


Molecular Basis for Actin Polymerization Kinetics Modulated by Solution Crowding

 and
and
Abstract
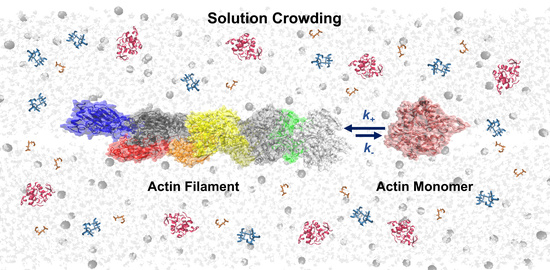
:
1. Introduction
2. Materials and Methods
2.1. Protein Purification and Sample Preparations
2.2. Flow Cell Preparation
2.3. TIRF Microscopy Imaging and Data Analysis
2.4. Pyrene Fluorescence Spectroscopy
2.5. Molecular Dynamics (MD) Simulations
2.6. Actin Subunit Diffusion Analysis from MD Simulations
2.7. Polar Solvation Energy Calculation from MD Simulations
3. Results and Discussion
3.1. Crowding Modulates Actin Filament Assembly Kinetics
3.2. Crowding Modulates the Diffusion of Actin Subunits near Filament Barbed Ends
3.3. Crowding Affects Filament Electrostatic Energy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fletcher, D.A.; Mullins, D. Cell mechanics and the cytoskeleton. Nature 2010, 463, 485–492. [Google Scholar] [CrossRef]
- Blanchoin, L.; Boujemaa-Paterski, R.; Sykes, C.; Plastino, J. Actin dynamics, architecture, and mechanics in cell motility. Physiol. Rev. 2014, 94, 235–263. [Google Scholar] [CrossRef]
- Pollard, T.D. Actin and Actin-Binding Proteins. Cold Spring Harb. Perspect. Biol. 2016, 8, a018226. [Google Scholar] [CrossRef]
- Castaneda, N.; Park, J.; Kang, E.H. Regulation of Actin Bundle Mechanics and Structure by Intracellular Environmental Factors. Front. Phys. 2021, 9, 675885. [Google Scholar] [CrossRef]
- Ellis, R.J. Macromolecular crowding: Obvious but underappreciated. Trends Biochem. Sci. 2001, 26, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.J.; Minton, A.P. Join the crowd. Nature 2003, 425, 27–28. [Google Scholar] [CrossRef] [PubMed]
- Rivas, G.; Minton, A.P. Macromolecular Crowding In Vitro, In Vivo, and In Between. Trends Biochem. Sci. 2016, 41, 970–981. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, S.B.; Minton, A.P. Macromolecular Crowding: Biochemical, Biophysical, and Physiological Consequences. Annu. Rev. Biophys. Biomol. Struct. 1993, 22, 27–65. [Google Scholar] [CrossRef]
- Minton, A.P. Implications of macromolecular crowding for protein assembly. Curr. Opin. Struct. Biol. 2000, 10, 34–39. [Google Scholar] [CrossRef]
- Kuznetsova, I.M.; Turoverov, K.K.; Uversky, V.N. What macromolecular crowding can do to a protein. Int. J. Mol. Sci. 2014, 15, 23090–23140. [Google Scholar] [CrossRef]
- Shahid, S.; Hassan, M.I.; Islam, A.; Ahmad, F. Size-dependent studies of macromolecular crowding on the thermodynamic stability, structure and functional activity of proteins: In vitro and in silico approaches. Biochim. Biophys. Acta (BBA) - Gen. Subj. 2017, 1861, 178–197. [Google Scholar] [CrossRef]
- Kim, J.S.; Yethiraj, A. Effect of Macromolecular Crowding on Reaction Rates: A Computational and Theoretical Study. Biophys. J. 2009, 96, 1333–1340. [Google Scholar] [CrossRef]
- Rivas, G.; Minton, A.P. Influence of Nonspecific Interactions on Protein Associations: Implications for Biochemistry In Vivo. Annu. Rev. Biochem. 2022, 91, 321–351. [Google Scholar] [CrossRef]
- Drenckhahn, D.; Pollard, T.D. Elongation of actin filaments is a diffusion-limited reaction at the barbed end and is accelerated by inert macromolecules. J. Biol. Chem. 1986, 261, 12754–12758. [Google Scholar] [CrossRef]
- Gao, M.; Winter, R. Kinetic Insights into the Elongation Reaction of Actin Filaments as a Function of Temperature, Pressure, and Macromolecular Crowding. ChemPhysChem 2015, 16, 3681–3686. [Google Scholar] [CrossRef] [PubMed]
- Lindner, R.A.; Ralston, G.B. Macromolecular crowding: Effects on actin polymerisation. Biophys. Chem. 1997, 66, 57–66. [Google Scholar] [CrossRef]
