


Actin Bundles Dynamics and Architecture


Abstract
:1. Introduction
2. Actin Organization in the Cell
3. Actin-Bundling Proteins
3.1. α-Actinin
3.2. Fimbrin/Plastin
3.3. Fascin
3.4. Espin
4. Assembly and Disassembly of Actin Bundles
4.1. Initiation
4.2. Elongation
4.3. Crosslinking
4.4. Disassembly/Severing
4.4.1. ADF/Cofilin Protein Family
4.4.2. Capping Proteins
4.4.3. Profilin
4.4.4. MICALs
4.4.5. Myosins
5. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pollard, T.D. Actin and Actin-Binding Proteins. Cold Spring Harb. Perspect. Biol. 2016, 8, a018226. [Google Scholar] [CrossRef] [Green Version]
- Lodish, H.; Berk, A.; Kaiser, C.A.; Kaiser, C.; Krieger, M.; Scott, M.P.; Bretscher, A.; Ploegh, H.; Matsudaira, P. Molecular Cell Biology; Macmillan: New York, NY, USA, 2008. [Google Scholar]
- Pollard, T.D.; Cooper, J.A. Actin, a central player in cell shape and movement. Science 2009, 326, 1208–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, C.M.; Ambaru, B.; Bajaj, R. Emerging Functions of Actins and Actin Binding Proteins in Trypanosomatids. Front. Cell Dev. Biol. 2020, 8, 587685. [Google Scholar] [CrossRef]
- Lappalainen, P. Actin-binding proteins: The long road to understanding the dynamic landscape of cellular actin networks. Mol. Biol. Cell 2016, 27, 2519–2522. [Google Scholar] [CrossRef]
- Pollard, T.D.; Blanchoin, L.; Mullins, R.D. Molecular mechanisms controlling actin filament dynamics in nonmuscle cells. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 545–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhang, Y.; Li, L.; Cao, J.; Guo, Y.; Wu, Y.; Gao, W. Fascin actin-bundling protein 1 in human cancer: Promising biomarker or therapeutic target? Mol. Ther. Oncolytics 2021, 20, 240–264. [Google Scholar] [CrossRef]
- Cheng, Y.W.; Zeng, F.M.; Li, D.J.; Wang, S.H.; He, J.Z.; Guo, Z.C.; Nie, P.J.; Wu, Z.Y.; Shi, W.Q.; Wen, B.; et al. P300/CBP-associated factor (PCAF)-mediated acetylation of Fascin at lysine 471 inhibits its actin-bundling activity and tumor metastasis in esophageal cancer. Cancer Commun. 2021, 41, 1398–1416. [Google Scholar] [CrossRef] [PubMed]
- Morley, S.C. The actin-bundling protein L-plastin supports T-cell motility and activation. Immunol. Rev. 2013, 256, 48–62. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, R.P.; Veltman, D.; Machesky, L.M. Actin-bundling proteins in cancer progression at a glance. J. Cell Sci. 2012, 125, 1073–1079. [Google Scholar] [CrossRef] [Green Version]
- Wolff, L.; Strathmann, E.A.; Müller, I.; Mählich, D.; Veltman, C.; Niehoff, A.; Wirth, B. Plastin 3 in health and disease: A matter of balance. Cell. Mol. Life Sci. 2021, 78, 5275–5301. [Google Scholar] [CrossRef] [PubMed]
- Livne, A.; Geiger, B. The inner workings of stress fibers—From contractile machinery to focal adhesions and back. J. Cell Sci. 2016, 129, 1293–1304. [Google Scholar] [CrossRef] [Green Version]
- Svitkina, T.M.; Bulanova, E.A.; Chaga, O.Y.; Vignjevic, D.M.; Kojima, S.; Vasiliev, J.M.; Borisy, G.G. Mechanism of filopodia initiation by reorganization of a dendritic network. J. Cell Biol. 2003, 160, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Xue, J.; Peterson, E.H. Architecture of the mouse utricle: Macular organization and hair bundle heights. J. Neurophysiol. 2008, 99, 718–733. [Google Scholar] [CrossRef] [Green Version]
- Tilney, L.G.; Saunders, J.C. Actin filaments, stereocilia, and hair cells of the bird cochlea. I. Length, number, width, and distribution of stereocilia of each hair cell are related to the position of the hair cell on the cochlea. J. Cell Biol. 1983, 96, 807–821. [Google Scholar] [CrossRef]
- Svitkina, T.M.; Verkhovsky, A.B.; McQuade, K.M.; Borisy, G.G. Analysis of the actin-myosin II system in fish epidermal keratocytes: Mechanism of cell body translocation. J. Cell Biol. 1997, 139, 397–415. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C. Bundling actin filaments from membranes: Some novel players. Front. Plant Sci. 2012, 3, 188. [Google Scholar] [CrossRef] [Green Version]
- Revenu, C.; Athman, R.; Robine, S.; Louvard, D. The co-workers of actin filaments: From cell structures to signals. Nat. Rev. Mol. Cell Biol. 2004, 5, 635–646. [Google Scholar] [CrossRef]
- Bartles, J.R. Parallel actin bundles and their multiple actin-bundling proteins. Curr. Opin. Cell Biol. 2000, 12, 72–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falzone, T.T.; Lenz, M.; Kovar, D.R.; Gardel, M.L. Assembly kinetics determine the architecture of alpha-actinin crosslinked F-actin networks. Nat. Commun. 2012, 3, 861. [Google Scholar] [CrossRef] [Green Version]
- Sherer, L.A.; Courtemanche, N. Cooperative bundling by fascin generates actin structures with architectures that depend on filament length. Front. Cell Dev. Biol. 2022, 10, 974047. [Google Scholar] [CrossRef] [PubMed]
- DeRosier, D.J.; Tilney, L.G. F-actin bundles are derivatives of microvilli: What does this tell us about how bundles might form? J. Cell Biol. 2000, 148, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Orbach, R.; Su, X. Surfing on Membrane Waves: Microvilli, Curved Membranes, and Immune Signaling. Front. Immunol. 2020, 11, 2187. [Google Scholar] [CrossRef] [PubMed]
- Tilney, L.G.; Derosier, D.J.; Mulroy, M.J. The organization of actin filaments in the stereocilia of cochlear hair cells. J. Cell Biol. 1980, 86, 244–259. [Google Scholar] [CrossRef] [Green Version]
- Tilney, L.G.; Tilney, M.S.; Guild, G.M. F actin bundles in Drosophila bristles. I. Two filament cross-links are involved in bundling. J. Cell Biol. 1995, 130, 629–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartles, J.R.; Wierda, A.; Zheng, L. Identification and characterization of espin, an actin-binding protein localized to the F-actin-rich junctional plaques of Sertoli cell ectoplasmic specializations. J. Cell Sci. 1996, 109, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Svitkina, T. The Actin Cytoskeleton and Actin-Based Motility. Cold Spring Harb. Perspect. Biol. 2018, 10, a018267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, M.; Doss, B.; Lim, C.T.; Voituriez, R.; Ladoux, B. Single cell rigidity sensing: A complex relationship between focal adhesion dynamics and large-scale actin cytoskeleton remodeling. Cell Adhes. Migr. 2016, 10, 554–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogilner, A.; Rubinstein, B. The physics of filopodial protrusion. Biophys. J. 2005, 89, 782–795. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.B.; Krey, J.F.; Hassan, A.; Metlagel, Z.; Tauscher, A.N.; Pagana, J.M.; Sherman, N.E.; Jeffery, E.D.; Spinelli, K.J.; Zhao, H.; et al. Molecular architecture of the chick vestibular hair bundle. Nat. Neurosci. 2013, 16, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Sekerkova, G.; Vranich, K.; Tilney, L.G.; Mugnaini, E.; Bartles, J.R. The deaf jerker mouse has a mutation in the gene encoding the espin actin-bundling proteins of hair cell stereocilia and lacks espins. Cell 2000, 102, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Lewis, A.K.; Bridgman, P.C. Nerve growth cone lamellipodia contain two populations of actin filaments that differ in organization and polarity. J. Cell Biol. 1992, 119, 1219–1243. [Google Scholar] [CrossRef]
- Gallop, J.L. Filopodia and their links with membrane traffic and cell adhesion. Semin. Cell Dev. Biol. 2020, 102, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Linder, S.; Cervero, P.; Eddy, R.; Condeelis, J. Mechanisms and roles of podosomes and invadopodia. Nat. Rev. Mol. Cell Biol. 2023, 24, 86–106. [Google Scholar] [CrossRef] [PubMed]
- Mellor, H. The role of formins in filopodia formation. Biochim. Biophys. Acta 2010, 1803, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.; Zhu, D.; Dong, H.; Zhang, P.; Xing, F.; Li, W.; Yan, R.; Zhou, J.; Xu, K.; Pan, L.; et al. Super-resolution microscopy reveals nanoscale architecture and regulation of podosome clusters in primary macrophages. iScience 2022, 25, 105514. [Google Scholar] [CrossRef] [PubMed]
- Alonso, F.; Spuul, P.; Daubon, T.; Kramer, I.; Genot, E. Variations on the theme of podosomes: A matter of context. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 545–553. [Google Scholar] [CrossRef]
- Murphy, D.A.; Courtneidge, S.A. The ‘ins’ and ‘outs’ of podosomes and invadopodia: Characteristics, formation and function. Nat. Rev. Mol. Cell Biol. 2011, 12, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Van Audenhove, I.; Debeuf, N.; Boucherie, C.; Gettemans, J. Fascin actin bundling controls podosome turnover and disassembly while cortactin is involved in podosome assembly by its SH3 domain in THP-1 macrophages and dendritic cells. Biochim. Biophys. Acta 2015, 1853, 940–952. [Google Scholar] [CrossRef]
- De Clercq, S.; Boucherie, C.; Vandekerckhove, J.; Gettemans, J.; Guillabert, A. L-plastin nanobodies perturb matrix degradation, podosome formation, stability and lifetime in THP-1 macrophages. PLoS ONE 2013, 8, e78108. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Mruk, D.D.; Wong, C.K.; Lee, W.M.; Han, D.; Cheng, C.Y. Actin-bundling protein plastin 3 is a regulator of ectoplasmic specialization dynamics during spermatogenesis in the rat testis. FASEB J. 2015, 29, 3788–3805. [Google Scholar] [CrossRef] [Green Version]
- Berruti, G.; Paiardi, C. The dynamic of the apical ectoplasmic specialization between spermatids and Sertoli cells: The case of the small GTPase Rap1. Biomed. Res. Int. 2014, 2014, 635979. [Google Scholar] [CrossRef]
- Kopera, I.A.; Bilinska, B.; Cheng, C.Y.; Mruk, D.D. Sertoli-germ cell junctions in the testis: A review of recent data. Philos. Trans. R Soc. B Biol. Sci. 2010, 365, 1593–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krey, J.F.; Krystofiak, E.S.; Dumont, R.A.; Vijayakumar, S.; Choi, D.; Rivero, F.; Kachar, B.; Jones, S.M.; Barr-Gillespie, P.G. Plastin 1 widens stereocilia by transforming actin filament packing from hexagonal to liquid. J. Cell Biol. 2016, 215, 467–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krey, J.F.; Barr-Gillespie, P.G. Molecular Composition of Vestibular Hair Bundles. Cold Spring Harb. Perspect. Med. 2019, 9, a033209. [Google Scholar] [CrossRef]
- Blanchoin, L.; Boujemaa-Paterski, R.; Sykes, C.; Plastino, J. Actin dynamics, architecture, and mechanics in cell motility. Physiol. Rev. 2014, 94, 235–263. [Google Scholar] [CrossRef] [Green Version]
- Tojkander, S.; Gateva, G.; Lappalainen, P. Actin stress fibers--assembly, dynamics and biological roles. J. Cell Sci. 2012, 125, 1855–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laporte, D.; Ojkic, N.; Vavylonis, D.; Wu, J.Q. Alpha-Actinin and fimbrin cooperate with myosin II to organize actomyosin bundles during contractile-ring assembly. Mol. Biol. Cell 2012, 23, 3094–3110. [Google Scholar] [CrossRef]
- Skau, C.T.; Courson, D.S.; Bestul, A.J.; Winkelman, J.D.; Rock, R.S.; Sirotkin, V.; Kovar, D.R. Actin filament bundling by fimbrin is important for endocytosis, cytokinesis, and polarization in fission yeast. J. Biol. Chem. 2011, 286, 26964–26977. [Google Scholar] [CrossRef] [Green Version]
- Mei, L.; Reynolds, M.J.; Garbett, D.; Gong, R.; Meyer, T.; Alushin, G.M. Structural mechanism for bidirectional actin cross-linking by T-plastin. Proc. Natl. Acad. Sci. USA 2022, 119, e2205370119. [Google Scholar] [CrossRef] [PubMed]
- Hotulainen, P.; Lappalainen, P. Stress fibers are generated by two distinct actin assembly mechanisms in motile cells. J. Cell Biol. 2006, 173, 383–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letort, G.; Ennomani, H.; Gressin, L.; Théry, M.; Blanchoin, L. Dynamic reorganization of the actin cytoskeleton. F1000Res. 2015, 4, 940. [Google Scholar] [CrossRef] [Green Version]
