
Peptide-Based siRNA Nanocomplexes Targeting Hepatic Stellate Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Synthesis of the Dimeric IGF2R Peptide Ligands
2.3. Fabrication and Characterization of the Nanocomplex
2.4. siRNA Entrapment and Serum Stability of the Nanocomplex
2.5. Cell Culture
2.6. Cytotoxicity of the Nanocomplex
2.7. Cellular Uptake of the Nanocomplex
2.8. Silencing Activity of the Nanocomplex
2.9. In Vivo Biodistribution Study
2.10. Statistical Analysis
3. Results
3.1. Fabrication and Characterization of the Nanocomplex
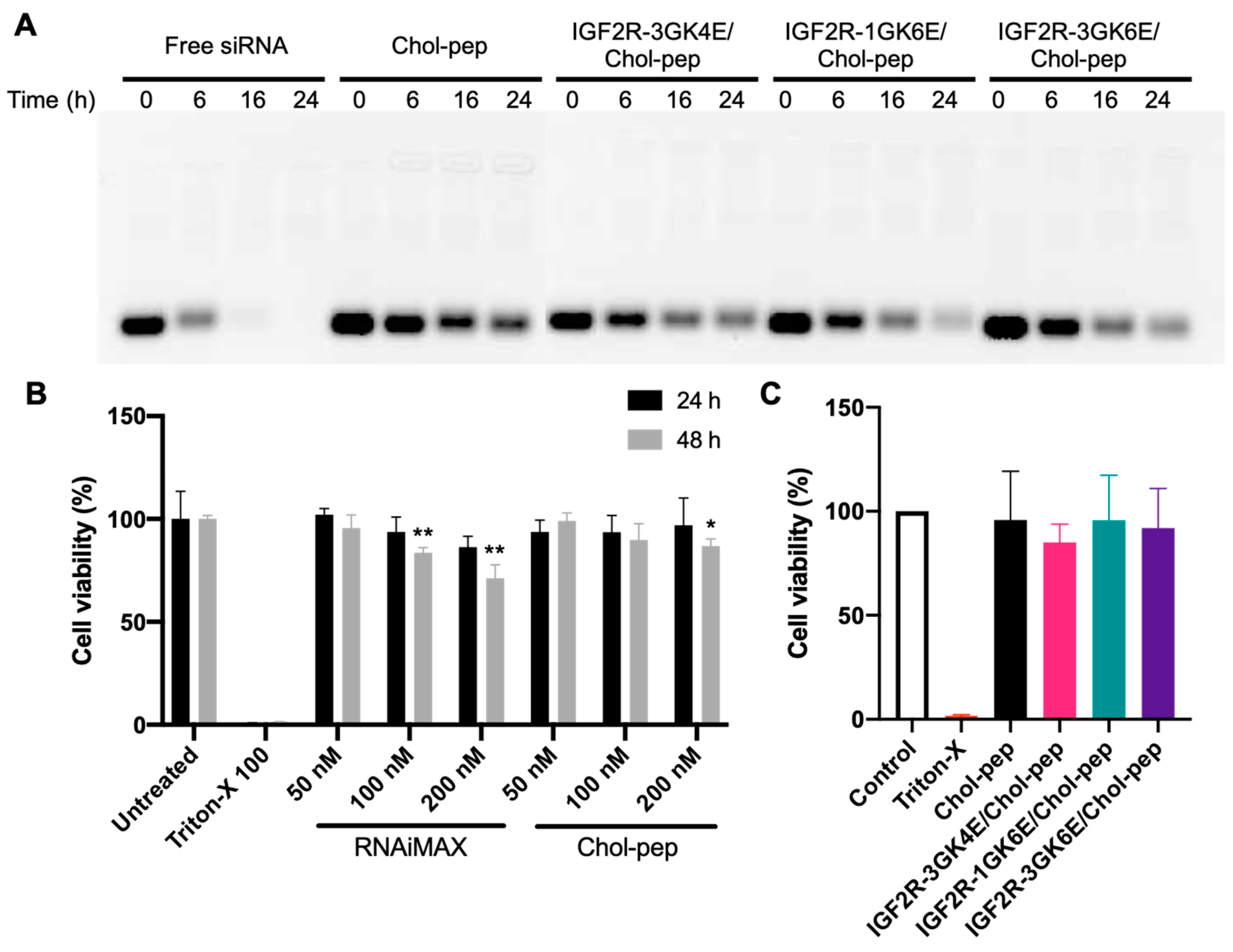
3.2. Serum Stability and Cytotoxicity of the Nanocomplex
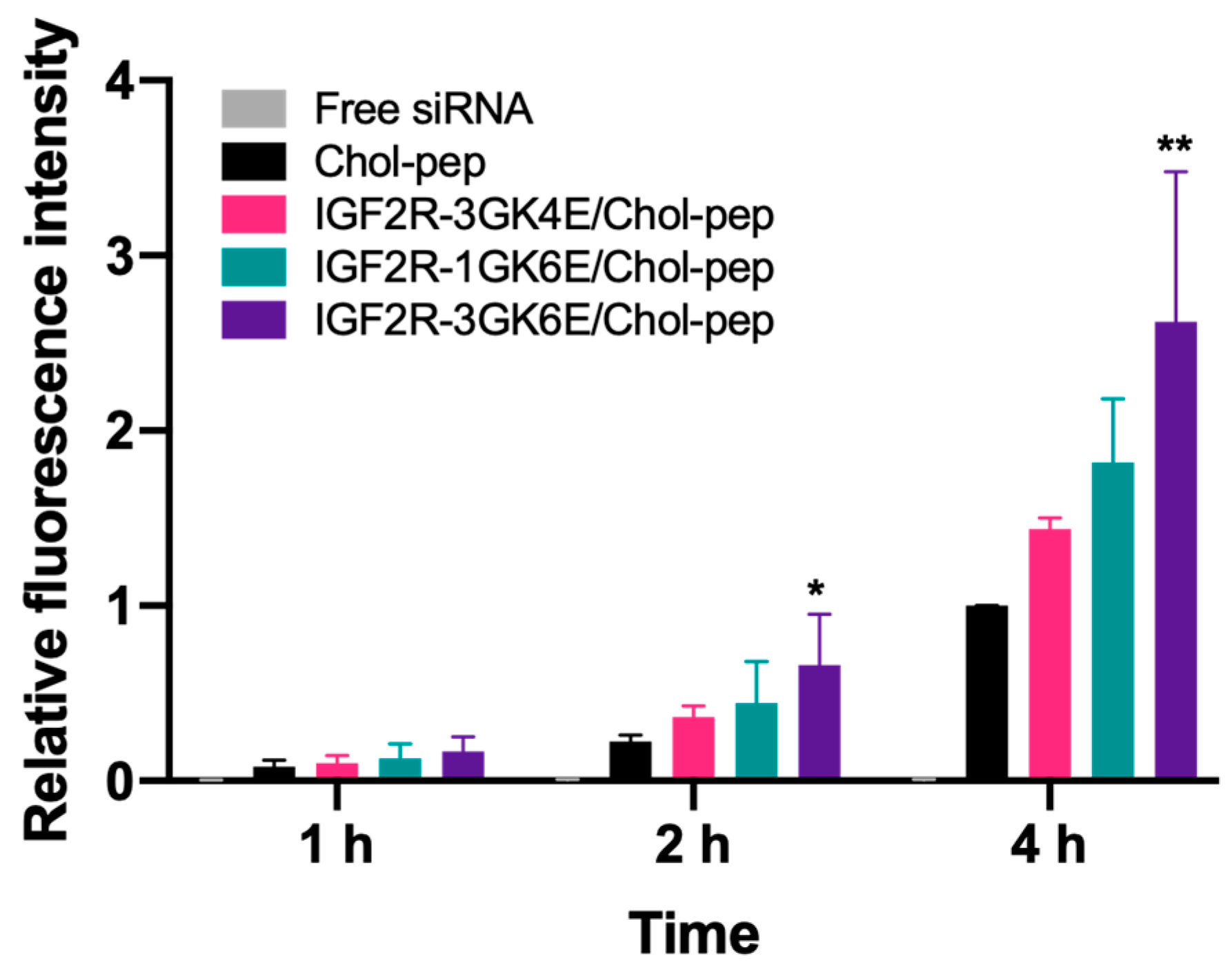
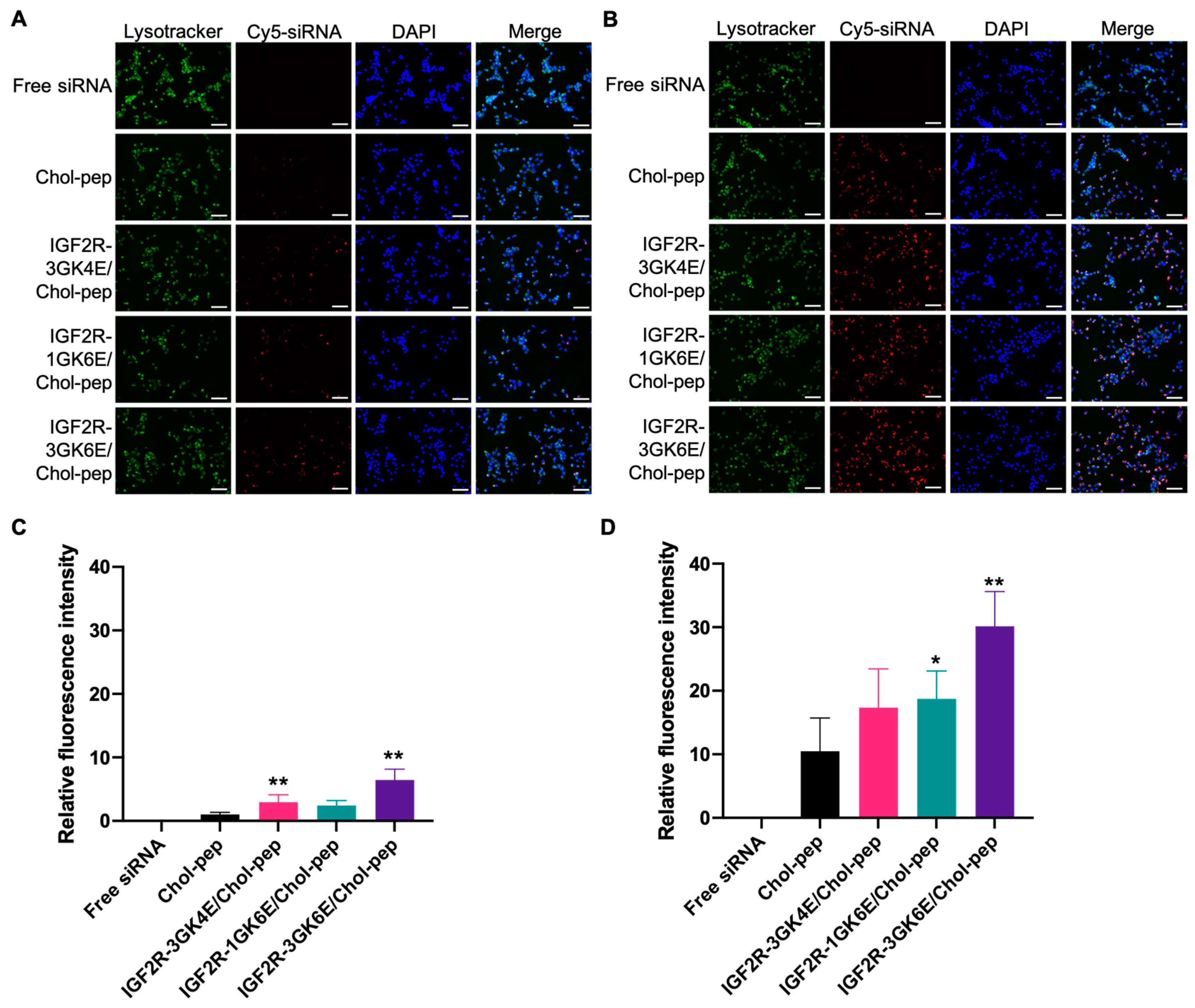
3.3. Cellular Uptake of the Nanocomplex
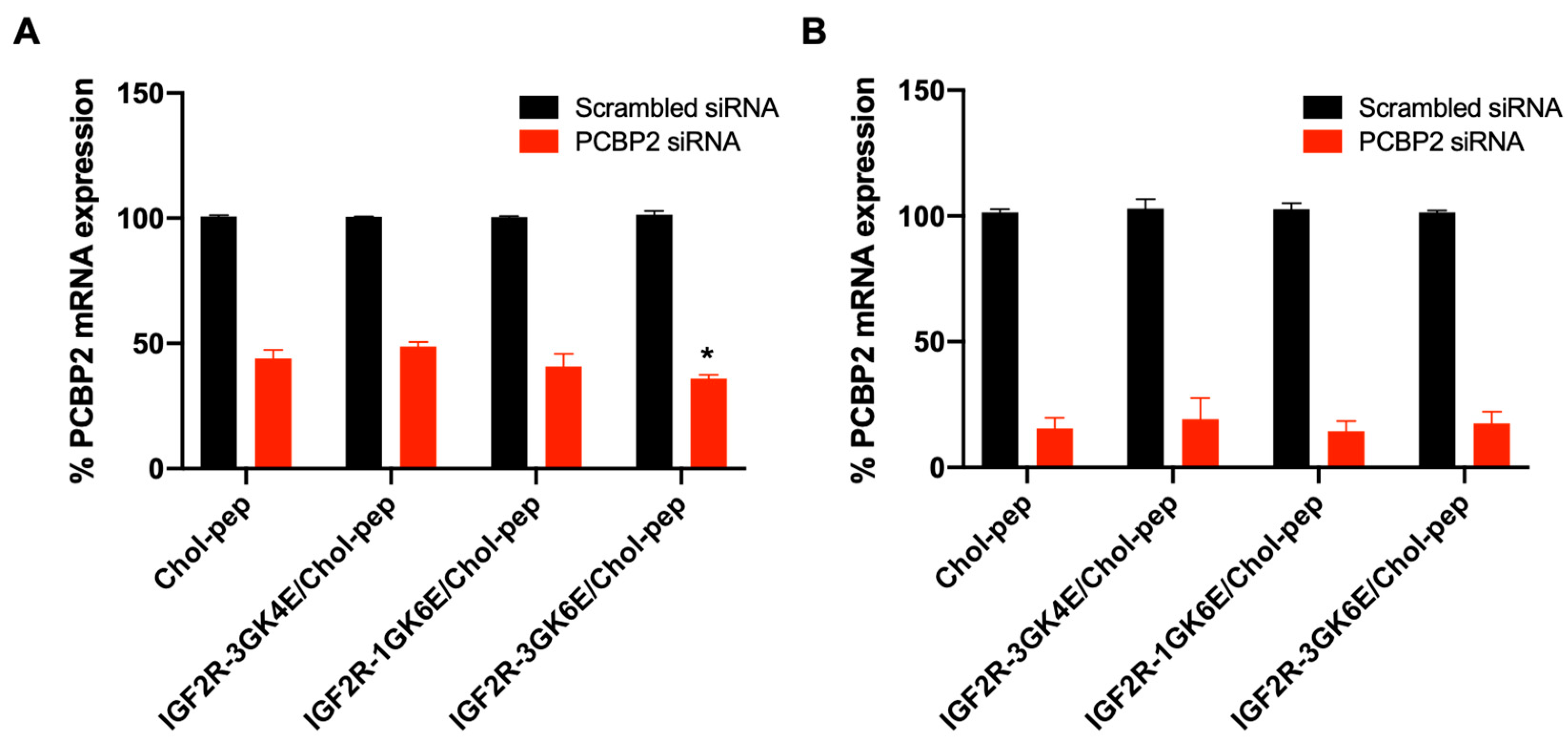
3.4. Silencing Activity of the Nanocomplex
3.5. In Vivo Biodistribution Study
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.J.; Wu, H.H.; Wang, Y.J. Strategies to prevent and reverse liver fibrosis in humans and laboratory animals. Arch. Toxicol. 2015, 89, 1727–1750. [Google Scholar] [CrossRef] [PubMed]
- Weiskirchen, R.; Weiskirchen, S.; Tacke, F. Recent advances in understanding liver fibrosis: Bridging basic science and individualized treatment concepts. F1000Research 2018, 7, F1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iredale, J.P. Models of liver fibrosis: Exploring the dynamic nature of inflammation and repair in a solid organ. J. Clin. Investig. 