- Rashid, R.; Chee, S.M.; Raghunath, M.; Wohland, T. Macromolecular crowding gives rise to microviscosity, anomalous diffusion and accelerated actin polymerization. Phys. Biol. 2015, 12, 034001. [Google Scholar] [CrossRef]
- Rosin, C.; Schummel, P.H.; Winter, R. Cosolvent and crowding effects on the polymerization kinetics of actin. Phys. Chem. Chem. Phys. 2015, 17, 8330–8337. [Google Scholar] [CrossRef]
- Schummel, P.H.; Haag, A.; Kremer, W.; Kalbitzer, H.R.; Winter, R. Cosolvent and Crowding Effects on the Temperature and Pressure Dependent Conformational Dynamics and Stability of Globular Actin. J. Phys. Chem. B 2016, 120, 6575–6586. [Google Scholar] [CrossRef] [PubMed]
- Frederick, K.B.; Sept, D.; De La Cruz, E.M. Effects of solution crowding on actin polymerization reveal the energetic basis for nucleotide-dependent filament stability. J. Mol. Biol. 2008, 378, 540–550. [Google Scholar] [CrossRef]
- Castaneda, N.; Lee, M.; Rivera-Jacquez, H.J.; Marracino, R.R.; Merlino, T.R.; Kang, H. Actin Filament Mechanics and Structure in Crowded Environments. J. Phys. Chem. B 2019, 123, 2770–2779. [Google Scholar] [CrossRef]
- Tellam, R.L.; Sculley, M.J.; Nichol, L.W.; Wills, P.R. The Influence of Poly(Ethylene Glycol) 6000 on the Properties of Skeletal-Muscle Actin. Biochem. J. 1983, 213, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Hosek, M.; Tang, J.X. Polymer-induced bundling of Factin and the depletion force. Phys. Rev. E 2004, 69, 051907. [Google Scholar] [CrossRef]
- Mukherjee, S.K.; Gautam, S.; Biswas, S.; Kundu, J.; Chowdhury, P.K. Do Macromolecular Crowding Agents Exert Only an Excluded Volume Effect? A Protein Solvation Study. J. Phys. Chem. B 2015, 119, 14145–14156. [Google Scholar] [CrossRef]
- Pardee, J.D.; Aspudich, J. [18] Purification of muscle actin. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1982; Volume 85, pp. 164–181. [Google Scholar]
- Kang, H.R.; Bradley, M.J.; McCullough, B.R.; Pierre, A.; Grintsevich, E.E.; Reisler, E.; De La Cruz, E.M. Identification of cation-binding sites on actin that drive polymerization and modulate bending stiffness. Proc. Natl. Acad. Sci. USA 2012, 109, 16923–16927. [Google Scholar] [CrossRef]
- Heidings, J.B.; Demosthene, B.; Merlino, T.R.; Castaneda, N.; Kang, E.H. Gelsolin-mediated actin filament severing in crowded environments. Biochem. Biophys. Res. Commun. 2020, 532, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Winterhoff, M.; Brühmann, S.; Franke, C.; Breitsprecher, D.; Faix, J. Visualization of Actin Assembly and Filament Turnover by In Vitro Multicolor TIRF Microscopy. Methods Mol. Biol. 2016, 1407, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kravchuk, P.; Krishnaprasad, A.; Roy, T.; Kang, E.H. Graphene Enhances Actin Filament Assembly Kinetics and Modulates NIH-3T3 Fibroblast Cell Spreading. Int. J. Mol. Sci. 2022, 23, 509. [Google Scholar] [CrossRef]
- Castaneda, N.; Zheng, T.Y.; Rivera-Jacquez, H.J.; Lee, H.-J.; Hyun, J.; Balaeff, A.; Huo, Q.; Kang, H. Cations Modulate Actin Bundle Mechanics, Assembly Dynamics, and Structure. J. Phys. Chem. B 2018, 122, 3826–3835. [Google Scholar] [CrossRef]
- Graham, J.S.; McCullough, B.R.; Kang, H.R.; Elam, W.A.; Cao, W.X.; De La Cruz, E.M. Multi-Platform Compatible Software for Analysis of Polymer Bending Mechanics. PLoS ONE 2014, 9, e94766. [Google Scholar] [CrossRef]
- Cooper, J.A.; Walker, S.B.; Pollard, T.D. Pyrene actin: Documentation of the validity of a sensitive assay for actin polymerization. J. Muscle Res. Cell Motil. 1983, 4, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, L.K.; Rosen, M.K.; Padrick, S.B. Measurement and Analysis of In Vitro Actin Polymerization. In Adhesion Protein Protocols, 3rd ed.; Humana Press: Totowa, NJ, USA, 2013; Volume 1046, pp. 273–293. [Google Scholar] [CrossRef]