- Cramer, L.P.; Siebert, M.; Mitchison, T.J. Identification of novel graded polarity actin filament bundles in locomoting heart fibroblasts: Implications for the generation of motile force. J. Cell Biol. 1997, 136, 1287–1305. [Google Scholar] [CrossRef]
- Burnette, D.T.; Manley, S.; Sengupta, P.; Sougrat, R.; Davidson, M.W.; Kachar, B.; Lippincott-Schwartz, J. A role for actin arcs in the leading-edge advance of migrating cells. Nat. Cell Biol. 2011, 13, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Khatau, S.B.; Hale, C.M.; Stewart-Hutchinson, P.J.; Patel, M.S.; Stewart, C.L.; Searson, P.C.; Hodzic, D.; Wirtz, D. A perinuclear actin cap regulates nuclear shape. Proc. Natl. Acad. Sci. USA 2009, 106, 19017–19022. [Google Scholar] [CrossRef] [Green Version]
- Pfisterer, K.; Jayo, A.; Parsons, M. Control of nuclear organization by F-actin binding proteins. Nucleus 2017, 8, 126–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasbiswas, K.; Hu, S.; Schnorrer, F.; Safran, S.A.; Bershadsky, A.D. Ordering of myosin II filaments driven by mechanical forces: Experiments and theory. Philos. Trans. R Soc. Lond. B Biol. Sci. 2018, 373, 20170114. [Google Scholar] [CrossRef] [Green Version]
- Fenix, A.M.; Neininger, A.C.; Taneja, N.; Hyde, K.; Visetsouk, M.R.; Garde, R.J.; Liu, B.; Nixon, B.R.; Manalo, A.E.; Becker, J.R.; et al. Muscle-specific stress fibers give rise to sarcomeres in cardiomyocytes. Elife 2018, 7, e42144. [Google Scholar] [CrossRef]
- Mangione, M.C.; Gould, K.L. Molecular form and function of the cytokinetic ring. J. Cell Sci. 2019, 132, jcs226928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, C.M.; Alberts, B.M. Anillin, a contractile ring protein that cycles from the nucleus to the cell cortex. J. Cell Biol. 1995, 131, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Sugawa, M.; Yamagishi, M.; Kodera, N.; Yajima, J. Visualizing dynamic actin cross-linking processes driven by the actin-binding protein anillin. FEBS Lett. 2020, 594, 1237–1247. [Google Scholar] [CrossRef]
- Chugh, P.; Paluch, E.K. The actin cortex at a glance. J. Cell Sci. 2018, 131, jcs186254. [Google Scholar] [CrossRef] [Green Version]
- Svitkina, T.M. Ultrastructure of the actin cytoskeleton. Curr. Opin. Cell Biol. 2018, 54, 1–8. [Google Scholar] [CrossRef]
- Tilney, L.G.; Connelly, P.S.; Vranich, K.A.; Shaw, M.K.; Guild, G.M. Why are two different cross-linkers necessary for actin bundle formation in vivo and what does each cross-link contribute? J. Cell Biol. 1998, 143, 121–133. [Google Scholar] [CrossRef]
- Sauvanet, C.; Wayt, J.; Pelaseyed, T.; Bretscher, A. Structure, regulation, and functional diversity of microvilli on the apical domain of epithelial cells. Annu. Rev. Cell Dev. Biol. 2015, 31, 593–621. [Google Scholar] [CrossRef]
- Revenu, C.; Ubelmann, F.; Hurbain, I.; El-Marjou, F.; Dingli, F.; Loew, D.; Delacour, D.; Gilet, J.; Brot-Laroche, E.; Rivero, F.; et al. A new role for the architecture of microvillar actin bundles in apical retention of membrane proteins. Mol. Biol. Cell 2012, 23, 324–336. [Google Scholar] [CrossRef]
- Ezzell, R.M.; Chafel, M.M.; Matsudaira, P.T. Differential localization of villin and fimbrin during development of the mouse visceral endoderm and intestinal epithelium. Development 1989, 106, 407–419. [Google Scholar] [CrossRef]
- Schwebach, C.L.; Kudryashova, E.; Kudryashov, D.S. Plastin 3 in X-Linked Osteoporosis: Imbalance of Ca(2+)-Dependent Regulation Is Equivalent to Protein Loss. Front. Cell Dev. Biol. 2020, 8, 635783. [Google Scholar] [CrossRef]
- Bashirzadeh, Y.; Wubshet, N.H.; Liu, A.P. Confinement Geometry Tunes Fascin-Actin Bundle Structures and Consequently the Shape of a Lipid Bilayer Vesicle. Front. Mol. Biosci. 2020, 7, 610277. [Google Scholar] [CrossRef]
- Matsudaira, P. Modular organization of actin crosslinking proteins. Trends Biochem. Sci. 1991, 16, 87–92. [Google Scholar] [CrossRef]
- Winder, S.J.; Ayscough, K.R. Actin-binding proteins. J. Cell Sci. 2005, 118, 651–654. [Google Scholar] [CrossRef] [Green Version]
- Meyer, R.K.; Aebi, U. Bundling of actin filaments by alpha-actinin depends on its molecular length. J. Cell Biol. 1990, 110, 2013–2024. [Google Scholar] [CrossRef] [Green Version]
- Jansen, S.; Collins, A.; Yang, C.; Rebowski, G.; Svitkina, T.; Dominguez, R. Mechanism of actin filament bundling by fascin. J. Biol. Chem. 2011, 286, 30087–30096. [Google Scholar] [CrossRef] [Green Version]
- Tseng, Y.; Wirtz, D. Mechanics and multiple-particle tracking microheterogeneity of alpha-actinin-cross-linked actin filament networks. Biophys. J. 2001, 81, 1643–1656. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, O.; Pokidysheva, E.; Hirst, L.S.; Bouxsein, N.; Li, Y.; Safinya, C.R. Structure of actin cross-linked with alpha-actinin: A network of bundles. Phys. Rev. Lett. 2003, 91, 148102. [Google Scholar] [CrossRef]
- Tseng, Y.; An, K.M.; Esue, O.; Wirtz, D. The bimodal role of filamin in controlling the architecture and mechanics of F-actin networks. J. Biol. Chem. 2004, 279, 1819–1826. [Google Scholar] [CrossRef] [Green Version]
- Broderick, M.J.; Winder, S.J. Spectrin, α-actinin, and dystrophin. Adv. Protein Chem. 2005, 70, 203–246. [Google Scholar] [CrossRef]
- Murphy, A.C.; Young, P.W. The actinin family of actin cross-linking proteins—A genetic perspective. Cell Biosci. 2015, 5, 49. [Google Scholar] [CrossRef] [Green Version]
- Taylor, K.A.; Taylor, D.W.; Schachat, F. Isoforms of alpha-actinin from cardiac, smooth, and skeletal muscle form polar arrays of actin filaments. J. Cell Biol. 2000, 149, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Ciobanasu, C.; Faivre, B.; Le Clainche, C. Actin dynamics associated with focal adhesions. Int. J. Cell Biol. 2012, 2012, 941292. [Google Scholar] [CrossRef] [Green Version]
- Stachowiak, M.R.; McCall, P.M.; Thoresen, T.; Balcioglu, H.E.; Kasiewicz, L.; Gardel, M.L.; O’Shaughnessy, B. Self-organization of myosin II in reconstituted actomyosin bundles. Biophys. J. 2012, 103, 1265–1274. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Dasbiswas, K.; Guo, Z.; Tee, Y.H.; Thiagarajan, V.; Hersen, P.; Chew, T.L.; Safran, S.A.; Zaidel-Bar, R.; Bershadsky, A.D. Long-range self-organization of cytoskeletal myosin II filament stacks. Nat. Cell Biol. 2017, 19, 133–141. [Google Scholar] [CrossRef]
- Foley, K.S.; Young, P.W. The non-muscle functions of actinins: An update. Biochem. J. 2014, 459, 1–13. [Google Scholar] [CrossRef]
- Tseng, C.C.; Zheng, R.H.; Lin, T.W.; Chou, C.C.; Shih, Y.C.; Liang, S.W.; Lee, H.H. α-Actinin-4 recruits Shp2 into focal adhesions to potentiate ROCK2 activation in podocytes. Life Sci. Alliance 2022, 5, e202201557. [Google Scholar] [CrossRef]
- Kanhai, D.; Mulder, R.; van Amstel, H.K.P.; Schutgens, R.; Lukens, M.; Tamminga, R.Y.J. Familial macrothrombocytopenia due to a double mutation in cis in the alpha-actinin 1 gene (ACTN1], previously considered to be chronic immune thrombocytopenic purpura. Pediatr. Blood Cancer 2018, 65, e274182018. [Google Scholar] [CrossRef]
- O’Sullivan, L.R.; Cahill, M.R.; Young, P.W. The Importance of Alpha-Actinin Proteins in Platelet Formation and Function, and Their Causative Role in Congenital Macrothrombocytopenia. Int. J. Mol. Sci 2021, 22, 9363. [Google Scholar] [CrossRef]
- Lindholm, M.E.; Jimenez-Morales, D.; Zhu, H.; Seo, K.; Amar, D.; Zhao, C.; Raja, A.; Madhvani, R.; Abramowitz, S.; Espenel, C.; et al. Mono-and Biallelic Protein-Truncating Variants in Alpha-Actinin 2 Cause Cardiomyopathy Through Distinct Mechanisms. Circ. Genom. Precis. Med. 2021, 14, e003419. [Google Scholar] [CrossRef]
- Kaplan, J.M.; Kim, S.H.; North, K.N.; Rennke, H.; Correia, L.A.; Tong, H.Q.; Mathis, B.J.; Rodríguez-Pérez, J.C.; Allen, P.G.; Beggs, A.H.; et al. Mutations in ACTN4, encoding α-actinin-4, cause familial focal segmental glomerulosclerosis. Nat. Genet. 2000, 24, 251–256. [Google Scholar] [CrossRef]
- Yang, C.; Glass, W.F., 2nd. Expression of α-actinin-1 in human glomerular mesangial cells in vivo and in vitro. Exp. Biol. Med. 2008, 233, 689–693. [Google Scholar] [CrossRef]
- Hsu, K.S.; Kao, H.Y. Alpha-actinin 4 and tumorigenesis of breast cancer. Vitam. Horm. 2013, 93, 323–351. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, S.; Tsuda, H.; Honda, K.; Onozato, K.; Takano, M.; Tamai, S.; Imoto, I.; Inazawa, J.; Yamada, T.; Matsubara, O. Actinin-4 gene amplification in ovarian cancer: A candidate oncogene associated with poor patient prognosis and tumor chemoresistance. Mod. Pathol. 2009, 22, 499–507. [Google Scholar] [CrossRef] [Green Version]
- Oikonomou, K.G.; Zachou, K.; Dalekos, G.N. Alpha-actinin: A multidisciplinary protein with important role in B-cell driven autoimmunity. Autoimmun. Rev. 2011, 10, 389–396. [Google Scholar] [CrossRef]
- Kovac, B.; Teo, J.L.; Mäkelä, T.P.; Vallenius, T. Assembly of non-contractile dorsal stress fibers requires α-actinin-1 and Rac1 in migrating and spreading cells. J. Cell Sci. 2013, 126, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.; Wang, J.H.; Pollak, M.R.; Wells, A. α-actinin-4 is essential for maintaining the spreading, motility and contractility of fibroblasts. PLoS ONE 2010, 5, e13921. [Google Scholar] [CrossRef] [Green Version]
- Khurana, S.; Chakraborty, S.; Lam, M.; Liu, Y.; Su, Y.T.; Zhao, X.; Saleem, M.A.; Mathieson, P.W.; Bruggeman, L.A.; Kao, H.Y. Familial focal segmental glomerulosclerosis (FSGS)-linked α-actinin 4 (ACTN4] protein mutants lose ability to activate transcription by nuclear hormone receptors. J. Biol. Chem. 2012, 287, 12027–12035. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Khurana, S.; Charkraborty, S.; Tian, Y.; Sedor, J.R.; Bruggman, L.A.; Kao, H.Y. α Actinin 4 (ACTN4] Regulates Glucocorticoid Receptor-mediated Transactivation and Transrepression in Podocytes. J. Biol. Chem. 2017, 292, 1637–1647. [Google Scholar] [CrossRef] [Green Version]
- Murphy, A.C.; Lindsay, A.J.; McCaffrey, M.W.; Djinović-Carugo, K.; Young, P.W. Congenital macrothrombocytopenia-linked mutations in the actin-binding domain of α-actinin-1 enhance F-actin association. FEBS Lett. 2016, 590, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Yasutomi, M.; Kunishima, S.; Okazaki, S.; Tanizawa, A.; Tsuchida, S.; Ohshima, Y. ACTN1 rod domain mutation associated with congenital macrothrombocytopenia. Ann. Hematol. 2016, 95, 141–144. [Google Scholar] [CrossRef]
- Shao, H.; Wingert, B.; Weins, A.; Pollak, M.R.; Camacho, C.; Wells, A. Focal segmental glomerulosclerosis ACTN4 mutants binding to actin: Regulation by phosphomimetic mutations. Sci. Rep. 2019, 9, 15517. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; MacArthur, D.G.; Gulbin, J.P.; Hahn, A.G.; Beggs, A.H.; Easteal, S.; North, K. ACTN3 genotype is associated with human elite athletic performance. Am. J. Hum. Genet. 2003, 73, 627–631. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Grange, M.; Wagner, T.; Kho, A.L.; Gautel, M.; Raunser, S. The molecular basis for sarcomere organization in vertebrate skeletal muscle. Cell 2021, 184, 2135–2150.e2113. [Google Scholar] [CrossRef]
- Franzot, G.; Sjöblom, B.; Gautel, M.; Djinović Carugo, K. The crystal structure of the actin binding domain from alpha-actinin in its closed conformation: Structural insight into phospholipid regulation of α-actinin. J. Mol. Biol. 2005, 348, 151–165. [Google Scholar] [CrossRef]
- Hampton, C.M.; Taylor, D.W.; Taylor, K.A. Novel structures for α-actinin:F-actin interactions and their implications for actin-membrane attachment and tension sensing in the cytoskeleton. J. Mol. Biol. 2007, 368, 92–104. [Google Scholar] [CrossRef] [Green Version]
- McGough, A.; Way, M.; DeRosier, D. Determination of the alpha-actinin-binding site on actin filaments by cryoelectron microscopy and image analysis. J. Cell Biol. 1994, 126, 433–443. [Google Scholar] [CrossRef]
