, C.; Francois, M.C.; Maibeche-Coisne, M.; Gavory, F.; Poulain, J.; et al. An Expressed Sequence Tag collection from the male antennae of the Noctuid moth Spodoptera littoralis: A resource for olfactory and pheromone detection research. BMC Genom. 2011, 12, 86. [Google Scholar] [CrossRef]
- Agerbirk, N.; Olsen, C.E. Glucosinolate structures in evolution. Phytochemistry 2012, 77, 16–45. [Google Scholar] [CrossRef]
- Kolm, R.H.; Danielson, U.H.; Zhang, Y.; Talalay, P.; Mannervik, B. Isothiocyanates as substrates for human glutathione transferases: Structure-activity studies. Biochem. J. 1995, 311 Pt 2, 453–459. [Google Scholar] [CrossRef]
- Harzsch, S.; Krieger, J. Crustacean olfactory systems: A comparative review and a crustacean perspective on olfaction in insects. Prog. Neurobiol. 2018, 161, 23–60. [Google Scholar] [CrossRef]
- Graziadei, P.P.; Graziadei, G.A. Neurogenesis and neuron regeneration in the olfactory system of mammals. I. Morphological aspects of differentiation and structural organization of the olfactory sensory neurons. J. Neurocytol. 1979, 8, 1–18. [Google Scholar] [CrossRef]
- Neiers, F.; Gourrat, K.; Canon, F.; Schwartz, M. Metabolism of Cysteine Conjugates and Production of Flavor Sulfur Compounds by a Carbon-Sulfur Lyase from the Oral Anaerobe Fusobacterium nucleatum. J. Agric. Food Chem. 2022, 70, 9969–9979. [Google Scholar] [CrossRef]
- Schwartz, M.; Canon, F.; Feron, G.; Neiers, F.; Gamero, A. Impact of Oral Microbiota on Flavor Perception: From Food Processing to In-Mouth Metabolization. Foods 2021, 10, 2006. [Google Scholar] [CrossRef]
- Ai, S.; Zhang, Y.; Chen, Y.; Zhang, T.; Zhong, G.; Yi, X. Insect-Microorganism Interaction Has Implicates on Insect Olfactory Systems. Insects 2022, 13, 1084. [Google Scholar] [CrossRef]

| Order | Insect Species | Cytosolic | Total | Ref. | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Delta | Epsilon | Omega | Sigma | Theta | Zeta | Unclassified | ||||
| Coleoptera | Lasioderma serricorne | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 3 | [98] |
| Agrilus planipennis | 5 | 9 | 0 | 2 | 0 | 0 | 0 | 16 | [98] | |
| Anoplophora glabripennis | 10 | 10 | 2 | 4 | 2 | 0 | 0 | 28 | [99] | |
| Rhaphuma horsfieldi | 5 | 8 | 3 | 2 | 1 | 1 | 0 | 20 | [98] | |
| Xylotrechus quadripes | 5 | 7 | 2 | 2 | 1 | 1 | 0 | 18 | [98] | |
| Diabrotica virgifera | 3 | 11 | 1 | 0 | 2 | 0 | 0 | 17 | [98] | |
| Leptinotarsa Decemlineata | 6 | 11 | 7 | 6 | 2 | 1 | 0 | 33 | [100] | |
| Phyllotreta striolata | 5 | 6 | 2 | 6 | 1 | 1 | 2 | 23 | [101] | |
| Dendroctonus armandi | 0 | 4 | 1 | 2 | 1 | 0 | 0 | 8 | [102] | |
| Dendroctonus ponderosae | 6 | 12 | 2 | 5 | 2 | 1 | 0 | 28 | [103] | |
| Lissorhoptrus oryzophilus | 3 | 7 | 2 | 8 | 1 | 1 | 2 | 24 | [98] | |