- Galkin, V.E.; Orlova, A.; Vos, M.R.; Schröder, G.F.; Egelman, E.H. Near-Atomic Resolution for One State of F-Actin. Structure 2015, 23, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Lee, M.; Lee, B.; Castaneda, N.; Tetard, L.; Kang, E.H. Crowding tunes the organization and mechanics of actin bundles formed by crosslinking proteins. FEBS Lett. 2021, 595, 26–40. [Google Scholar] [CrossRef]
- Martínez, L.; Andrade, R.; Birgin, E.G.; Martínez, J.M. PACKMOL: A Package for Building Initial Configurations for Molecular Dynamics Simulations. J. Comput. Chem. 2009, 30, 2157–2164. [Google Scholar] [CrossRef]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [PubMed]
- Vorobyov, I.; Anisimov, V.M.; Greene, S.; Venable, R.M.; Moser, A.; Pastor, R.W.; MacKerell, A.D. Additive and classical drude polarizable force fields for linear and cyclic ethers. J. Chem. Theory Comput. 2007, 3, 1120–1133. [Google Scholar] [CrossRef]
- Guvench, O.; Mallajosyula, S.S.; Raman, E.P.; Hatcher, E.; Vanommeslaeghe, K.; Foster, T.J.; Jamison, I.F.W., 2nd; MacKerell, J.A.D., Jr. CHARMM Additive All-Atom Force Field for Carbohydrate Derivatives and Its Utility in Polysaccharide and Carbohydrate–Protein Modeling. J. Chem. Theory Comput. 2011, 7, 3162–3180. [Google Scholar] [CrossRef]
- Li, L.; Li, C.; Sarkar, S.; Zhang, J.; Witham, S.; Zhang, Z.; Wang, L.; Smith, N.; Petukh, M.; Alexov, E. DelPhi: A comprehensive suite for DelPhi software and associated resources. BMC Biophys. 2012, 5, 9. [Google Scholar] [CrossRef]
- Kuhn, J.R.; Pollard, T.D. Real-time measurements of actin filament polymerization by total internal reflection fluorescence microscopy. Biophys. J. 2005, 88, 1387–1402. [Google Scholar] [CrossRef]
- Crevenna, A.H.; Naredi-Rainer, N.; Schönichen, A.; Dzubiella, J.; Barber, D.L.; Lamb, D.C.; Wedlich-Söldner, R. Electrostatics control actin filament nucleation and elongation kinetics. J. Biol. Chem. 2013, 288, 12102–12113. [Google Scholar] [CrossRef]
- Kovar, D.R.; Harris, E.S.; Mahaffy, R.; Higgs, H.N.; Pollard, T.D. Control of the assembly of ATP- and ADP-actin by formins and profilin. Cell 2006, 124, 423–435. [Google Scholar] [CrossRef]
- Minton, A.P. The influence of macromolecular crowding and macromolecular confinement on biochemical reactions in physiological media. J. Biol. Chem. 2001, 276, 10577–10580. [Google Scholar] [CrossRef] [PubMed]
- Knowles, D.B.; Shkel, I.A.; Phan, N.M.; Sternke, M.; Lingeman, E.; Cheng, X.; Cheng, L.; O’connor, K.; Record, M.T. Chemical Interactions of Polyethylene Glycols (PEGs) and Glycerol with Protein Functional Groups: Applications to Effects of PEG and Glycerol on Protein Processes. Biochemistry 2015, 54, 3528–3542. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.J.; Olgenblum, G.I.; Propst, A.; Harries, D.; Pielak, G.J. Resolving the Enthalpy of Protein Stabilization by Macromolecular Crowding. Protein Sci. 2023, 32, e4573. [Google Scholar] [CrossRef]
- Sarkar, M.; Li, C.; Pielak, G.J. Soft interactions and crowding. Biophys. Rev. 2013, 5, 187–194. [Google Scholar] [CrossRef]
- Wang, P.-H.; Yu, I.; Feig, M.; Sugita, Y. Influence of protein crowder size on hydration structure and dynamics in macromolecular crowding. Chem. Phys. Lett. 2017, 671, 63–70. [Google Scholar] [CrossRef]
- Sarkar, M.; Lu, J.; Pielak, G.J. Protein crowder charge and protein stability. Biochemistry 2014, 53, 1601–1606. [Google Scholar] [CrossRef]
- Yadav, S.; Shire, S.J.; Kalonia, D.S. Viscosity Analysis of High Concentration Bovine Serum Albumin Aqueous Solutions. Pharm. Res. 2011, 28, 1973–1983. [Google Scholar] [CrossRef]