- Sjöblom, B.; Salmazo, A.; Djinović-Carugo, K. α-actinin structure and regulation. Cell. Mol. Life Sci. 2008, 65, 2688–2701. [Google Scholar] [CrossRef]
- Burridge, K.; Feramisco, J.R. Non-muscle alpha actinins are calcium-sensitive actin-binding proteins. Nature 1981, 294, 565–567. [Google Scholar] [CrossRef]
- Flood, G.; Kahana, E.; Gilmore, A.P.; Rowe, A.J.; Gratzer, W.B.; Critchley, D.R. Association of structural repeats in the α-actinin rod domain. Alignment of inter-subunit interactions. J. Mol. Biol. 1995, 252, 227–234. [Google Scholar] [CrossRef]
- Flood, G.; Rowe, A.J.; Critchley, D.R.; Gratzer, W.B. Further analysis of the role of spectrin repeat motifs in α-actinin dimer formation. Eur. Biophys. J. 1997, 25, 431–435. [Google Scholar] [CrossRef]
- Ribeiro, E.A., Jr.; Pinotsis, N.; Ghisleni, A.; Salmazo, A.; Konarev, P.V.; Kostan, J.; Sjöblom, B.; Schreiner, C.; Polyansky, A.A.; Gkougkoulia, E.A.; et al. The structure and regulation of human muscle α-actinin. Cell 2014, 159, 1447–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrego-Diaz, E.; Kerff, F.; Lee, S.H.; Ferron, F.; Li, Y.; Dominguez, R. Crystal structure of the actin-binding domain of α-actinin 1: Evaluating two competing actin-binding models. J. Struct. Biol. 2006, 155, 230–238. [Google Scholar] [CrossRef]
- Kuhlman, P.A.; Hemmings, L.; Critchley, D.R. The identification and characterisation of an actin-binding site in α-actinin by mutagenesis. FEBS Lett. 1992, 304, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, A.; Ohanian, V.; Critchley, D. The structure and function of α-actinin. J. Muscle Res. Cell Motil. 1989, 10, 280–289. [Google Scholar] [CrossRef]
- Liu, J.; Taylor, D.W.; Taylor, K.A. A 3-D reconstruction of smooth muscle α-actinin by CryoEm reveals two different conformations at the actin-binding region. J. Mol. Biol. 2004, 338, 115–125. [Google Scholar] [CrossRef]
- Lee, S.H.; Weins, A.; Hayes, D.B.; Pollak, M.R.; Dominguez, R. Crystal structure of the actin-binding domain of α-actinin-4 Lys255Glu mutant implicated in focal segmental glomerulosclerosis. J. Mol. Biol. 2008, 376, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galkin, V.E.; Orlova, A.; Salmazo, A.; Djinovic-Carugo, K.; Egelman, E.H. Opening of tandem calponin homology domains regulates their affinity for F-actin. Nat. Struct. Mol. Biol. 2010, 17, 614–616. [Google Scholar] [CrossRef]
- Winkelman, J.D.; Suarez, C.; Hocky, G.M.; Harker, A.J.; Morganthaler, A.N.; Christensen, J.R.; Voth, G.A.; Bartles, J.R.; Kovar, D.R. Fascin- and α-Actinin-Bundled Networks Contain Intrinsic Structural Features that Drive Protein Sorting. Curr. Biol. 2016, 26, 2697–2706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oda, T.; Yanagisawa, H. Cryo-electron tomography of cardiac myofibrils reveals a 3D lattice spring within the Z-discs. Commun. Biol. 2020, 3, 585. [Google Scholar] [CrossRef]
- Pinotsis, N.; Zielinska, K.; Babuta, M.; Arolas, J.L.; Kostan, J.; Khan, M.B.; Schreiner, C.; Salmazo, A.; Ciccarelli, L.; Puchinger, M.; et al. Calcium modulates the domain flexibility and function of an α-actinin similar to the ancestral α-actinin. Proc. Natl. Acad. Sci. USA 2020, 117, 22101–22112. [Google Scholar] [CrossRef] [PubMed]
- Corgan, A.M.; Singleton, C.; Santoso, C.B.; Greenwood, J.A. Phosphoinositides differentially regulate alpha-actinin flexibility and function. Biochem. J. 2004, 378, 1067–1072. [Google Scholar] [CrossRef] [Green Version]
- Fraley, T.S.; Pereira, C.B.; Tran, T.C.; Singleton, C.; Greenwood, J.A. Phosphoinositide binding regulates α-actinin dynamics: Mechanism for modulating cytoskeletal remodeling. J. Biol. Chem. 2005, 280, 15479–15482. [Google Scholar] [CrossRef] [Green Version]
- Fraley, T.S.; Tran, T.C.; Corgan, A.M.; Nash, C.A.; Hao, J.; Critchley, D.R.; Greenwood, J.A. Phosphoinositide binding inhibits α-actinin bundling activity. J. Biol. Chem. 2003, 278, 24039–24045. [Google Scholar] [CrossRef] [Green Version]
- Izaguirre, G.; Aguirre, L.; Hu, Y.P.; Lee, H.Y.; Schlaepfer, D.D.; Aneskievich, B.J.; Haimovich, B. The cytoskeletal/non-muscle isoform of α-actinin is phosphorylated on its actin-binding domain by the focal adhesion kinase. J. Biol. Chem. 2001, 276, 28676–28685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, H.; Wu, C.; Wells, A. Phosphorylation of α-actinin 4 upon epidermal growth factor exposure regulates its interaction with actin. J. Biol. Chem. 2010, 285, 2591–2600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprague, C.R.; Fraley, T.S.; Jang, H.S.; Lal, S.; Greenwood, J.A. Phosphoinositide binding to the substrate regulates susceptibility to proteolysis by calpain. J. Biol. Chem. 2008, 283, 9217–9223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raynaud, F.; Bonnal, C.; Fernandez, E.; Bremaud, L.; Cerutti, M.; Lebart, M.C.; Roustan, C.; Ouali, A.; Benyamin, Y. The calpain 1-α-actinin interaction. Resting complex between the calcium-dependent protease and its target in cytoskeleton. Eur. J. Biochem. 2003, 270, 4662–4670. [Google Scholar] [CrossRef]
- Chopra, A.; Kutys, M.L.; Zhang, K.; Polacheck, W.J.; Sheng, C.C.; Luu, R.J.; Eyckmans, J.; Hinson, J.T.; Seidman, J.G.; Seidman, C.E.; et al. Force Generation via β-Cardiac Myosin, Titin, and α-Actinin Drives Cardiac Sarcomere Assembly from Cell-Matrix Adhesions. Dev. Cell 2018, 44, 87–96.e5. [Google Scholar] [CrossRef] [Green Version]
- Young, P.; Ferguson, C.; Banuelos, S.; Gautel, M. Molecular structure of the sarcomeric Z-disk: Two types of titin interactions lead to an asymmetrical sorting of alpha-actinin. EMBO J. 1998, 17, 1614–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, P.; Gautel, M. The interaction of titin and alpha-actinin is controlled by a phospholipid-regulated intramolecular pseudoligand mechanism. EMBO J. 2000, 19, 6331–6340. [Google Scholar] [CrossRef]
- Luther, P.K.; Squire, J.M. Muscle Z-band ultrastructure: Titin Z-repeats and Z-band periodicities do not match. J. Mol. Biol. 2002, 319, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Partynska, A.; Gomulkiewicz, A.; Dziegiel, P.; Podhorska-Okolow, M. The Role of Zyxin in Carcinogenesis. Anticancer Res. 2020, 40, 5981–5988. [Google Scholar] [CrossRef]
- Bauer, K.; Kratzer, M.; Otte, M.; de Quintana, K.L.; Hagmann, J.; Arnold, G.J.; Eckerskorn, C.; Lottspeich, F.; Siess, W. Human CLP36, a PDZ-domain and LIM-domain protein, binds to α-actinin-1 and associates with actin filaments and stress fibers in activated platelets and endothelial cells. Blood 2000, 96, 4236–4245. [Google Scholar] [CrossRef]
- Otey, C.A.; Carpen, O. Alpha-actinin revisited: A fresh look at an old player. Cell Motil. Cytoskelet. 2004, 58, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Burton, K.M.; Cao, H.; Chen, J.; Qiang, L.; Krueger, E.W.; Johnson, K.M.; Bamlet, W.R.; Zhang, L.; McNiven, M.A.; Razidlo, G.L. Dynamin 2 interacts with α-actinin 4 to drive tumor cell invasion. Mol. Biol. Cell 2020, 31, 439–451. [Google Scholar] [CrossRef]
- Trulsson, M.; Yu, H.; Gisselsson, L.; Chao, Y.; Urbano, A.; Aits, S.; Mossberg, A.K.; Svanborg, C. HAMLET binding to α-actinin facilitates tumor cell detachment. PLoS ONE 2011, 6, e17179. [Google Scholar] [CrossRef] [Green Version]
- Aksenova, V.; Turoverova, L.; Khotin, M.; Magnusson, K.E.; Tulchinsky, E.; Melino, G.; Pinaev, G.P.; Barlev, N.; Tentler, D. Actin-binding protein alpha-actinin 4 (ACTN4] is a transcriptional co-activator of RelA/p65 sub-unit of NF-kB. Oncotarget 2013, 4, 362–372. [Google Scholar] [CrossRef] [Green Version]
- Bretscher, A.; Weber, K. Fimbrin, a new microfilament-associated protein present in microvilli and other cell surface structures. J. Cell Biol. 1980, 86, 335–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arpin, M.; Friederich, E.; Algrain, M.; Vernel, F.; Louvard, D. Functional differences between L- and T-plastin isoforms. J. Cell Biol. 1994, 127, 1995–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretscher, A. Fimbrin is a cytoskeletal protein that crosslinks F-actin in vitro. Proc. Natl. Acad. Sci. USA 1981, 78, 6849–6853. [Google Scholar] [CrossRef] [Green Version]
- Delanote, V.; Vandekerckhove, J.; Gettemans, J. Plastins: Versatile modulators of actin organization in (patho)physiological cellular processes. Acta Pharmacol. Sin. 2005, 26, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Leavitt, J.; Goldman, D.; Merril, C.; Kakunaga, T. Changes in gene expression accompanying chemically-induced malignant transformation of human fibroblasts. Carcinogenesis 1982, 3, 61–70. [Google Scholar] [CrossRef]
- Leavitt, J.; Kakunaga, T. Expression of a variant form of actin and additional polypeptide changes following chemical-induced in vitro neoplastic transformation of human fibroblasts. J. Biol. Chem. 1980, 255, 1650–1661. [Google Scholar] [CrossRef]
- Lin, C.S.; Park, T.; Chen, Z.P.; Leavitt, J. Human plastin genes. Comparative gene structure, chromosome location, and differential expression in normal and neoplastic cells. J. Biol. Chem. 1993, 268, 2781–2792. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, H.; Yamada, M.; Noguchi, A.; Fujii, K.; Shibasaki, M.; Mukai, R.; Yabe, T.; Kondo, I. Genetic analysis of human lymphocyte proteins by two-dimensional gel electrophoresis: 2. Genetic polymorphism of lymphocyte cytosol 64K polypeptide. Hum. Genet. 1982, 60, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, D.; Djeu, J.; Latter, G.; Burbeck, S.; Leavitt, J. Abundant synthesis of the transformation-induced protein of neoplastic human fibroblasts, plastin, in normal lymphocytes. Cancer Res. 1985, 45, 5643–5647. [Google Scholar] [PubMed]
- Shinomiya, H. Plastin family of actin-bundling proteins: Its functions in leukocytes, neurons, intestines, and cancer. Int. J. Cell Biol 2012, 2012, 213492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Horta, O.; Bademci, G.; Tokgoz-Yilmaz, S.; Guo, S.; Zafeer, F.; Sineni, C.J.; Duman, D.; Farooq, A.; Tekin, M. Novel variant p.E269K confirms causative role of PLS1 mutations in autosomal dominant hearing loss. Clin. Genet. 2019, 96, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.; Bullen, A.; Johnson, S.L.; Grimm-Gunter, E.M.; Rivero, F.; Marcotti, W.; Forge, A.; Daudet, N. Absence of plastin 1 causes abnormal maintenance of hair cell stereocilia and a moderate form of hearing loss in mice. Hum. Mol. Genet. 2015, 24, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Grimm-Gunter, E.M.; Revenu, C.; Ramos, S.; Hurbain, I.; Smyth, N.; Ferrary, E.; Louvard, D.; Robine, S.; Rivero, F. Plastin 1 binds to keratin and is required for terminal web assembly in the intestinal epithelium. Mol. Biol. Cell 2009, 20, 2549–2562. [Google Scholar] [CrossRef] [Green Version]
- Wabnitz, G.; Balta, E.; Samstag, Y. L-plastin regulates the stability of the immune synapse of naive and effector T-cells. Adv. Biol. Regul. 2017, 63, 107–114. [Google Scholar] [CrossRef]
- Bhatlekar, S.; Manne, B.K.; Basak, I.; Edelstein, L.C.; Tugolukova, E.; Stoller, M.L.; Cody, M.J.; Morley, S.C.; Nagalla, S.; Weyrich, A.S.; et al. miR-125a-5p regulates megakaryocyte proplatelet formation via the actin-bundling protein L-plastin. Blood 2020, 136, 1760–1772. [Google Scholar] [CrossRef]
- Joshi, H.; Morley, S.C. Efficient T Cell Migration and Activation Require L-Plastin. Front. Immunol. 2022, 13, 916137. [Google Scholar] [CrossRef] [PubMed]
- Joshi, H.; Almgren-Bell, A.; Anaya, E.P.; Todd, E.M.; Van Dyken, S.J.; Seth, A.; McIntire, K.M.; Singamaneni, S.; Sutterwala, F.; Morley, S.C. L-plastin enhances NLRP3 inflammasome assembly and bleomycin-induced lung fibrosis. Cell Rep. 2022, 38, 110507. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.Y.; Szasz, T.P.; Stewart-Hutchinson, P.J.; Sivapalan, J.; Todd, E.M.; Deady, L.E.; Cooper, J.A.; Onken, M.D.; Morley, S.C. L-Plastin promotes podosome longevity and supports macrophage motility. Mol. Immunol. 2016, 78, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Galiegue-Zouitina, S.; Quief, S.; Hildebrand, M.P.; Denis, C.; Detourmignies, L.; Lai, J.L.; Kerckaert, J.P. Nonrandom fusion of L-plastin(LCP1] and LAZ3(BCL6] genes by t[3;13](q27;q14] chromosome translocation in two cases of B-cell non-Hodgkin lymphoma. Genes Chromosomes Cancer 1999, 26, 97–105. [Google Scholar] [CrossRef]