2007, 117, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Tsukada, S.; Parsons, C.J.; Rippe, R.A. Mechanisms of liver fibrosis. Clin. Chim. Acta 2006, 364, 33–60. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zern, M.A. Hepatic stellate cells: A target for the treatment of liver fibrosis. J. Gastroenterol. 2000, 35, 665–672. [Google Scholar] [CrossRef]
- Lindquist, J.N.; Parsons, C.J.; Stefanovic, B.; Brenner, D.A. Regulation of alpha1(I) collagen messenger RNA decay by interactions with alphaCP at the 3′-untranslated region. J. Biol. Chem. 2004, 279, 23822–23829. [Google Scholar] [CrossRef] [Green Version]
- Shukla, R.S.; Qin, B.; Wan, Y.J.; Cheng, K. PCBP2 siRNA reverses the alcohol-induced pro-fibrogenic effects in hepatic stellate cells. Pharm. Res. 2011, 28, 3058–3068. [Google Scholar] [CrossRef] [Green Version]
- Setten, R.L.; Rossi, J.J.; Han, S.P. The current state and future directions of RNAi-based therapeutics. Nat. Rev. Drug Discov. 2019, 18, 421–446. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Z.; Jin, W.; Barve, A.; Wan, Y.Y.; Cheng, K. Silencing of alpha-complex protein-2 reverses alcohol- and cytokine-induced fibrogenesis in hepatic stellate cells. Liver Res. 2017, 1, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Tatiparti, K.; Sau, S.; Kashaw, S.K.; Iyer, A.K. siRNA Delivery Strategies: A Comprehensive Review of Recent Developments. Nanomaterials 2017, 7, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Li, Y.; Jain, A.; Chen, Z.; Liu, H.; Jin, W.; Cheng, K. Development of a peptide-modified siRNA nanocomplex for hepatic stellate cells. Nanomedicine 2018, 14, 51–61. [Google Scholar] [CrossRef]