- Schummel, P.H.; Gao, M.; Winter, R. Modulation of the Polymerization Kinetics of α/β-Tubulin by Osmolytes and Macromolecular Crowding. ChemPhysChem 2017, 18, 189–197. [Google Scholar] [CrossRef]
- Zimmerle, C.T.; Frieden, C. Effect of pH on the mechanism of actin polymerization. Biochemistry 1988, 27, 7766–7772. [Google Scholar] [CrossRef] [PubMed]
- Feig, M.; Sugita, Y. Variable Interactions between Protein Crowders and Biomolecular Solutes Are Important in Understanding Cellular Crowding. J. Phys. Chem. B 2012, 116, 599–605. [Google Scholar] [CrossRef] [PubMed]
- McGrath, J.L.; Tardy, Y.; Dewey, C.F., Jr.; Meister, J.J.; Hartwig, J.H. Simultaneous measurements of actin filament turnover, filament fraction, and monomer diffusion in endothelial cells. Biophys. J. 1998, 75, 2070–2078. [Google Scholar] [CrossRef] [PubMed]
- Zicha, D.; Dobbie, I.M.; Holt, M.R.; Monypenny, J.; Soong, D.Y.; Gray, C.; Dunn, G.A. Rapid actin transport during cell protrusion. Science 2003, 300, 142–145. [Google Scholar] [CrossRef]
- Sept, D.; Elcock, A.H.; McCammon, J.A. Computer simulations of actin polymerization can explain the barbed-pointed end asymmetry. J. Mol. Biol. 1999, 294, 1181–1189. [Google Scholar] [CrossRef]
- Sept, D.; Xu, J.Y.; Pollard, T.D.; McCammon, J.A. Annealing accounts for the length of actin filaments formed by spontaneous polymerization. Biophys. J. 1999, 77, 2911–2919. [Google Scholar] [CrossRef] [PubMed]
- Harada, R.; Sugita, Y.; Feig, M. Protein Crowding Affects Hydration Structure and Dynamics. J. Am. Chem. Soc. 2012, 134, 4842–4849. [Google Scholar] [CrossRef]
- Harada, R.; Tochio, N.; Kigawa, T.; Sugita, Y.; Feig, M. Reduced Native State Stability in Crowded Cellular Environment Due to Protein–Protein Interactions. J. Am. Chem. Soc. 2013, 135, 3696–3701. [Google Scholar] [CrossRef] [PubMed]
- Nawrocki, G.; Karaboga, A.; Sugita, Y.; Feig, M. Effect of protein–protein interactions and solvent viscosity on the rotational diffusion of proteins in crowded environments. Phys. Chem. Chem. Phys. 2019, 21, 876–883. [Google Scholar] [CrossRef]
- Sept, D.; McCammon, J.A. Thermodynamics and kinetics of actin filament nucleation. Biophys. J. 2001, 81, 667–674. [Google Scholar] [CrossRef]
- Ohnuki, J.; Yodogawa, A.; Takano, M. Electrostatic balance between global repulsion and local attraction in reentrant polymerization of actin. Cytoskeleton 2017, 74, 504–511. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | PEG | BSA | Sucrose |
|---|---|---|---|
| 5.36 | 12.4 | 4.55 | 2.25 |
| Control | PEG | BSA | Sucrose |
|---|---|---|---|
| −1295.6 | −1373.08 | −34,752.64 | −1249.28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demosthene, B.; Lee, M.; Marracino, R.R.; Heidings, J.B.; Kang, E.H. Molecular Basis for Actin Polymerization Kinetics Modulated by Solution Crowding. Biomolecules 2023, 13, 786. https://doi.org/10.3390/biom13050786
Demosthene B, Lee M, Marracino RR, Heidings JB, Kang EH. Molecular Basis for Actin Polymerization Kinetics Modulated by Solution Crowding. Biomolecules. 2023; 13(5):786. https://doi.org/10.3390/biom13050786
Chicago/Turabian StyleDemosthene, Bryan, Myeongsang Lee, Ryan R. Marracino, James B. Heidings, and Ellen Hyeran Kang. 2023. "Molecular Basis for Actin Polymerization Kinetics Modulated by Solution Crowding" Biomolecules 13, no. 5: 786. https://doi.org/10.3390/biom13050786
APA StyleDemosthene, B., Lee, M., Marracino, R. R., Heidings, J. B., & Kang, E. H. (2023). Molecular Basis for Actin Polymerization Kinetics Modulated by Solution Crowding. Biomolecules, 13(5), 786. https://doi.org/10.3390/biom13050786