- Samstag, Y.; Klemke, M. Ectopic expression of L-plastin in human tumor cells: Diagnostic and therapeutic implications. Adv. Enzym. Regul. 2007, 47, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chafel, M.M.; Shen, W.; Matsudaira, P. Sequential expression and differential localization of I-, L-, and T-fimbrin during differentiation of the mouse intestine and yolk sac. Dev. Dyn. 1995, 203, 141–151. [Google Scholar] [CrossRef]
- Petit, F.; Longoni, M.; Wells, J.; Maser, R.; Dysart, M.J.; Contreras, H.T.M.; Frénois, F.; Bogenschutz, E.; Pober, B.R.; Clark, R.D.; et al. Missense variants affecting the actin-binding domains of PLS3 cause X-linked congenital diaphragmatic hernia and body wall defects. medRxiv 2021. preprint. [Google Scholar] [CrossRef]
- Dor-On, E.; Raviv, S.; Cohen, Y.; Adir, O.; Padmanabhan, K.; Luxenburg, C. T-plastin is essential for basement membrane assembly and epidermal morphogenesis. Sci. Signal. 2017, 10, eaal3154. [Google Scholar] [CrossRef] [PubMed]
- Hosseinibarkooie, S.; Peters, M.; Torres-Benito, L.; Rastetter, R.H.; Hupperich, K.; Hoffmann, A.; Mendoza-Ferreira, N.; Kaczmarek, A.; Janzen, E.; Milbradt, J.; et al. The Power of Human Protective Modifiers: PLS3 and CORO1C Unravel Impaired Endocytosis in Spinal Muscular Atrophy and Rescue SMA Phenotype. Am. J. Hum. Genet. 2016, 99, 647–665. [Google Scholar] [CrossRef] [Green Version]
- Walsh, M.B.; Janzen, E.; Wingrove, E.; Hosseinibarkooie, S.; Muela, N.R.; Davidow, L.; Dimitriadi, M.; Norabuena, E.M.; Rubin, L.L.; Wirth, B.; et al. Genetic modifiers ameliorate endocytic and neuromuscular defects in a model of spinal muscular atrophy. BMC Biol. 2020, 18, 127. [Google Scholar] [CrossRef]
- Lyon, A.N.; Pineda, R.H.; Hao, L.T.; Kudryashova, E.; Kudryashov, D.S.; Beattie, C.E. Calcium binding is essential for plastin 3 function in Smn-deficient motoneurons. Hum. Mol. Genet. 2014, 23, 1990–2004. [Google Scholar] [CrossRef] [Green Version]
- Begue, E.; Jean-Louis, F.; Bagot, M.; Jauliac, S.; Cayuela, J.M.; Laroche, L.; Parquet, N.; Bachelez, H.; Bensussan, A.; Courtois, G.; et al. Inducible expression and pathophysiologic functions of T-plastin in cutaneous T-cell lymphoma. Blood 2012, 120, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Velthaus, A.; Cornils, K.; Hennigs, J.K.; Grub, S.; Stamm, H.; Wicklein, D.; Bokemeyer, C.; Heuser, M.; Windhorst, S.; Fiedler, W.; et al. The Actin Binding Protein Plastin-3 Is Involved in the Pathogenesis of Acute Myeloid Leukemia. Cancers 2019, 11, 1663. [Google Scholar] [CrossRef] [Green Version]
- Xiong, F.; Wu, G.H.; Wang, B.; Chen, Y.J. Plastin-3 is a diagnostic and prognostic marker for pancreatic adenocarcinoma and distinguishes from diffuse large B-cell lymphoma. Cancer Cell Int. 2021, 21, 411. [Google Scholar] [CrossRef]
- Xin, Z.; Li, D.; Mao, F.; Du, Y.; Wang, X.; Xu, P.; Li, Z.; Qian, J.; Yao, J. PLS3 predicts poor prognosis in pancreatic cancer and promotes cancer cell proliferation via PI3K/AKT signaling. J. Cell. Physiol. 2020, 235, 8416–8423. [Google Scholar] [CrossRef]
- Hanein, D.; Volkmann, N.; Goldsmith, S.; Michon, A.M.; Lehman, W.; Craig, R.; DeRosier, D.; Almo, S.; Matsudaira, P. An atomic model of fimbrin binding to F-actin and its implications for filament crosslinking and regulation. Nat. Struct. Biol. 1998, 5, 787–792. [Google Scholar] [CrossRef]
- Schwebach, C.L.; Kudryashova, E.; Agrawal, R.; Zheng, W.; Egelman, E.H.; Kudryashov, D.S. Allosteric regulation controls actin-bundling properties of human plastins. Nat. Struct. Mol. Biol. 2022, 29, 519–528. [Google Scholar] [CrossRef]
- Klein, M.G.; Shi, W.; Ramagopal, U.; Tseng, Y.; Wirtz, D.; Kovar, D.R.; Staiger, C.J.; Almo, S.C. Structure of the actin crosslinking core of fimbrin. Structure 2004, 12, 999–1013. [Google Scholar] [CrossRef] [Green Version]
- Volkmann, N.; DeRosier, D.; Matsudaira, P.; Hanein, D. An atomic model of actin filaments cross-linked by fimbrin and its implications for bundle assembly and function. J. Cell Biol. 2001, 153, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Schwebach, C.L.; Kudryashova, E.; Zheng, W.; Orchard, M.; Smith, H.; Runyan, L.A.; Egelman, E.H.; Kudryashov, D.S. Osteogenesis imperfecta mutations in plastin 3 lead to impaired calcium regulation of actin bundling. Bone Res. 2020, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Schwebach, C.L.; Agrawal, R.; Lindert, S.; Kudryashova, E.; Kudryashov, D.S. The Roles of Actin-Binding Domains 1 and 2 in the Calcium-Dependent Regulation of Actin Filament Bundling by Human Plastins. J. Mol. Biol. 2017, 429, 2490–2508. [Google Scholar] [CrossRef] [PubMed]
- Galkin, V.E.; Orlova, A.; Cherepanova, O.; Lebart, M.C.; Egelman, E.H. High-resolution cryo-EM structure of the F-actin-fimbrin/plastin ABD2 complex. Proc. Natl. Acad. Sci. USA 2008, 105, 1494–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, H.; Jensen, K.V.; Woodman, A.G.; Hyndman, M.E.; Vogel, H.J. The Calcium-Dependent Switch Helix of L-Plastin Regulates Actin Bundling. Sci. Rep. 2017, 7, 40662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namba, Y.; Ito, M.; Zu, Y.; Shigesada, K.; Maruyama, K. Human T cell L-plastin bundles actin filaments in a calcium-dependent manner. J. Biochem. 1992, 112, 503–507. [Google Scholar] [CrossRef]
- Lin, C.S.; Shen, W.; Chen, Z.P.; Tu, Y.H.; Matsudaira, P. Identification of I-plastin, a human fimbrin isoform expressed in intestine and kidney. Mol. Cell. Biol. 1994, 14, 2457–2467. [Google Scholar] [CrossRef]
- Miyakawa, T.; Shinomiya, H.; Yumoto, F.; Miyauchi, Y.; Tanaka, H.; Ojima, T.; Kato, Y.S.; Tanokura, M. Different Ca²+- sensitivities between the EF-hands of T- and L-plastins. Biochem. Biophys. Res. Commun. 2012, 429, 137–141. [Google Scholar] [CrossRef]
- Shirayama, S.; Numata, O. Tetrahymena fimbrin localized in the division furrow bundles actin filaments in a calcium-independent manner. J. Biochem. 2003, 134, 591–598. [Google Scholar] [CrossRef]
- Kovar, D.R.; Staiger, C.J.; Weaver, E.A.; McCurdy, D.W. AtFim1 is an actin filament crosslinking protein from Arabidopsis thaliana. Plant J. 2000, 24, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Nakano, K.; Satoh, K.; Morimatsu, A.; Ohnuma, M.; Mabuchi, I. Interactions among a fimbrin, a capping protein, and an actin-depolymerizing factor in organization of the fission yeast actin cytoskeleton. Mol. Biol. Cell 2001, 12, 3515–3526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunning, P.; O’Neill, G.; Hardeman, E. Tropomyosin-based regulation of the actin cytoskeleton in time and space. Physiol. Rev. 2008, 88, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Christensen, J.R.; Hocky, G.M.; Homa, K.E.; Morganthaler, A.N.; Hitchcock-DeGregori, S.E.; Voth, G.A.; Kovar, D.R. Competition between Tropomyosin, Fimbrin, and ADF/Cofilin drives their sorting to distinct actin filament networks. Elife 2017, 6, e23152. [Google Scholar] [CrossRef]
- Skau, C.T.; Kovar, D.R. Fimbrin and tropomyosin competition regulates endocytosis and cytokinesis kinetics in fission yeast. Curr. Biol. 2010, 20, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Correia, I.; Chu, D.; Chou, Y.H.; Goldman, R.D.; Matsudaira, P. Integrating the actin and vimentin cytoskeletons. adhesion-dependent formation of fimbrin-vimentin complexes in macrophages. J. Cell Biol. 1999, 146, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Jayo, A.; Parsons, M. Fascin: A key regulator of cytoskeletal dynamics. Int. J. Biochem. Cell Biol. 2010, 42, 1614–1617. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Huang, F.K.; Huang, J.; Chen, S.; Jakoncic, J.; Leo-Macias, A.; Diaz-Avalos, R.; Chen, L.; Zhang, J.J.; Huang, X.Y.; et al. Molecular mechanism of fascin function in filopodial formation. J. Biol. Chem. 2013, 288, 274–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; Fujii, Y.; Hirano, K.; Higaki, T.; Nagasaki, A.; Ishikawa, R.; Okajima, T.; Katoh, K. Fascin in lamellipodia contributes to cell elasticity by controlling the orientation of filamentous actin. Genes Cells 2019, 24, 202–213. [Google Scholar] [CrossRef] [Green Version]
- Vignjevic, D.; Kojima, S.; Aratyn, Y.; Danciu, O.; Svitkina, T.; Borisy, G.G. Role of fascin in filopodial protrusion. J. Cell Biol. 2006, 174, 863–875. [Google Scholar] [CrossRef] [Green Version]
- Van Audenhove, I.; Denert, M.; Boucherie, C.; Pieters, L.; Cornelissen, M.; Gettemans, J. Fascin Rigidity and L-plastin Flexibility Cooperate in Cancer Cell Invadopodia and Filopodia. J. Biol. Chem. 2016, 291, 9148–9160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.L.; Pan, M.H.; Yang, F.L.; Zong, Z.A.; Tang, F.; Pan, Z.N.; Lu, X.; Ren, Y.P.; Wang, J.L.; Sun, S.C. FASCIN regulates actin assembly for spindle movement and polar body extrusion in mouse oocyte meiosis. J. Cell. Physiol. 2021, 236, 7725–7733. [Google Scholar] [CrossRef]
- Lamb, M.C.; Tootle, T.L. Fascin in Cell Migration: More Than an Actin Bundling Protein. Biology 2020, 9, 403. [Google Scholar] [CrossRef]
- Jayo, A.; Malboubi, M.; Antoku, S.; Chang, W.; Ortiz-Zapater, E.; Groen, C.; Pfisterer, K.; Tootle, T.; Charras, G.; Gundersen, G.G.; et al. Fascin Regulates Nuclear Movement and Deformation in Migrating Cells. Dev. Cell 2016, 38, 371–383. [Google Scholar] [CrossRef] [Green Version]
- Saad, A.; Bijian, K.; Qiu, D.; da Silva, S.D.; Marques, M.; Chang, C.H.; Nassour, H.; Ramotar, D.; Damaraju, S.; Mackey, J.; et al. Insights into a novel nuclear function for Fascin in the regulation of the amino-acid transporter SLC3A2. Sci. Rep. 2016, 6, 36699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, R.E. Preparation and purification of polymerized actin from sea urchin egg extracts. J. Cell Biol. 1975, 66, 305–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cant, K.; Knowles, B.A.; Mooseker, M.S.; Cooley, L. Drosophila singed, a fascin homolog, is required for actin bundle formation during oogenesis and bristle extension. J. Cell Biol. 1994, 125, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Otto, J.J.; Kane, R.E.; Bryan, J. Formation of filopodia in coelomocytes: Localization of fascin, a 58,000 dalton actin cross-linking protein. Cell 1979, 17, 285–293. [Google Scholar] [CrossRef]
- Yamashiro-Matsumura, S.; Matsumura, F. Purification and characterization of an F-actin-bundling 55-kilodalton protein from HeLa cells. J. Biol. Chem. 1985, 260, 5087–5097. [Google Scholar] [CrossRef]
- Yamashiro, S.; Yamakita, Y.; Ono, S.; Matsumura, F. Fascin, an actin-bundling protein, induces membrane protrusions and increases cell motility of epithelial cells. Mol. Biol. Cell 1998, 9, 993–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tubb, B.E.; Bardien-Kruger, S.; Kashork, C.D.; Shaffer, L.G.; Ramagli, L.S.; Xu, J.; Siciliano, M.J.; Bryan, J. Characterization of human retinal fascin gene (FSCN2) at 17q25: Close physical linkage of fascin and cytoplasmic actin genes. Genomics 2000, 65, 146–156. [Google Scholar] [CrossRef]
- Lin-Jones, J.; Burnside, B. Retina-specific protein fascin 2 is an actin cross-linker associated with actin bundles in photoreceptor inner segments and calycal processes. Invest. Ophthalmol. Vis. Sci. 2007, 48, 1380–1388. [Google Scholar] [CrossRef]