- Jain, A.; Barve, A.; Zhao, Z.; Fetse, J.P.; Liu, H.; Li, Y.; Cheng, K. Targeted Delivery of an siRNA/PNA Hybrid Nanocomplex Reverses Carbon Tetrachloride-Induced Liver Fibrosis. Adv. Ther. 2019, 2, 1900046. [Google Scholar] [CrossRef] [PubMed]
- Sauna, Z. Immunogenicity of Protein-Based Therapeutics. Available online: https://www.fda.gov/vaccines-blood-biologics/biologics-research-projects/immunogenicity-protein-based-therapeutics (accessed on 21 February 2023).
- Qin, B.; Chen, Z.; Jin, W.; Cheng, K. Development of cholesteryl peptide micelles for siRNA delivery. J. Control. Release 2013, 172, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhao, Z.; Lin, C.Y.; Liu, Y.; Staveley, O.K.F.; Li, G.; Cheng, K. Silencing PCBP2 normalizes desmoplastic stroma and improves the antitumor activity of chemotherapy in pancreatic cancer. Theranostics 2021, 11, 2182–2200. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, Y.; Liu, H.; Jain, A.; Patel, P.V.; Cheng, K. Co-delivery of IKBKE siRNA and cabazitaxel by hybrid nanocomplex inhibits invasiveness and growth of triple-negative breast cancer. Sci. Adv. 2020, 6, eabb0616. [Google Scholar] [CrossRef]
- Chen, Z.; Jain, A.; Liu, H.; Zhao, Z.; Cheng, K. Targeted Drug Delivery to Hepatic Stellate Cells for the Treatment of Liver Fibrosis. J. Pharmacol. Exp. Ther. 2019, 370, 695–702. [Google Scholar] [CrossRef] [Green Version]