| Sitophilus oryzae | 2 | 12 | 3 | 6 | 2 | 1 | 0 | 26 | [104] | |
| Aethina tumida | 3 | 19 | 1 | 7 | 1 | 5 | 7 | 43 | [105] | |
| Oryctes borbonicus | 4 | 5 | 3 | 15 | 3 | 1 | 0 | 31 | [106] | |
| Onthophagus taurus | 4 | 7 | 3 | 1 | 4 | 0 | 0 | 19 | [98] | |
| Nicrophorus vespilloides | 8 | 6 | 0 | 1 | 3 | 0 | 0 | 18 | [98] | |
| Asbolus verrucosus | 3 | 14 | 2 | 2 | 1 | 0 | 0 | 22 | [98] | |
| Tribolium castaneum | 3 | 19 | 3 | 7 | 1 | 1 | 2 | 36 | [79] | |
| Tenebrio molitor | 2 | 13 | 1 | 5 | 1 | 1 | 2 | 25 | [107] | |
| Diptera | Chironomus riparius | 3 | 1 | 1 | 4 | 1 | 1 | 2 | 13 | [108] |
| Aedes aegypti | 8 | 8 | 1 | 1 | 4 | 1 | 3 | 26 | [109] | |
| Anopheles gambiae | 17 | 8 | 1 | 1 | 2 | 1 | 2 | 32 | [79] | |
| Culex quinquefasciatus | 14 | 10 | 1 | 1 | 6 | 0 | 3 | 35 | [110,111] | |
| Drosophila melanogaster | 11 | 14 | 4 | 1 | 4 | 2 | 1 | 37 | [79] | |
| Bactrocera dorsalis | 9 | 5 | 3 | 1 | 3 | 3 | 1 | 25 | [112] | |
| Ceratitis capitata | 7 | 14 | 1 | 1 | 3 | 2 | 1 | 29 | [113] | |
| Hemiptera | Bemisia tabaci | 14 | 0 | 1 | 6 | 0 | 2 | 0 | 23 | [114] |
| Orius laevigatus | 1 | 0 | 2 | 16 | 1 | 1 | 0 | 21 | [115] | |
| Acyrthosiphon pisum | 16 | 1 | 2 | 6 | 2 | 0 | 3 | 30 | [79] | |
| Myzus persicae | 8 | 0 | 0 | 8 | 2 | 0 | 0 | 18 | [116] | |
| Laodelphax striatellus | 1 | 1 | 1 | 3 | 1 | 1 | 0 | 8 | [117] | |
| Nilaparvata lugens | 2 | 1 | 1 | 3 | 1 | 1 | 0 | 9 | [118,119] | |
| Supraphorura furcifera | 2 | 1 | 1 | 1 | 1 | 1 | 0 | 7 | [119] | |
| Rhodnius prolixus | 1 | 0 | 1 | 7 | 4 | 1 | 0 | 14 | [120] | |
| Diaphorina citri | 2 | 2 | 0 | 3 | 0 | 0 | 1 | 8 | [114] | |
| Hymeno-ptera | Apis mellifera | 2 | 0 | 2 | 4 | 1 | 1 | 1 | 11 | [79] |
| Bombus impatiens | 5 | 0 | 2 | 4 | 1 | 1 | 0 | 13 | [121] | |
| Bombus terrestris | 5 | 0 | 2 | 4 | 1 | 1 | 0 | 13 | [121] | |
| Meteorus pulchricornis | 4 | 0 | 3 | 7 | 0 | 1 | 0 | 15 | [122] | |
| Nasonia vitripennis | 5 | 0 | 2 | 8 | 3 | 1 | 0 | 19 | [123] | |
| Pteromalus puparum | 5 | 0 | 2 | 8 | 3 | 1 | 0 | 19 | [124] | |
| Lepidoptera | Bombyx mori | 4 | 8 | 4 | 2 | 1 | 2 | 2 | 23 | [125] |
| Cnaphalocrocis medinalis | 4 | 9 | 3 | 5 | 0 | 2 | 2 | 25 | [107] | |
| Heortia vitessoides | 3 | 2 | 3 | 3 | 1 | 2 | 2 | 16 | [126] | |
| Spodoptera litura | 5 | 21 | 3 | 7 | 1 | 2 | 3 | 42 | [127] | |
| Danaus plexippus | 3 | 6 | 3 | 5 | 1 | 3 | 2 | 23 | [98] | |
| Pieris rapae | 3 | 3 | 4 | 4 | 1 | 2 | 0 | 17 | [128] | |
| Plutella xylostella | 5 | 5 | 5 | 2 | 1 | 2 | 2 | 22 | [129] | |
| Manduca sexta | 6 | 9 | 4 | 2 | 1 | 2 | 1 | 25 | [98] | |
| Cydia pomonella | 4 | 3 | 2 | 1 | 1 | 1 | 1 | 13 | [130] | |
| Orthoptera | Locusta migratoria | 10 | 0 | 3 | 12 | 2 | 1 | 0 | 28 | [131] |
| Phthir-aptera | Pediculus humanus | 4 | 0 | 1 | 4 | 1 | 1 | 0 | 11 | [132] |