- Shin, J.B.; Longo-Guess, C.M.; Gagnon, L.H.; Saylor, K.W.; Dumont, R.A.; Spinelli, K.J.; Pagana, J.M.; Wilmarth, P.A.; David, L.L.; Gillespie, P.G.; et al. The R109H variant of fascin-2, a developmentally regulated actin crosslinker in hair-cell stereocilia, underlies early-onset hearing loss of DBA/2J mice. J. Neurosci. 2010, 30, 9683–9694. [Google Scholar] [CrossRef]
- Adams, J.C. Roles of fascin in cell adhesion and motility. Curr. Opin. Cell Biol. 2004, 16, 590–596. [Google Scholar] [CrossRef]
- Tubb, B.; Mulholland, D.J.; Vogl, W.; Lan, Z.J.; Niederberger, C.; Cooney, A.; Bryan, J. Testis fascin (FSCN3]: A novel paralog of the actin-bundling protein fascin expressed specifically in the elongate spermatid head. Exp. Cell Res. 2002, 275, 92–109. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Unar, A.; Dil, S.; Ali, I.; Khan, K.; Khan, I.; Shi, Q. Testis-specific fascin component FSCN3 is dispensable for mouse spermatogenesis and fertility. Mol. Biol. Rep. 2022, 49, 6261–6268. [Google Scholar] [CrossRef] [PubMed]
- Sedeh, R.S.; Fedorov, A.A.; Fedorov, E.V.; Ono, S.; Matsumura, F.; Almo, S.C.; Bathe, M. Structure, evolutionary conservation, and conformational dynamics of Homo sapiens fascin-1, an F-actin crosslinking protein. J. Mol. Biol. 2010, 400, 589–604. [Google Scholar] [CrossRef] [PubMed]
- Lamb, M.C.; Anliker, K.K.; Tootle, T.L. Fascin regulates protrusions and delamination to mediate invasive, collective cell migration in vivo. Dev. Dyn. 2020, 249, 961–982. [Google Scholar] [CrossRef] [PubMed]
- Aramaki, S.; Mayanagi, K.; Jin, M.; Aoyama, K.; Yasunaga, T. Filopodia formation by crosslinking of F-actin with fascin in two different binding manners. Cytoskeleton 2016, 73, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Yamakita, Y.; Ono, S.; Matsumura, F.; Yamashiro, S. Phosphorylation of human fascin inhibits its actin binding and bundling activities. J. Biol. Chem. 1996, 271, 12632–12638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villari, G.; Jayo, A.; Zanet, J.; Fitch, B.; Serrels, B.; Frame, M.; Stramer, B.M.; Goult, B.T.; Parsons, M. A direct interaction between fascin and microtubules contributes to adhesion dynamics and cell migration. J. Cell Sci. 2015, 128, 4601–4614. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Lu, S.; Mulaj, M.; Fang, B.; Keeley, T.; Wan, L.; Hao, J.; Muschol, M.; Sun, J.; Yang, S. Monoubiquitination Inhibits the Actin Bundling Activity of Fascin. J. Biol. Chem. 2016, 291, 27323–27333. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, R.; Yamashiro, S.; Kohama, K.; Matsumura, F. Regulation of actin binding and actin bundling activities of fascin by caldesmon coupled with tropomyosin. J. Biol. Chem. 1998, 273, 26991–26997. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, Y.; Hayashi, K.; Shirao, T.; Ishikawa, R.; Kohama, K. Inhibition by drebrin of the actin-bundling activity of brain fascin, a protein localized in filopodia of growth cones. J. Neurochem. 1996, 66, 980–988. [Google Scholar] [CrossRef]
- Elkhatib, N.; Neu, M.B.; Zensen, C.; Schmoller, K.M.; Louvard, D.; Bausch, A.R.; Betz, T.; Vignjevic, D.M. Fascin plays a role in stress fiber organization and focal adhesion disassembly. Curr. Biol. 2014, 24, 1492–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, M.C.; Kaluarachchi, C.P.; Lansakara, T.I.; Mellentine, S.Q.; Lan, Y.; Tivanski, A.V.; Tootle, T.L. Fascin limits Myosin activity within Drosophila border cells to control substrate stiffness and promote migration. Elife 2021, 10, e69836. [Google Scholar] [CrossRef] [PubMed]
- Breitsprecher, D.; Koestler, S.A.; Chizhov, I.; Nemethova, M.; Mueller, J.; Goode, B.L.; Small, J.V.; Rottner, K.; Faix, J. Cofilin cooperates with fascin to disassemble filopodial actin filaments. J. Cell Sci. 2011, 124, 3305–3318. [Google Scholar] [CrossRef] [Green Version]
- Harker, A.J.; Katkar, H.H.; Bidone, T.C.; Aydin, F.; Voth, G.A.; Applewhite, D.A.; Kovar, D.R. Ena/VASP processive elongation is modulated by avidity on actin filaments bundled by the filopodia cross-linker fascin. Mol. Biol. Cell 2019, 30, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Wan, X.; Wang, H.H.; Pan, Z.N.; Zhang, Y.; Sun, S.C. FMNL3 regulates FASCIN for actin-mediated spindle migration and cytokinesis in mouse oocytes. Biol. Reprod. 2020, 102, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, R.; Breitsprecher, D.; Collins, A.; Corrêa, I.R., Jr.; Xu, M.Q.; Goode, B.L. The formin Daam1 and fascin directly collaborate to promote filopodia formation. Curr. Biol. 2013, 23, 1373–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Fonovic, M.; Suyama, K.; Bogyo, M.; Scott, M.P. Rab35 controls actin bundling by recruiting fascin as an effector protein. Science 2009, 325, 1250–1254. [Google Scholar] [CrossRef] [PubMed]
- Adams, G., Jr.; López, M.P.; Cartagena-Rivera, A.X.; Waterman, C.M. Survey of cancer cell anatomy in nonadhesive confinement reveals a role for filamin-A and fascin-1 in leader bleb-based migration. Mol. Biol. Cell 2021, 32, 1772–1791. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Taylor, M.D.; Singh, P.K.; Yang, S. How does fascin promote cancer metastasis? FEBS J. 2021, 288, 1434–1446. [Google Scholar] [CrossRef]
- Darnel, A.D.; Behmoaram, E.; Vollmer, R.T.; Corcos, J.; Bijian, K.; Sircar, K.; Su, J.; Jiao, J.; Alaoui-Jamali, M.A.; Bismar, T.A. Fascin regulates prostate cancer cell invasion and is associated with metastasis and biochemical failure in prostate cancer. Clin. Cancer Res. 2009, 15, 1376–1383. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Dey, R.; Wang, Y.; Jakoncic, J.; Kurinov, I.; Huang, X.Y. Structural Insights into the Induced-fit Inhibition of Fascin by a Small-Molecule Inhibitor. J. Mol. Biol. 2018, 430, 1324–1335. [Google Scholar] [CrossRef]
- Chen, B.; Li, A.; Wang, D.; Wang, M.; Zheng, L.; Bartles, J.R. Espin contains an additional actin-binding site in its N terminus and is a major actin-bundling protein of the Sertoli cell-spermatid ectoplasmic specialization junctional plaque. Mol. Biol. Cell 1999, 10, 4327–4339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekerkova, G.; Zheng, L.; Loomis, P.A.; Changyaleket, B.; Whitlon, D.S.; Mugnaini, E.; Bartles, J.R. Espins are multifunctional actin cytoskeletal regulatory proteins in the microvilli of chemosensory and mechanosensory cells. J. Neurosci. 2004, 24, 5445–5456. [Google Scholar] [CrossRef] [Green Version]
- Sekerkova, G.; Zheng, L.; Loomis, P.A.; Mugnaini, E.; Bartles, J.R. Espins and the actin cytoskeleton of hair cell stereocilia and sensory cell microvilli. Cell. Mol. Life Sci. 2006, 63, 2329–2341. [Google Scholar] [CrossRef] [Green Version]
- Sekerkova, G.; Zheng, L.; Mugnaini, E.; Bartles, J.R. Espin actin-cytoskeletal proteins are in rat type I spiral ganglion neurons and include splice-isoforms with a functional nuclear localization signal. J. Comp. Neurol. 2008, 509, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Sekerkova, G.; Loomis, P.A.; Changyaleket, B.; Zheng, L.; Eytan, R.; Chen, B.; Mugnaini, E.; Bartles, J.R. Novel espin actin-bundling proteins are localized to Purkinje cell dendritic spines and bind the Src homology 3 adapter protein insulin receptor substrate p53. J. Neurosci. 2003, 23, 1310–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donaudy, F.; Zheng, L.; Ficarella, R.; Ballana, E.; Carella, M.; Melchionda, S.; Estivill, X.; Bartles, J.R.; Gasparini, P. Espin gene (ESPN) mutations associated with autosomal dominant hearing loss cause defects in microvillar elongation or organisation. J. Med. Genet. 2006, 43, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Sekerkova, G.; Richter, C.P.; Bartles, J.R. Roles of the espin actin-bundling proteins in the morphogenesis and stabilization of hair cell stereocilia revealed in CBA/CaJ congenic jerker mice. PLoS Genet. 2011, 7, e1002032. [Google Scholar] [CrossRef] [Green Version]
- Naz, S.; Griffith, A.J.; Riazuddin, S.; Hampton, L.L.; Battey, J.F., Jr.; Khan, S.N.; Riazuddin, S.; Wilcox, E.R.; Friedman, T.B. Mutations of ESPN cause autosomal recessive deafness and vestibular dysfunction. J. Med. Genet. 2004, 41, 591–595. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Zhang, L.; Tan, F.; Liu, Y.; Chu, C.; Zhu, W.; Wang, Y.; Qi, Z.; Chai, R. Espin distribution as revealed by super-resolution microscopy of stereocilia. Am. J. Transl. Res. 2020, 12, 130–141. [Google Scholar]
- Bartles, J.R.; Zheng, L.; Li, A.; Wierda, A.; Chen, B. Small espin: A third actin-bundling protein and potential forked protein ortholog in brush border microvilli. J. Cell Biol. 1998, 143, 107–119. [Google Scholar] [CrossRef]
- Sekerkova, G.; Zheng, L.; Mugnaini, E.; Bartles, J.R. Differential expression of espin isoforms during epithelial morphogenesis, stereociliogenesis and postnatal maturation in the developing inner ear. Dev. Biol. 2006, 291, 83–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loomis, P.A.; Zheng, L.; Sekerkova, G.; Changyaleket, B.; Mugnaini, E.; Bartles, J.R. Espin cross-links cause the elongation of microvillus-type parallel actin bundles in vivo. J. Cell Biol. 2003, 163, 1045–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grieshaber, S.S.; Lankenau, D.H.; Talbot, T.; Holland, S.; Petersen, N.S. Expression of the 53 kD forked protein rescues F-actin bundle formation and mutant bristle phenotypes in Drosophila. Cell Motil. Cytoskelet. 2001, 50, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, S.; Avenarius, M.R.; Grati, M.; Krey, J.F.; Windsor, A.M.; Sousa, A.D.; Ballesteros, A.; Cui, R.; Millis, B.A.; Salles, F.T.; et al. Stereocilia-staircase spacing is influenced by myosin III motors and their cargos espin-1 and espin-like. Nat. Commun. 2016, 7, 10833. [Google Scholar] [CrossRef] [Green Version]
- Salles, F.T.; Merritt, R.C., Jr.; Manor, U.; Dougherty, G.W.; Sousa, A.D.; Moore, J.E.; Yengo, C.M.; Dosé, A.C.; Kachar, B. Myosin IIIa boosts elongation of stereocilia by transporting espin 1 to the plus ends of actin filaments. Nat. Cell Biol. 2009, 11, 443–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Co, C.; Wong, D.T.; Gierke, S.; Chang, V.; Taunton, J. Mechanism of actin network attachment to moving membranes: Barbed end capture by N-WASP WH2 domains. Cell 2007, 128, 901–913. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Beeler, D.M.; Bartles, J.R. Characterization and regulation of an additional actin-filament-binding site in large isoforms of the stereocilia actin-bundling protein espin. J. Cell Sci. 2014, 127, 1306–1317. [Google Scholar] [CrossRef] [Green Version]
- Loomis, P.A.; Kelly, A.E.; Zheng, L.; Changyaleket, B.; Sekerkova, G.; Mugnaini, E.; Ferreira, A.; Mullins, R.D.; Bartles, J.R. Targeted wild-type and jerker espins reveal a novel, WH2-domain-dependent way to make actin bundles in cells. J. Cell Sci. 2006, 119, 1655–1665. [Google Scholar] [CrossRef] [Green Version]
- Courtemanche, N.; Pollard, T.D. Interaction of profilin with the barbed end of actin filaments. Biochemistry 2013, 52, 6456–6466. [Google Scholar] [CrossRef]
- Morgan, A.A.; Rubenstein, E. Proline: The distribution, frequency, positioning, and common functional roles of proline and polyproline sequences in the human proteome. PLoS ONE 2013, 8, e53785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merritt, R.C.; Manor, U.; Salles, F.T.; Grati, M.; Dose, A.C.; Unrath, W.C.; Quintero, O.A.; Yengo, C.M.; Kachar, B. Myosin IIIB uses an actin-binding motif in its espin-1 cargo to reach the tips of actin protrusions. Curr. Biol. 2012, 22, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Li, J.; Raval, M.H.; Yao, N.; Deng, X.; Lu, Q.; Nie, S.; Feng, W.; Wan, J.; Yengo, C.M.; et al. Myosin III-mediated cross-linking and stimulation of actin bundling activity of Espin. Elife 2016, 5, e12856. [Google Scholar] [CrossRef] [PubMed]