- Puxbaum, V.; Nimmerfall, E.; Bauerl, C.; Taub, N.; Blaas, P.M.; Wieser, J.; Mikula, M.; Mikulits, W.; Ng, K.M.; Yeoh, G.C.; et al. M6P/IGF2R modulates the invasiveness of liver cells via its capacity to bind mannose 6-phosphate residues. J. Hepatol. 2012, 57, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Jin, W.; Liu, H.; Zhao, Z.; Cheng, K. Discovery of Peptide ligands for hepatic stellate cells using phage display. Mol. Pharm. 2015, 12, 2180–2188. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.; Yang, N.; Mahato, R.I. TGF-beta1 gene silencing for treating liver fibrosis. Mol. Pharm. 2009, 6, 772–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Aramundiz, J.V.; Lozano, M.V.; Sousa-Herves, A.; Fernandez-Megia, E.; Csaba, N. Polypeptides and polyaminoacids in drug delivery. Expert Opin. Drug Deliv. 2012, 9, 183–201. [Google Scholar] [CrossRef] [PubMed]
- Mainini, F.; Eccles, M.R. Lipid and Polymer-Based Nanoparticle siRNA Delivery Systems for Cancer Therapy. Molecules 2020, 25, 2692. [Google Scholar] [CrossRef]
- Harris, T.J.; Green, J.J.; Fung, P.W.; Langer, R.; Anderson, D.G.; Bhatia, S.N. Tissue-specific gene delivery via nanoparticle coating. Biomaterials 2010, 31, 998–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farkhani, S.M.; Shirani, A.; Mohammadi, S.; Zakeri-Milani, P.; Shahbazi Mojarrad, J.; Valizadeh, H. Effect of poly-glutamate on uptake efficiency and cytotoxicity of cell penetrating peptides. IET Nanobiotechnol. 