| Psocoptera | Liposcelis entomophila | 17 | 0 | 1 | 13 | 3 | 1 | 0 | 35 | [133] |
| Order | Insect Species | Location | GST Classes | Ref. |
|---|---|---|---|---|
| Coleoptera | Agrilus planipennis | Antennae | Delta | [142] |
| Dendroctonus valens | Antennae | Not indicated | [143] | |
| Phyllotreta striolata | Antennae | Delta and Epsilon | [101] | |
| Diptera | Aedes albopictus | Antennae/maxillary palps | Not indicated | [151] |
| Drosophila melanogaster | Antennae/maxillary palps/labellum | Delta, Epsilon, Omega, Sigma, Theta, and Zeta | [5,17,134] | |
| Hymenoptera | Apis melifera | Antennae | Delta | [144] |
| Ixodida | Dermacentor variabilis | Haller’s organ | Epsilon and Mu | [145] |
| Lepidoptera | Bombyx Mori | Antennae | Delta | [137] |
| Chilo suppressalis | Antennae | Delta, Epsilon, Omega, Sigma, Theta, and Zeta | [136] | |
| Cydia pomonella | Antennae | Delta, Epsilon, Omega, Sigma, Theta, and Zeta | [130] | |
| Epiphyas postvittana | Antennae | Delta, Epsilon, Omega, Sigma, and Theta | [152] | |
| Grapholita molesta | Antennae | Delta | [148] | |
| Helicoverpa armigera | Antenna | Delta | [139] | |
| Heortia vitessoides | Antennae | Delta and Epsilon | [126] | |
| Manduca sexta | Antennae | Delta | [140] | |
| Papilio xuthus | Antennae, labella, and tarsi | Delta | [153] | |
| Plodia interpunctella | Antennae | Delta, Epsilon, Omega, Sigma, Theta, and Zeta | [154] | |
| Spodoptera littoralis | Antennae | Delta, Epsilon, Omega, Sigma, Theta, and Zeta | [141,155] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwartz, M.; Boichot, V.; Fraichard, S.; Muradova, M.; Senet, P.; Nicolai, A.; Lirussi, F.; Bas, M.; Canon, F.; Heydel, J.-M.; et al. Role of Insect and Mammal Glutathione Transferases in Chemoperception. Biomolecules 2023, 13, 322. https://doi.org/10.3390/biom13020322
Schwartz M, Boichot V, Fraichard S, Muradova M, Senet P, Nicolai A, Lirussi F, Bas M, Canon F, Heydel J-M, et al. Role of Insect and Mammal Glutathione Transferases in Chemoperception. Biomolecules. 2023; 13(2):322. https://doi.org/10.3390/biom13020322
Chicago/Turabian StyleSchwartz, Mathieu, Valentin Boichot, Stéphane Fraichard, Mariam Muradova, Patrick Senet, Adrien Nicolai, Frederic Lirussi, Mathilde Bas, Francis Canon, Jean-Marie Heydel, and et al. 2023. "Role of Insect and Mammal Glutathione Transferases in Chemoperception" Biomolecules 13, no. 2: 322. https://doi.org/10.3390/biom13020322
APA StyleSchwartz, M., Boichot, V., Fraichard, S., Muradova, M., Senet, P., Nicolai, A., Lirussi, F., Bas, M., Canon, F., Heydel, J. -M., & Neiers, F. (2023). Role of Insect and Mammal Glutathione Transferases in Chemoperception. Biomolecules, 13(2), 322. https://doi.org/10.3390/biom13020322