- Bisi, S.; Marchesi, S.; Rizvi, A.; Carra, D.; Beznoussenko, G.V.; Ferrara, I.; Deflorian, G.; Mironov, A.; Bertalot, G.; Pisati, F.; et al. IRSp53 controls plasma membrane shape and polarized transport at the nascent lumen in epithelial tubules. Nat. Commun. 2020, 11, 3516. [Google Scholar] [CrossRef] [PubMed]
- Purdy, K.R.; Bartles, J.R.; Wong, G.C. Structural polymorphism of the actin-espin system: A prototypical system of filaments and linkers in stereocilia. Phys. Rev. Lett. 2007, 98, 058105. [Google Scholar] [CrossRef] [PubMed]
- Kitajiri, S.; Sakamoto, T.; Belyantseva, I.A.; Goodyear, R.J.; Stepanyan, R.; Fujiwara, I.; Bird, J.E.; Riazuddin, S.; Riazuddin, S.; Ahmed, Z.M.; et al. Actin-bundling protein TRIOBP forms resilient rootlets of hair cell stereocilia essential for hearing. Cell 2010, 141, 786–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pangrsic, T.; Singer, J.H.; Koschak, A. Voltage-Gated Calcium Channels: Key Players in Sensory Coding in the Retina and the Inner Ear. Physiol. Rev. 2018, 98, 2063–2096. [Google Scholar] [CrossRef]
- Wang, L.; Zou, J.; Shen, Z.; Song, E.; Yang, J. Whirlin interacts with espin and modulates its actin-regulatory function: An insight into the mechanism of Usher syndrome type II. Hum. Mol. Genet. 2012, 21, 692–710. [Google Scholar] [CrossRef] [Green Version]
- Grega-Larson, N.E.; Crawley, S.W.; Tyska, M.J. Impact of cordon-bleu expression on actin cytoskeleton architecture and dynamics. Cytoskeleton 2016, 73, 670–679. [Google Scholar] [CrossRef] [Green Version]
- Rottner, K.; Faix, J.; Bogdan, S.; Linder, S.; Kerkhoff, E. Actin assembly mechanisms at a glance. J. Cell Sci. 2017, 130, 3427–3435. [Google Scholar] [CrossRef] [Green Version]
- Pellegrin, S.; Mellor, H. Actin stress fibres. J. Cell Sci. 2007, 120, 3491–3499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmann, T.; Dehghani, F. The Cytoskeleton-A Complex Interacting Meshwork. Cells 2019, 8, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revach, O.Y.; Grosheva, I.; Geiger, B. Biomechanical regulation of focal adhesion and invadopodia formation. J. Cell Sci. 2020, 133, jcs244848. [Google Scholar] [CrossRef]
- Tilney, L.G.; Bonder, E.M.; DeRosier, D.J. Actin filaments elongate from their membrane-associated ends. J. Cell Biol. 1981, 90, 485–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Higgs, H.N. Dissecting requirements for auto-inhibition of actin nucleation by the formin, mDia1. J. Biol. Chem. 2005, 280, 6986–6992. [Google Scholar] [CrossRef] [Green Version]
- Haga, R.B.; Ridley, A.J. Rho GTPases: Regulation and roles in cancer cell biology. Small GTPases 2016, 7, 207–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohnert, K.A.; Willet, A.H.; Kovar, D.R.; Gould, K.L. Formin-based control of the actin cytoskeleton during cytokinesis. Biochem. Soc. Trans. 2013, 41, 1750–1754. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Czech, L.; Gerboth, S.; Kojima, S.; Scita, G.; Svitkina, T. Novel roles of formin mDia2 in lamellipodia and filopodia formation in motile cells. PLoS Biol. 2007, 5, e317. [Google Scholar] [CrossRef] [Green Version]
- Barzik, M.; Kotova, T.I.; Higgs, H.N.; Hazelwood, L.; Hanein, D.; Gertler, F.B.; Schafer, D.A. Ena/VASP proteins enhance actin polymerization in the presence of barbed end capping proteins. J. Biol. Chem. 2005, 280, 28653–28662. [Google Scholar] [CrossRef] [Green Version]
- Damiano-Guercio, J.; Kurzawa, L.; Mueller, J.; Dimchev, G.; Schaks, M.; Nemethova, M.; Pokrant, T.; Brühmann, S.; Linkner, J.; Blanchoin, L.; et al. Loss of Ena/VASP interferes with lamellipodium architecture, motility and integrin-dependent adhesion. Elife 2020, 9, e55351. [Google Scholar] [CrossRef]
- Young, L.E.; Heimsath, E.G.; Higgs, H.N. Cell type-dependent mechanisms for formin-mediated assembly of filopodia. Mol. Biol. Cell 2015, 26, 4646–4659. [Google Scholar] [CrossRef]
- Chesarone, M.A.; DuPage, A.G.; Goode, B.L. Unleashing formins to remodel the actin and microtubule cytoskeletons. Nat. Rev. Mol. Cell Biol. 2010, 11, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Sechi, A.S.; Wehland, J. ENA/VASP proteins: Multifunctional regulators of actin cytoskeleton dynamics. Front. Biosci. 2004, 9, 1294–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovar, D.R.; Harris, E.S.; Mahaffy, R.; Higgs, H.N.; Pollard, T.D. Control of the assembly of ATP- and ADP-actin by formins and profilin. Cell 2006, 124, 423–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitsprecher, D.; Kiesewetter, A.K.; Linkner, J.; Vinzenz, M.; Stradal, T.E.; Small, J.V.; Curth, U.; Dickinson, R.B.; Faix, J. Molecular mechanism of Ena/VASP-mediated actin-filament elongation. EMBO J. 2011, 30, 456–467. [Google Scholar] [CrossRef]
- Zigmond, S.H.; Evangelista, M.; Boone, C.; Yang, C.; Dar, A.C.; Sicheri, F.; Forkey, J.; Pring, M. Formin leaky cap allows elongation in the presence of tight capping proteins. Curr. Biol. 2003, 13, 1820–1823. [Google Scholar] [CrossRef] [Green Version]
- Hotulainen, P.; Llano, O.; Smirnov, S.; Tanhuanpää, K.; Faix, J.; Rivera, C.; Lappalainen, P. Defining mechanisms of actin polymerization and depolymerization during dendritic spine morphogenesis. J. Cell Biol. 2009, 185, 323–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vig, A.T.; Foldi, I.; Szikora, S.; Migh, E.; Gombos, R.; Toth, M.A.; Huber, T.; Pinter, R.; Talian, G.C.; Mihaly, J.; et al. The activities of the C-terminal regions of the formin protein disheveled-associated activator of morphogenesis (DAAM) in actin dynamics. J. Biol. Chem. 2017, 292, 13566–13583. [Google Scholar] [CrossRef]
- Guild, G.M.; Connelly, P.S.; Ruggiero, L.; Vranich, K.A.; Tilney, L.G. Long continuous actin bundles in Drosophila bristles are constructed by overlapping short filaments. J. Cell Biol. 2003, 162, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Barr-Gillespie, P.G. Assembly of hair bundles, an amazing problem for cell biology. Mol. Biol. Cell 2015, 26, 2727–2732. [Google Scholar] [CrossRef]
- Manor, U.; Disanza, A.; Grati, M.; Andrade, L.; Lin, H.; Di Fiore, P.P.; Scita, G.; Kachar, B. Regulation of stereocilia length by myosin XVa and whirlin depends on the actin-regulatory protein Eps8. Curr. Biol. 2011, 21, 167–172. [Google Scholar] [CrossRef] [Green Version]
- McGrath, J.; Roy, P.; Perrin, B.J. Stereocilia morphogenesis and maintenance through regulation of actin stability. Semin. Cell Dev. Biol. 2017, 65, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Belyantseva, I.A.; Boger, E.T.; Friedman, T.B. Myosin XVa localizes to the tips of inner ear sensory cell stereocilia and is essential for staircase formation of the hair bundle. Proc. Natl. Acad. Sci. USA 2003, 100, 13958–13963. [Google Scholar] [CrossRef] [Green Version]
- Belyantseva, I.A.; Boger, E.T.; Naz, S.; Frolenkov, G.I.; Sellers, J.R.; Ahmed, Z.M.; Griffith, A.J.; Friedman, T.B. Myosin-XVa is required for tip localization of whirlin and differential elongation of hair-cell stereocilia. Nat. Cell Biol. 2005, 7, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Boëda, B.; El-Amraoui, A.; Bahloul, A.; Goodyear, R.; Daviet, L.; Blanchard, S.; Perfettini, I.; Fath, K.R.; Shorte, S.; Reiners, J.; et al. Myosin VIIa, harmonin and cadherin 23, three Usher I gene products that cooperate to shape the sensory hair cell bundle. EMBO J. 2002, 21, 6689–6699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreland, Z.G.; Bird, J.E. Myosin motors in sensory hair bundle assembly. Curr. Opin. Cell Biol. 2022, 79, 102132. [Google Scholar] [CrossRef] [PubMed]
- Gong, R.; Jiang, F.; Moreland, Z.G.; Reynolds, M.J.; de Los Reyes, S.E.; Gurel, P.; Shams, A.; Heidings, J.B.; Bowl, M.R.; Bird, J.E.; et al. Structural basis for tunable control of actin dynamics by myosin-15 in mechanosensory stereocilia. Sci. Adv. 2022, 8, eabl4733. [Google Scholar] [CrossRef]
- Rich, S.K.; Baskar, R.; Terman, J.R. Propagation of F-actin disassembly via Myosin15-Mical interactions. Sci. Adv. 2021, 7, eabg0147. [Google Scholar] [CrossRef]
- Castaneda, N.; Park, J.; Kang, E.H. Regulation of Actin Bundle Mechanics and Structure by Intracellular Environmental Factors. Front. Phys. 2021, 9, 675885. [Google Scholar] [CrossRef]
- Caberlotto, E.; Michel, V.; de Monvel, J.B.; Petit, C. Coupling of the mechanotransduction machinery and F-actin polymerization in the cochlear hair bundles. Bioarchitecture 2011, 1, 169–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zampini, V.; Ruttiger, L.; Johnson, S.L.; Franz, C.; Furness, D.N.; Waldhaus, J.; Xiong, H.; Hackney, C.M.; Holley, M.C.; Offenhauser, N.; et al. Eps8 regulates hair bundle length and functional maturation of mammalian auditory hair cells. PLoS Biol. 2011, 9, e10010482011. [Google Scholar] [CrossRef] [PubMed]
- Kazmierczak, P.; Sakaguchi, H.; Tokita, J.; Wilson-Kubalek, E.M.; Milligan, R.A.; Muller, U.; Kachar, B. Cadherin 23 and protocadherin 15 interact to form tip-link filaments in sensory hair cells. Nature 2007, 449, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Heckman, C.A.; Ademuyiwa, O.M.; Cayer, M.L. How filopodia respond to calcium in the absence of a calcium-binding structural protein: Non-channel functions of TRP. Cell Commun. Signal. 2022, 20, 130. [Google Scholar] [CrossRef] [PubMed]
- Kovar, D.R.; Pollard, T.D. Insertional assembly of actin filament barbed ends in association with formins produces piconewton forces. Proc. Natl. Acad Sci. USA 2004, 101, 14725–14730. [Google Scholar] [CrossRef] [Green Version]
- Brieher, W.M.; Coughlin, M.; Mitchison, T.J. Fascin-mediated propulsion of Listeria monocytogenes independent of frequent nucleation by the Arp2/3 complex. J. Cell Biol. 2004, 165, 233–242. [Google Scholar] [CrossRef]
- Vignjevic, D.; Peloquin, J.; Borisy, G.G. in vitro assembly of filopodia-like bundles. Methods Enzymol. 2006, 406, 727–739. [Google Scholar] [CrossRef]
- Tilney, L.G.; Connelly, P.; Smith, S.; Guild, G.M. F-actin bundles in Drosophila bristles are assembled from modules composed of short filaments. J. Cell Biol. 1996, 135, 1291–1308. [Google Scholar] [CrossRef] [Green Version]
- Tilney, L.G.; DeRosier, D.J. How to make a curved Drosophila bristle using straight actin bundles. Proc. Natl. Acad. Sci. USA 2005, 102, 18785–18792. [Google Scholar] [CrossRef] [Green Version]
- Perrin, B.J.; Strandjord, D.M.; Narayanan, P.; Henderson, D.M.; Johnson, K.R.; Ervasti, J.M. β-Actin and fascin-2 cooperate to maintain stereocilia length. J. Neurosci. 2013, 33, 8114–8121. [Google Scholar] [CrossRef] [Green Version]
- Roy, P.; Perrin, B.J. The stable actin core of mechanosensory stereocilia features continuous turnover of actin cross-linkers. Mol. Biol. Cell 2018, 29, 1856–1865. [Google Scholar] [CrossRef]
- Avenarius, M.R.; Saylor, K.W.; Lundeberg, M.R.; Wilmarth, P.A.; Shin, J.B.; Spinelli, K.J.; Pagana, J.M.; Andrade, L.; Kachar, B.; Choi, D.; et al. Correlation of actin crosslinker and capper expression levels with stereocilia growth phases. Mol. Cell Proteom. 2014, 13, 606–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Türmer, K.; Orbán, J.; Gróf, P.; Nyitrai, M. FASCIN and alpha-actinin can regulate the conformation of actin filaments. Biochim. Biophys. Acta 2015, 1850, 1855–1861. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; George, S.P. The role of actin bundling proteins in the assembly of filopodia in epithelial cells. Cell Adhes. Migr. 2011, 5, 409–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, J.R.; Homa, K.E.; Morganthaler, A.N.; Brown, R.R.; Suarez, C.; Harker, A.J.; O’Connell, M.E.; Kovar, D.R. Cooperation between tropomyosin and α-actinin inhibits fimbrin association with actin filament networks in fission yeast. Elife 2019, 8, e47279. [Google Scholar] [CrossRef]
- Claessens, M.M.; Semmrich, C.; Ramos, L.; Bausch, A.R. Helical twist controls the thickness of F-actin bundles. Proc. Natl. Acad. Sci. USA 2008, 105, 8819–8822. [Google Scholar] [CrossRef] [Green Version]