2016, 10, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, T.A.; Olson, E.S.; Timmers, M.M.; Jiang, T.; Tsien, R.Y. Systemic in vivo distribution of activatable cell penetrating peptides is superior to that of cell penetrating peptides. Integr. Biol. 2009, 1, 371–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadi, S.; Zakeri-Milani, P.; Golkar, N.; Farkhani, S.M.; Shirani, A.; Shahbazi Mojarrad, J.; Nokhodchi, A.; Valizadeh, H. Synthesis and cellular characterization of various nano-assemblies of cell penetrating peptide-epirubicin-polyglutamate conjugates for the enhancement of antitumor activity. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tian, B.; Li, Y.; Lei, T.; Meng, J.; Yang, L.; Zhang, Y.; Chen, F.; Zhang, H.; Xu, H.; et al. A Copper-Mediated Disulfiram-Loaded pH-Triggered PEG-Shedding TAT Peptide-Modified Lipid Nanocapsules for Use in Tumor Therapy. ACS Appl. Mater. Interfaces 2015, 7, 25147–25161. [Google Scholar] [CrossRef]
- Kaufner, L.; Cartier, R.; Wüstneck, R.; Fichtner, I.; Pietschmann, S.; Bruhn, H.; Schütt, D.; Thünemann, A.F.; Pison, U. Poly (ethylene oxide)-block-poly (glutamic acid) coated maghemite nanoparticles: In vitro characterization and in vivo behaviour. Nanotechnology 2007, 18, 115710. [Google Scholar] [CrossRef]
- Chen, R.; Wang, J.; Qian, C.; Ji, Y.; Zhu, C.; Wu, L.; Li, W.; Bi, X.; Wang, Y.; Cao, G. From Nanofibers to Nanorods: Nanostructure of Peptide-Drug Conjugates Regulated by Polypeptide-PEG Derivative and Enhanced Antitumor Effect. Adv. Funct. Mater. 2019, 29, 1806058. [Google Scholar] [CrossRef]