- Claessens, M.M.; Bathe, M.; Frey, E.; Bausch, A.R. Actin-binding proteins sensitively mediate F-actin bundle stiffness. Nat. Mater. 2006, 5, 748–753. [Google Scholar] [CrossRef]
- Tilney, L.G.; DeRosier, D.J. Actin filaments, stereocilia, and hair cells of the bird cochlea. IV. How the actin filaments become organized in developing stereocilia and in the cuticular plate. Dev. Biol. 1986, 116, 119–129. [Google Scholar] [CrossRef]
- Stokes, D.L.; DeRosier, D.J. Growth conditions control the size and order of actin bundles in vitro. Biophys. J. 1991, 59, 456–465. [Google Scholar] [CrossRef] [Green Version]
- Haviv, L.; Gov, N.; Ideses, Y.; Bernheim-Groswasser, A. Thickness distribution of actin bundles in vitro. Eur. Biophys. J. 2008, 37, 447–454. [Google Scholar] [CrossRef]
- Winkelman, J.D.; Bilancia, C.G.; Peifer, M.; Kovar, D.R. Ena/VASP Enabled is a highly processive actin polymerase tailored to self-assemble parallel-bundled F-actin networks with Fascin. Proc. Natl. Acad. Sci. USA 2014, 111, 4121–4126. [Google Scholar] [CrossRef] [Green Version]
- Nast-Kolb, T.; Bleicher, P.; Payr, M.; Bausch, A.R. VASP localization to lipid bilayers induces polymerization driven actin bundle formation. Mol. Biol. Cell 2022, 33, mbcE21110577. [Google Scholar] [CrossRef]
- Machaidze, G.; Sokoll, A.; Shimada, A.; Lustig, A.; Mazur, A.; Wittinghofer, A.; Aebi, U.; Mannherz, H.G. Actin filament bundling and different nucleating effects of mouse Diaphanous-related formin FH2 domains on actin/ADF and actin/cofilin complexes. J. Mol. Biol. 2010, 403, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Esue, O.; Harris, E.S.; Higgs, H.N.; Wirtz, D. The filamentous actin cross-linking/bundling activity of mammalian formins. J. Mol. Biol 2008, 384, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Gurel, P.S.; Ge, P.; Grintsevich, E.E.; Shu, R.; Blanchoin, L.; Zhou, Z.H.; Reisler, E.; Higgs, H.N. INF2-mediated severing through actin filament encirclement and disruption. Curr. Biol. 2014, 24, 156–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, E.S.; Rouiller, I.; Hanein, D.; Higgs, H.N. Mechanistic differences in actin bundling activity of two mammalian formins, FRL1 and mDia2. J. Biol. Chem. 2006, 281, 14383–14392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Tanoury, Z.; Schaffner-Reckinger, E.; Halavatyi, A.; Hoffmann, C.; Moes, M.; Hadzic, E.; Catillon, M.; Yatskou, M.; Friederich, E. Quantitative kinetic study of the actin-bundling protein L-plastin and of its impact on actin turn-over. PLoS ONE 2010, 5, e9210. [Google Scholar] [CrossRef] [Green Version]
- Karpova, T.S.; Tatchell, K.; Cooper, J.A. Actin filaments in yeast are unstable in the absence of capping protein or fimbrin. J. Cell Biol. 1995, 131, 1483–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reymann, A.C.; Boujemaa-Paterski, R.; Martiel, J.L.; Guerin, C.; Cao, W.; Chin, H.F.; de la Cruz, E.M.; Thery, M.; Blanchoin, L. Actin network architecture can determine myosin motor activity. Science 2012, 336, 1310–1314. [Google Scholar] [CrossRef] [Green Version]
- Houdusse, A.; Titus, M.A. The many roles of myosins in filopodia, microvilli and stereocilia. Curr. Biol. 2021, 31, r586–r602. [Google Scholar] [CrossRef] [PubMed]
- Wubshet, N.H.; Bashirzadeh, Y.; Liu, A.P. Fascin-induced actin protrusions are suppressed by dendritic networks in giant unilamellar vesicles. Mol. Biol. Cell 2021, 32, 1634–1640. [Google Scholar] [CrossRef]
- Frémont, S.; Hammich, H.; Bai, J.; Wioland, H.; Klinkert, K.; Rocancourt, M.; Kikuti, C.; Stroebel, D.; Romet-Lemonne, G.; Pylypenko, O.; et al. Oxidation of F-actin controls the terminal steps of cytokinesis. Nat. Commun. 2017, 8, 14528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, I.; Vavylonis, D.; Pollard, T.D. Polymerization kinetics of ADP- and ADP-Pi-actin determined by fluorescence microscopy. Proc. Natl. Acad. Sci. USA 2007, 104, 8827–8832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallavarapu, A.; Mitchison, T. Regulated actin cytoskeleton assembly at filopodium tips controls their extension and retraction. J. Cell Biol. 1999, 146, 1097–1106. [Google Scholar] [CrossRef]
- Schneider, M.E.; Belyantseva, I.A.; Azevedo, R.B.; Kachar, B. Rapid renewal of auditory hair bundles. Nature 2002, 418, 837–838. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, P.; Chatterton, P.; Ikeda, A.; Ikeda, S.; Corey, D.P.; Ervasti, J.M.; Perrin, B.J. Length regulation of mechanosensitive stereocilia depends on very slow actin dynamics and filament-severing proteins. Nat. Commun. 2015, 6, 6855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meenderink, L.M.; Gaeta, I.M.; Postema, M.M.; Cencer, C.S.; Chinowsky, C.R.; Krystofiak, E.S.; Millis, B.A.; Tyska, M.J. Actin Dynamics Drive Microvillar Motility and Clustering during Brush Border Assembly. Dev. Cell 2019, 50, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.S.; Piazza, V.; Perrin, B.J.; Rzadzinska, A.K.; Poczatek, J.C.; Wang, M.; Prosser, H.M.; Ervasti, J.M.; Corey, D.P.; Lechene, C.P. Multi-isotope imaging mass spectrometry reveals slow protein turnover in hair-cell stereocilia. Nature 2012, 481, 520–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, M.C.; Barzik, M.; Bird, J.E.; Zhang, D.S.; Lechene, C.P.; Corey, D.P.; Cunningham, L.L.; Friedman, T.B. Live-cell imaging of actin dynamics reveals mechanisms of stereocilia length regulation in the inner ear. Nat. Commun. 2015, 6, 6873. [Google Scholar] [CrossRef] [Green Version]
- Iwasa, J.H.; Mullins, R.D. Spatial and temporal relationships between actin-filament nucleation, capping, and disassembly. Curr. Biol. 2007, 17, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Loisel, T.P.; Boujemaa, R.; Pantaloni, D.; Carlier, M.F. Reconstitution of actin-based motility of Listeria and Shigella using pure proteins. Nature 1999, 401, 613–616. [Google Scholar] [CrossRef]
- Bugyi, B.; Carlier, M.F. Control of actin filament treadmilling in cell motility. Annu. Rev. Biophys. 2010, 39, 449–470. [Google Scholar] [CrossRef]
- Bernstein, B.W.; Bamburg, J.R. ADF/cofilin: A functional node in cell biology. Trends Cell Biol. 2010, 20, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Andrianantoandro, E.; Pollard, T.D. Mechanism of actin filament turnover by severing and nucleation at different concentrations of ADF/cofilin. Mol. Cell 2006, 24, 13–23. [Google Scholar] [CrossRef]
- Bobkov, A.A.; Muhlrad, A.; Shvetsov, A.; Benchaar, S.; Scoville, D.; Almo, S.C.; Reisler, E. Cofilin (ADF) affects lateral contacts in F-actin. J. Mol. Biol. 2004, 337, 93–104. [Google Scholar] [CrossRef]
- Chin, S.M.; Jansen, S.; Goode, B.L. TIRF microscopy analysis of human Cof1, Cof2, and ADF effects on actin filament severing and turnover. J. Mol. Biol. 2016, 428, 1604–1616. [Google Scholar] [CrossRef] [Green Version]
- Suarez, C.; Roland, J.; Boujemaa-Paterski, R.; Kang, H.; McCullough, B.R.; Reymann, A.C.; Guerin, C.; Martiel, J.L.; De La Cruz, E.M.; Blanchoin, L. Cofilin tunes the nucleotide state of actin filaments and severs at bare and decorated segment boundaries. Curr. Biol. 2011, 21, 862–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wioland, H.; Guichard, B.; Senju, Y.; Myram, S.; Lappalainen, P.; Jégou, A.; Romet-Lemonne, G. ADF/Cofilin Accelerates Actin Dynamics by Severing Filaments and Promoting Their Depolymerization at Both Ends. Curr. Biol. 2017, 27, 1956–1967.e1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wioland, H.; Jegou, A.; Romet-Lemonne, G. Quantitative Variations with pH of Actin Depolymerizing Factor/Cofilin’s Multiple Actions on Actin Filaments. Biochemistry 2019, 58, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Kotila, T.; Wioland, H.; Enkavi, G.; Kogan, K.; Vattulainen, I.; Jégou, A.; Romet-Lemonne, G.; Lappalainen, P. Mechanism of synergistic actin filament pointed end depolymerization by cyclase-associated protein and cofilin. Nat. Commun. 2019, 10, 5320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiuchi, T.; Ohashi, K.; Kurita, S.; Mizuno, K. Cofilin promotes stimulus-induced lamellipodium formation by generating an abundant supply of actin monomers. J. Cell Biol. 2007, 177, 465–476. [Google Scholar] [CrossRef] [Green Version]
- Carlier, M.F.; Laurent, V.; Santolini, J.; Melki, R.; Didry, D.; Xia, G.X.; Hong, Y.; Chua, N.H.; Pantaloni, D. Actin depolymerizing factor (ADF/cofilin) enhances the rate of filament turnover: Implication in actin-based motility. J. Cell Biol. 1997, 136, 1307–1322. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Cordero, J.J.; Magalhaes, M.A.; Eddy, R.J.; Hodgson, L.; Condeelis, J. Functions of cofilin in cell locomotion and invasion. Nat. Rev. Mol. Cell Biol. 2013, 14, 405–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Bernstein, B.W.; Bamburg, J.R. Regulating actin-filament dynamics in vivo. Trends Biochem. Sci. 2000, 25, 19–23. [Google Scholar] [CrossRef]
- Faix, J.; Breitsprecher, D.; Stradal, T.E.; Rottner, K. Filopodia: Complex models for simple rods. Int. J. Biochem. Cell Biol. 2009, 41, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Bretscher, A.; Weber, K. Localization of actin and microfilament-associated proteins in the microvilli and terminal web of the intestinal brush border by immunofluorescence microscopy. J. Cell Biol. 1978, 79, 839–845. [Google Scholar] [CrossRef]
- Oser, M.; Condeelis, J. The cofilin activity cycle in lamellipodia and invadopodia. J. Cell Biochem. 2009, 108, 1252–1262. [Google Scholar] [CrossRef] [Green Version]
- Vardouli, L.; Moustakas, A.; Stournaras, C. LIM-kinase 2 and cofilin phosphorylation mediate actin cytoskeleton reorganization induced by transforming growth factor-beta. J. Biol. Chem. 2005, 280, 11448–11457. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.Y.; DerMardirossian, C.; Bokoch, G.M. Cofilin phosphatases and regulation of actin dynamics. Curr. Opin. Cell Biol. 2006, 18, 26–31. [Google Scholar] [CrossRef]
- Zhao, H.; Hakala, M.; Lappalainen, P. ADF/cofilin binds phosphoinositides in a multivalent manner to act as a PIP[2]-density sensor. Biophys. J. 2010, 98, 2327–2336. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Courtemanche, N.; Pollard, T.D. Aip1 promotes actin filament severing by cofilin and regulates constriction of the cytokinetic contractile ring. J. Biol. Chem. 2015, 290, 2289–2300. [Google Scholar] [CrossRef] [Green Version]
- Brieher, W.M.; Kueh, H.Y.; Ballif, B.A.; Mitchison, T.J. Rapid actin monomer–insensitive depolymerization of Listeria actin comet tails by cofilin, coronin, and Aip1. J. Cell Biol. 2006, 175, 315–324. [Google Scholar] [CrossRef]
- Purde, V.; Busch, F.; Kudryashova, E.; Wysocki, V.H.; Kudryashov, D.S. Oligomerization Affects the Ability of Human Cyclase-Associated Proteins 1 and 2 to Promote Actin Severing by Cofilins. Int. J. Mol. Sci. 2019, 20, 5647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, A.W.; Belyantseva, I.A.; Hsu, P.D.; Friedman, T.B.; Heller, S. Twinfilin 2 regulates actin filament lengths in cochlear stereocilia. J. Neurosci. 2009, 29, 15083–15088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shekhar, S.; Hoeprich, G.J.; Gelles, J.; Goode, B.L. Twinfilin bypasses assembly conditions and actin filament aging to drive barbed end depolymerization. J. Cell Biol. 2021, 220, e202006022. [Google Scholar] [CrossRef]
- Sinnar, S.A.; Antoku, S.; Saffin, J.M.; Cooper, J.A.; Halpain, S. Capping protein is essential for cell migration in vivo and for filopodial morphology and dynamics. Mol. Biol. Cell 2014, 25, 2152–2160. [Google Scholar] [CrossRef]