- Craig, J.A.; Rexeisen, E.L.; Mardilovich, A.; Shroff, K.; Kokkoli, E. Effect of linker and spacer on the design of a fibronectin-mimetic peptide evaluated via cell studies and AFM adhesion forces. Langmuir 2008, 24, 10282–10292. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.A.; Fonhus, M.S.; Kindberg, G.M.; Tolleshaug, H.; Berg, T. Enhanced activity of lysosomal proteases in activated rat hepatic stellate cells is associated with a concomitant increase in the number of the mannose-6-phosphate/insulin-like growth factor II receptor. Cell Biol. Int. 2013, 37, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Aryal, S.; Park, H.; Leary, J.F.; Key, J. Top-down fabrication-based nano/microparticles for molecular imaging and drug delivery. Int. J. Nanomed. 2019, 14, 6631–6644. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.-Y.; Mamani, U.-F.; Guo, Y.; Liu, Y.; Cheng, K. Peptide-Based siRNA Nanocomplexes Targeting Hepatic Stellate Cells. Biomolecules 2023, 13, 448. https://doi.org/10.3390/biom13030448
Lin C-Y, Mamani U-F, Guo Y, Liu Y, Cheng K. Peptide-Based siRNA Nanocomplexes Targeting Hepatic Stellate Cells. Biomolecules. 2023; 13(3):448. https://doi.org/10.3390/biom13030448
Chicago/Turabian StyleLin, Chien-Yu, Umar-Farouk Mamani, Yuhan Guo, Yanli Liu, and Kun Cheng. 2023. "Peptide-Based siRNA Nanocomplexes Targeting Hepatic Stellate Cells" Biomolecules 13, no. 3: 448. https://doi.org/10.3390/biom13030448
APA StyleLin, C. -Y., Mamani, U. -F., Guo, Y., Liu, Y., & Cheng, K. (2023). Peptide-Based siRNA Nanocomplexes Targeting Hepatic Stellate Cells. Biomolecules, 13(3), 448. https://doi.org/10.3390/biom13030448