- Zwolak, A.; Fujiwara, I.; Hammer, J.A., 3rd; Tjandra, N. Structural basis for capping protein sequestration by myotrophin (V-1]. J. Biol. Chem. 2010, 285, 25767–25781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, S.; Minakata, S.; Koike, R.; Kawahata, I.; Narita, A.; Kitazawa, M.; Ota, M.; Yamakuni, T.; Maeda, Y.; Nitanai, Y.; et al. Two distinct mechanisms for actin capping protein regulation—Steric and allosteric inhibition. PLoS Biol. 2010, 8, e10004162010. [Google Scholar] [CrossRef] [Green Version]
- Jung, G.; Alexander, C.J.; Wu, X.S.; Piszczek, G.; Chen, B.C.; Betzig, E.; Hammer, J.A. V-1 regulates capping protein activity in vivo. Proc. Natl. Acad. Sci. USA 2016, 113, E6610–E6619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakala, M.; Wioland, H.; Tolonen, M.; Kotila, T.; Jegou, A.; Romet-Lemonne, G.; Lappalainen, P. Twinfilin uncaps filament barbed ends to promote turnover of lamellipodial actin networks. Nat. Cell Biol. 2021, 23, 147–159. [Google Scholar] [CrossRef]
- Johnston, A.B.; Collins, A.; Goode, B.L. High-speed depolymerization at actin filament ends jointly catalysed by Twinfilin and Srv2/CAP. Nat. Cell Biol. 2015, 17, 1504–1511. [Google Scholar] [CrossRef] [Green Version]
- Avenarius, M.R.; Krey, J.F.; Dumont, R.A.; Morgan, C.P.; Benson, C.B.; Vijayakumar, S.; Cunningham, C.L.; Scheffer, D.I.; Corey, D.P.; Muller, U.; et al. Heterodimeric capping protein is required for stereocilia length and width regulation. J. Cell Biol. 2017, 216, 3861–3881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vartiainen, M.K.; Sarkkinen, E.M.; Matilainen, T.; Salminen, M.; Lappalainen, P. Mammals have two twinfilin isoforms whose subcellular localizations and tissue distributions are differentially regulated. J. Biol Chem. 2003, 278, 34347–34355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goode, B.L.; Drubin, D.G.; Lappalainen, P. Regulation of the cortical actin cytoskeleton in budding yeast by twinfilin, a ubiquitous actin monomer-sequestering protein. J. Cell Biol. 1998, 142, 723–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vartiainen, M.; Ojala, P.J.; Auvinen, P.; Peranen, J.; Lappalainen, P. Mouse A6/twinfilin is an actin monomer-binding protein that localizes to the regions of rapid actin dynamics. Mol. Cell Biol. 2000, 20, 1772–1783. [Google Scholar] [CrossRef] [Green Version]
- Nag, S.; Ma, Q.; Wang, H.; Chumnarnsilpa, S.; Lee, W.L.; Larsson, M.; Kannan, B.; Hernandez-Valladares, M.; Burtnick, L.D.; Robinson, R.C. Ca2+ binding by domain 2 plays a critical role in the activation and stabilization of gelsolin. Proc. Natl. Acad. Sci. USA 2009, 106, 13713–13718. [Google Scholar] [CrossRef] [Green Version]
- Nag, S.; Larsson, M.; Robinson, R.C.; Burtnick, L.D. Gelsolin: The tail of a molecular gymnast. Cytoskeleton 2013, 70, 360–384. [Google Scholar] [CrossRef] [PubMed]
- Silacci, P.; Mazzolai, L.; Gauci, C.; Stergiopulos, N.; Yin, H.L.; Hayoz, D. Gelsolin superfamily proteins: Key regulators of cellular functions. Cell Mol. Life Sci. 2004, 61, 2614–2623. [Google Scholar] [CrossRef] [Green Version]
- Mburu, P.; Romero, M.R.; Hilton, H.; Parker, A.; Townsend, S.; Kikkawa, Y.; Brown, S.D. Gelsolin plays a role in the actin polymerization complex of hair cell stereocilia. PLoS ONE 2010, 5, e11627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudryashova, E.; Ankita; Ulrichs, H.; Shekhar, S.; Kudryashov, D.S. Pointed-end processive elongation of actin filaments by Vibrio effectors VopF and VopL. Sci. Adv. 2022, 8, eadc9239. [Google Scholar] [CrossRef]
- Jégou, A.; Niedermayer, T.; Orbán, J.; Didry, D.; Lipowsky, R.; Carlier, M.F.; Romet-Lemonne, G. Individual actin filaments in a microfluidic flow reveal the mechanism of ATP hydrolysis and give insight into the properties of profilin. PLoS Biol. 2011, 9, e1001161. [Google Scholar] [CrossRef] [Green Version]
- Pernier, J.; Shekhar, S.; Jegou, A.; Guichard, B.; Carlier, M.F. Profilin Interaction with Actin Filament Barbed End Controls Dynamic Instability, Capping, Branching, and Motility. Dev. Cell 2016, 36, 201–214. [Google Scholar] [CrossRef] [Green Version]
- Funk, J.; Merino, F.; Venkova, L.; Heydenreich, L.; Kierfeld, J.; Vargas, P.; Raunser, S.; Piel, M.; Bieling, P. Profilin and formin constitute a pacemaker system for robust actin filament growth. Elife 2019, 8, e50963. [Google Scholar] [CrossRef] [PubMed]
- Kotila, T.; Kogan, K.; Enkavi, G.; Guo, S.; Vattulainen, I.; Goode, B.L.; Lappalainen, P. Structural basis of actin monomer re-charging by cyclase-associated protein. Nat. Commun. 2018, 9, 1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terman, J.R.; Mao, T.; Pasterkamp, R.J.; Yu, H.H.; Kolodkin, A.L. MICALs, a family of conserved flavoprotein oxidoreductases, function in plexin-mediated axonal repulsion. Cell 2002, 109, 887–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Nakamoto, T.; Ogawa, S.; Seo, S.; Matsumura, T.; Tachibana, K.; Morimoto, C.; Hirai, H. MICAL, a novel CasL interacting molecule, associates with vimentin. J. Biol. Chem. 2002, 277, 14933–14941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, R.J.; Spaeth, C.S.; Yesilyurt, H.G.; Terman, J.R. SelR reverses Mical-mediated oxidation of actin to regulate F-actin dynamics. Nat. Cell Biol. 2013, 15, 1445–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanoni, M.A.; Vitali, T.; Zucchini, D. MICAL, the flavoenzyme participating in cytoskeleton dynamics. Int. J. Mol. Sci. 2013, 14, 6920–6959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frémont, S.; Romet-Lemonne, G.; Houdusse, A.; Echard, A. Emerging roles of MICAL family proteins—From actin oxidation to membrane trafficking during cytokinesis. J. Cell Sci. 2017, 130, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Hung, R.J.; Terman, J.R. Extracellular inhibitors, repellents, and semaphorin/plexin/MICAL-mediated actin filament disassembly. Cytoskeleton 2011, 68, 415–433. [Google Scholar] [CrossRef] [Green Version]
- Hung, R.J.; Yazdani, U.; Yoon, J.; Wu, H.; Yang, T.; Gupta, N.; Huang, Z.; van Berkel, W.J.; Terman, J.R. Mical links semaphorins to F-actin disassembly. Nature 2010, 463, 823–827. [Google Scholar] [CrossRef] [Green Version]
- Alto, L.T.; Terman, J.R. MICALs. Curr. Biol. 2018, 28, R538–R5412018. [Google Scholar] [CrossRef] [Green Version]
- Grintsevich, E.E.; Ge, P.; Sawaya, M.R.; Yesilyurt, H.G.; Terman, J.R.; Zhou, Z.H.; Reisler, E. Catastrophic disassembly of actin filaments via Mical-mediated oxidation. Nat. Commun. 2017, 8, 2183. [Google Scholar] [CrossRef] [Green Version]
- Grintsevich, E.E.; Yesilyurt, H.G.; Rich, S.K.; Hung, R.J.; Terman, J.R.; Reisler, E. F-actin dismantling through a redox-driven synergy between Mical and cofilin. Nat. Cell Biol. 2016, 18, 876–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellers, J.R. Myosins: A diverse superfamily. Biochim. Biophys. Acta 2000, 1496, 3–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomakin, A.J.; Lee, K.C.; Han, S.J.; Bui, D.A.; Davidson, M.; Mogilner, A.; Danuser, G. Competition for actin between two distinct F-actin networks defines a bistable switch for cell polarization. Nat. Cell Biol. 2015, 17, 1435–1445. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, N.A.; Burnette, D.T.; Forscher, P. Myosin II functions in actin-bundle turnover in neuronal growth cones. Nat. Cell Biol. 2006, 8, 215–226. [Google Scholar] [CrossRef]
- Vemula, V.; Huber, T.; Usaj, M.; Bugyi, B.; Mansson, A. Myosin and gelsolin cooperate in actin filament severing and actomyosin motor activity. J. Biol. Chem. 2021, 296, 100181. [Google Scholar] [CrossRef]
- Janco, M.; Dedova, I.; Bryce, N.S.; Hardeman, E.C.; Gunning, P.W. Visualizing the in vitro assembly of tropomyosin/actin filaments using TIRF microscopy. Biophys. Rev. 2020, 12, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Gray, K.T.; Kostyukova, A.S.; Fath, T. Actin regulation by tropomodulin and tropomyosin in neuronal morphogenesis and function. Mol. Cell Neurosci. 2017, 84, 48–57. [Google Scholar] [CrossRef]
- Furutani, Y.; Yoshihara, Y. Proteomic Analysis of Dendritic Filopodia-Rich Fraction Isolated by Telencephalin and Vitronectin Interaction. Front. Synaptic Neurosci. 2018, 10, 27. [Google Scholar] [CrossRef]
- Qin, W.; Cho, K.F.; Cavanagh, P.E.; Ting, A.Y. Deciphering molecular interactions by proximity labeling. Nat. Methods 2021, 18, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Jasnin, M.; Hervy, J.; Balor, S.; Bouissou, A.; Proag, A.; Voituriez, R.; Schneider, J.; Mangeat, T.; Maridonneau-Parini, I.; Baumeister, W.; et al. Elasticity of podosome actin networks produces nanonewton protrusive forces. Nat. Commun. 2022, 13, 3842. [Google Scholar] [CrossRef]
- Prakash, K.; Diederich, B.; Heintzmann, R.; Schermelleh, L. Super-resolution microscopy: A brief history and new avenues. Philos. Trans. R. Soc. A 2022, 380, 20210110. [Google Scholar] [CrossRef] [PubMed]
- Wassie, A.T.; Zhao, Y.; Boyden, E.S. Expansion microscopy: Principles and uses in biological research. Nat. Methods 2019, 16, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Kraus, J.; Russell, R.W.; Kudryashova, E.; Xu, C.; Katyal, N.; Perilla, J.R.; Kudryashov, D.S.; Polenova, T. Magic angle spinning NMR structure of human cofilin-2 assembled on actin filaments reveals isoform-specific conformation and binding mode. Nat. Commun. 2022, 13, 2114. [Google Scholar] [CrossRef]
- Yehl, J.; Kudryashova, E.; Reisler, E.; Kudryashov, D.; Polenova, T. Structural Analysis of Human Cofilin 2/Filamentous Actin Assemblies: Atomic-Resolution Insights from Magic Angle Spinning NMR Spectroscopy. Sci. Rep. 2017, 7, 44506. [Google Scholar] [CrossRef] [Green Version]
- Warnet, X.L.; Arnold, A.A.; Marcotte, I.; Warschawski, D.E. In-Cell Solid-State NMR: An Emerging Technique for the Study of Biological Membranes. Biophys. J. 2015, 109, 2461–2466. [Google Scholar] [CrossRef] [Green Version]





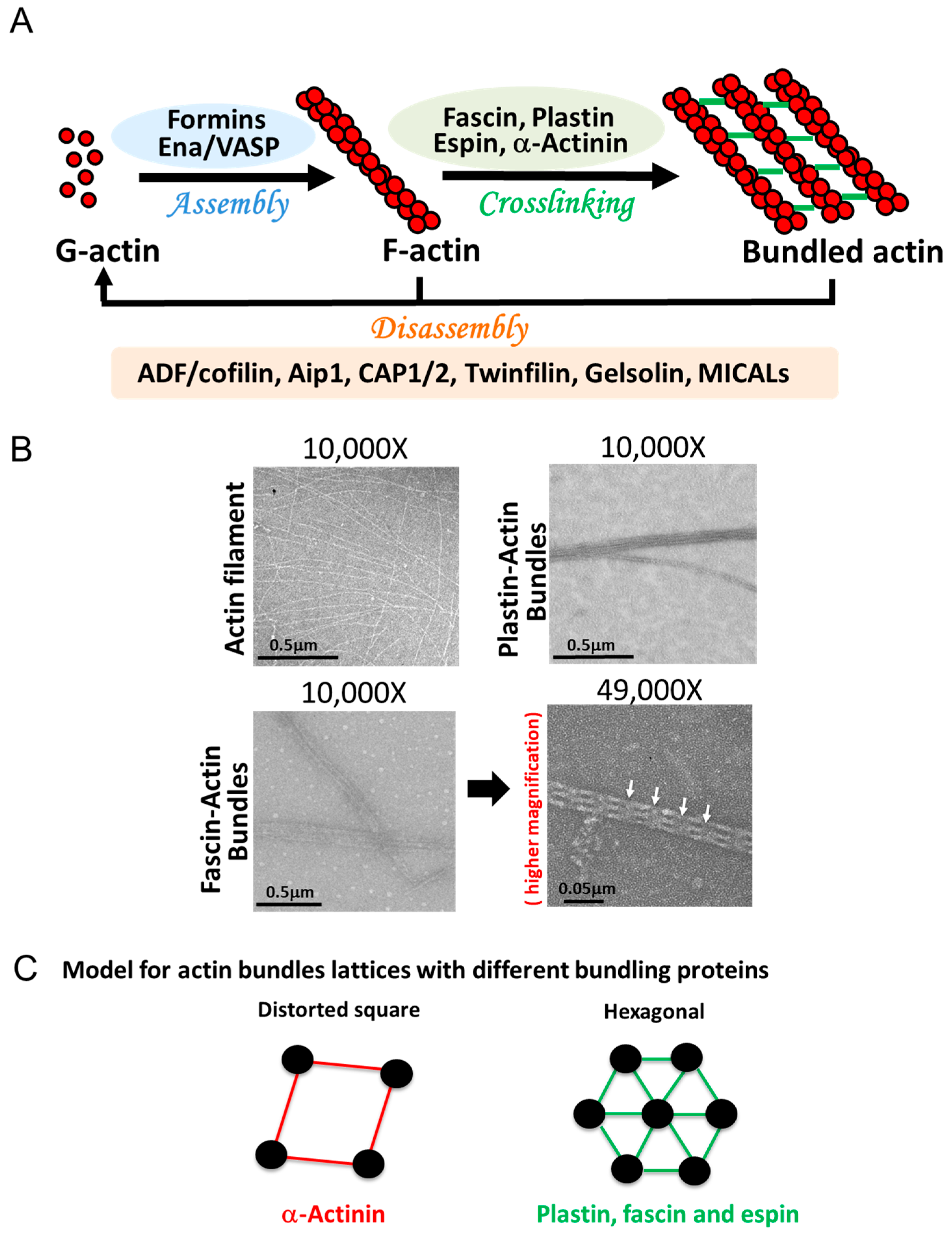{kind=link}
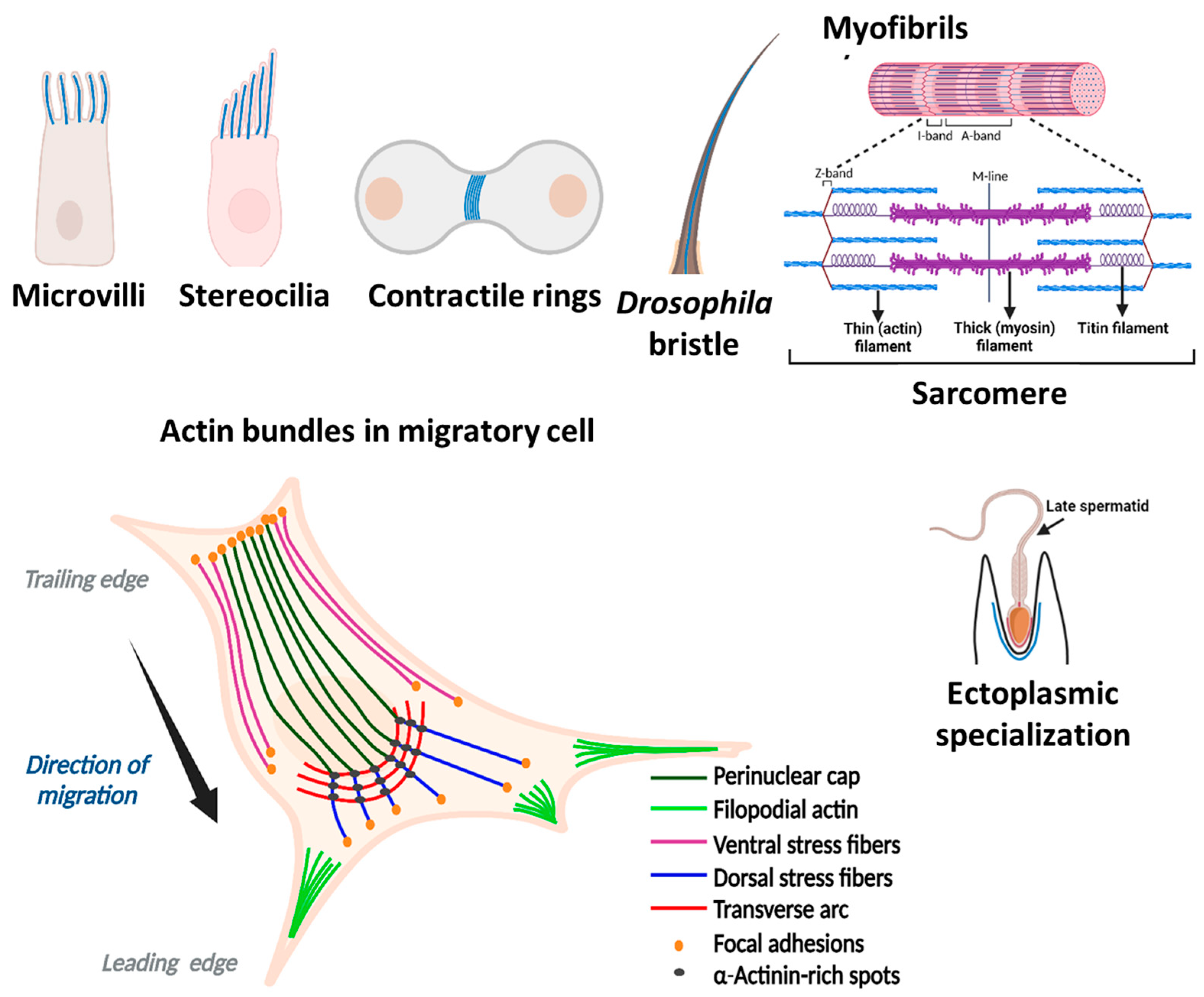{kind=link}
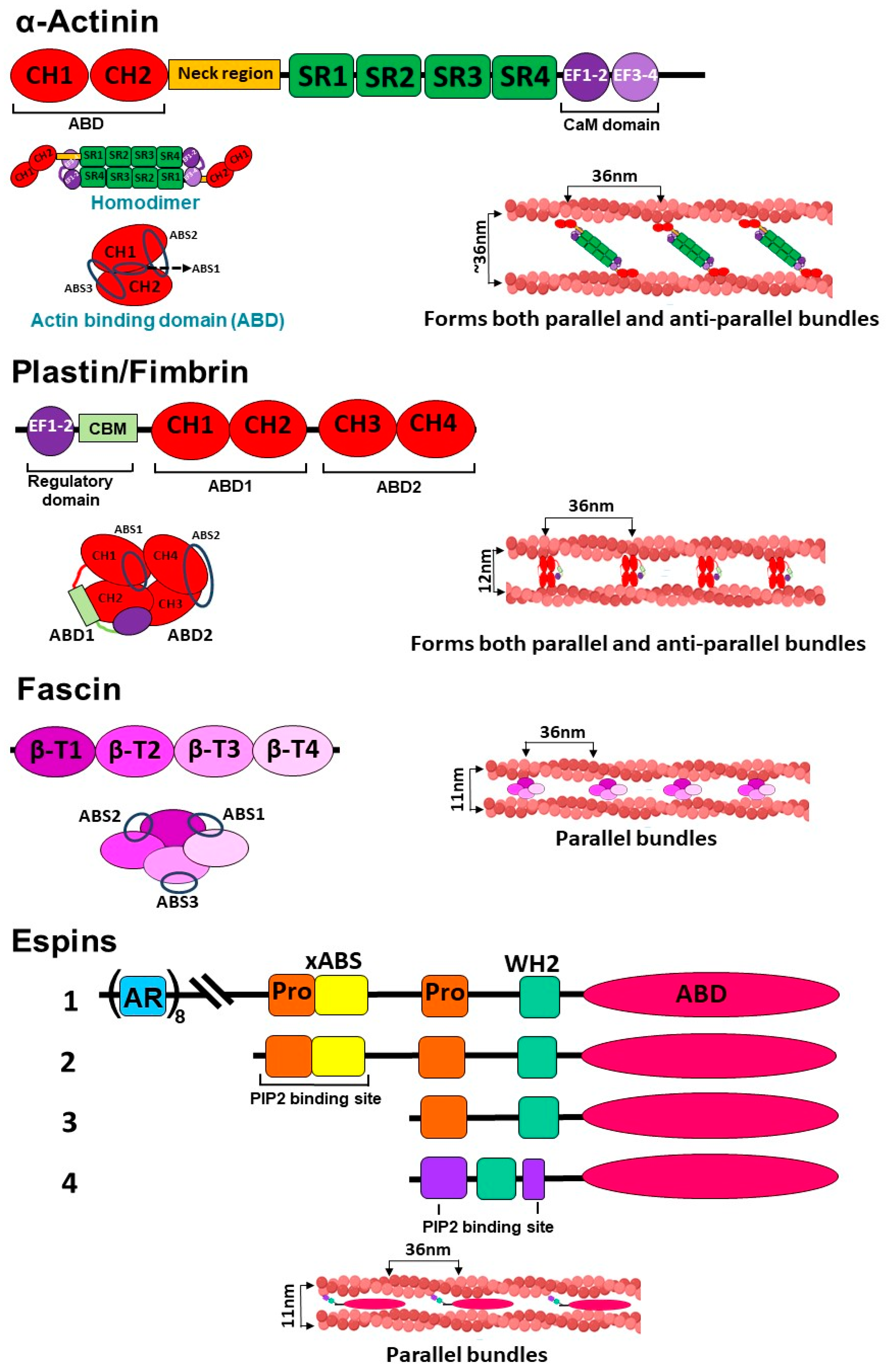{kind=link}
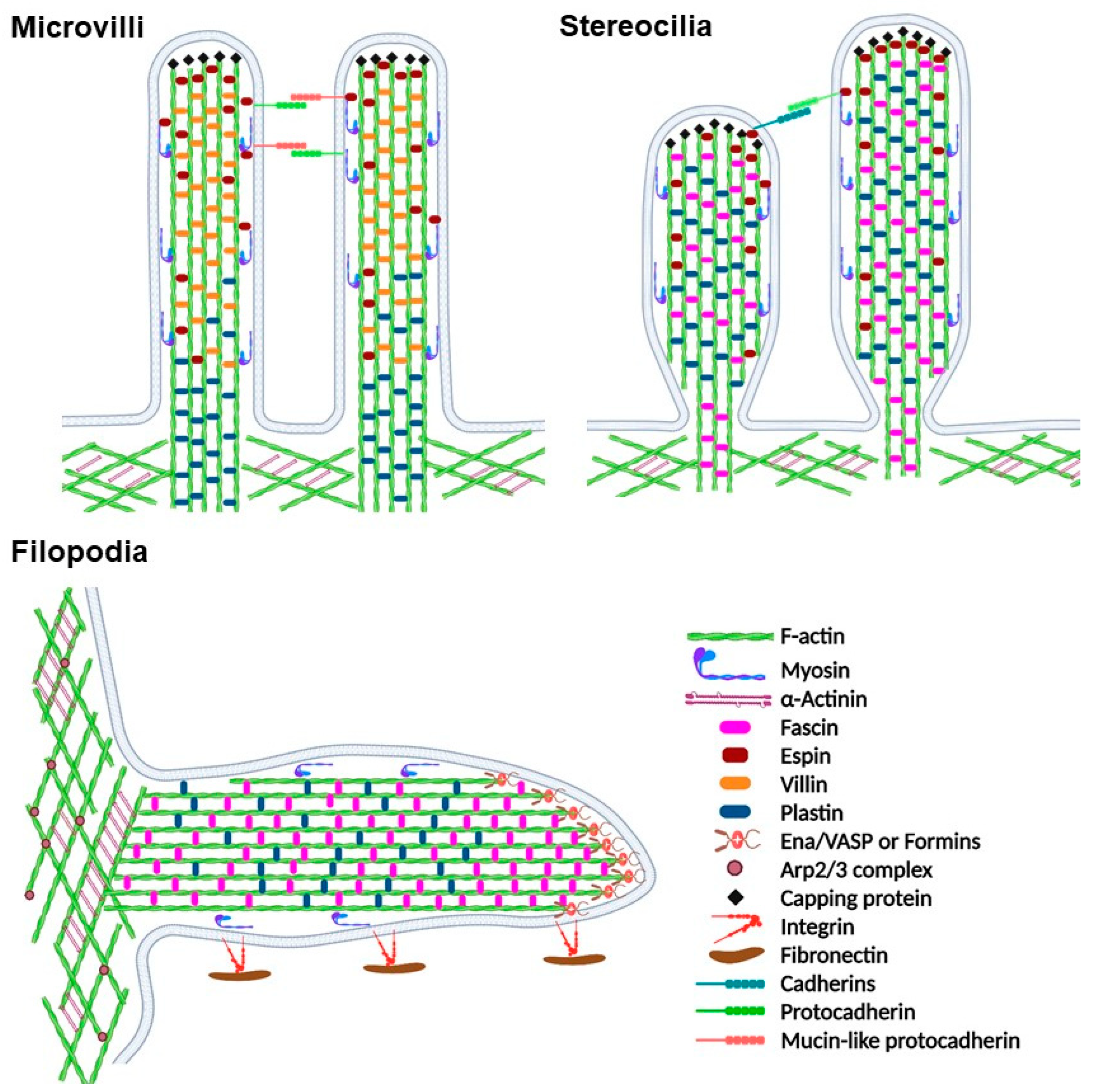{kind=link}
| Location | Microvilli | Stereocilia | Filopodia | Stress fibers | Bristles (drosophila) |
|---|---|---|---|---|---|
| Cell type | Most cells (frequent in epithelial cells) | Auditory and vestibular sensory cells | Motile cells | Most cells (prominent in fibroblasts, smooth muscle, and endothelial cells) | Sensory organ precursor cells |
| Function | Increase apical surface area for absorption | Mechano-electrical signaling | Sensory and guiding | Contraction and adhesions | Mechanosensing |
| Length | 100 nm to 2 µm | 1.5–15 µm | ≤10 µm | ≥2 µm | Macrochaetes: 250–300 μm, Microchaetes: 70 μm (non-continuous 1–5 µm units) |
| Diameter | 50–100 nm | ~200 nm | 20–200 nm | Varies from cell to cell | Varies |
| Number of actin filaments | 30–40 | ~400–3000 | 10–30 | 10–30 | 7–18 bundles with hundreds of filaments |
| Actin filament organization | Parallel (unipolar) | Parallel (unipolar) | Parallel (unipolar) | Mixed (bipolar) | Parallel |
| Bundling proteins | Espin, plastin, villin | Fascin, espin, plastin | Fascin, α-actinin, plastin, espin | α-Actinin, fascin filamin | singed (fascin), forked (espin) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajan, S.; Kudryashov, D.S.; Reisler, E. Actin Bundles Dynamics and Architecture. Biomolecules 2023, 13, 450. https://doi.org/10.3390/biom13030450
Rajan S, Kudryashov DS, Reisler E. Actin Bundles Dynamics and Architecture. Biomolecules. 2023; 13(3):450. https://doi.org/10.3390/biom13030450
Chicago/Turabian StyleRajan, Sudeepa, Dmitri S. Kudryashov, and Emil Reisler. 2023. "Actin Bundles Dynamics and Architecture" Biomolecules 13, no. 3: 450. https://doi.org/10.3390/biom13030450
APA StyleRajan, S., Kudryashov, D. S., & Reisler, E. (2023). Actin Bundles Dynamics and Architecture. Biomolecules, 13(3), 450. https://doi.org/10.3390/biom13030450