
Emerging Role and Mechanism of the FTO Gene in Cardiovascular Diseases

Abstract
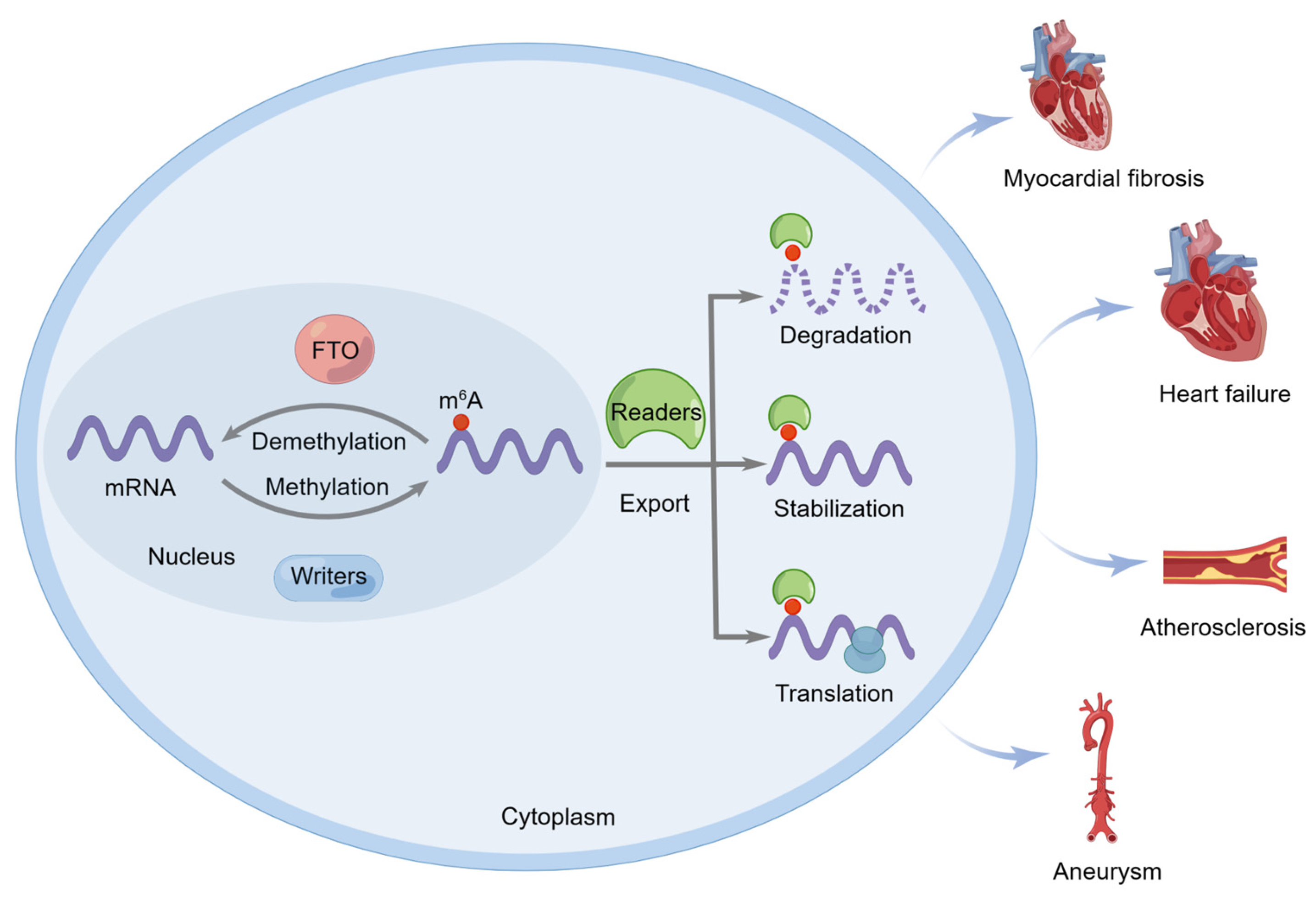
:1. Introduction
2. FTO Genetic Variants and CVD Risk
2.1. Incident CVD
2.2. Hypertension
2.3. Acute Coronary Syndrome
2.4. Aortic Valve Stenosis
2.5. Others
3. Role and Mechanism of FTO in CVDs
3.1. Myocardial Fibrosis
3.2. Myocarditis
3.3. Ischemia–Reperfusion Injury
3.4. Heart Failure
3.5. Atherosclerosis
3.6. Hypertension
3.7. Abdominal Aortic Aneurysm
3.8. Arrhythmia
4. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsao, C.W.; Aday, A.W.; Almarzooq, Z.I.; Alonso, A.; Beaton, A.Z.; Bittencourt, M.S.; Boehme, A.K.; Buxton, A.E.; Carson, A.P.; Commodore-Mensah, Y.; et al. Heart Disease and Stroke Statistics-2022 Update: A Report From the American Heart Association. Circulation 2022, 145, e153–e639. [Google Scholar] [CrossRef] [PubMed]
- Frayling, T.M.; Timpson, N.J.; Weedon, M.N.; Zeggini, E.; Freathy, R.M.; Lindgren, C.M.; Perry, J.R.; Elliott, K.S.; Lango, H.; Rayner, N.W.; et al. A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science 2007, 316, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Gerken, T.; Girard, C.A.; Tung, Y.C.; Webby, C.J.; Saudek, V.; Hewitson, K.S.; Yeo, G.S.; McDonough, M.A.; Cunliffe, S.; McNeill, L.A.; et al. The obesity-associated FTO gene encodes a 2-oxoglutarate-dependent nucleic acid demethylase. Science 2007, 318, 1469–1472. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Yang, C.G.; Yang, S.; Jian, X.; Yi, C.; Zhou, Z.; He, C. Oxidative demethylation of 3-methylthymine and 3-methyluracil in single-stranded DNA and RNA by mouse and human FTO. FEBS Lett. 2008, 582, 3313–3319. [Google Scholar] [CrossRef]
- Sanchez-Pulido, L.; Andrade-Navarro, M.A. The FTO (fat mass and obesity associated) gene codes for a novel member of the non-heme dioxygenase superfamily. BMC Biochem. 2007, 8, 23. [Google Scholar] [CrossRef]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.G.; et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef]
- Wei, J.; Liu, F.; Lu, Z.; Fei, Q.; Ai, Y.; He, P.C.; Shi, H.; Cui, X.; Su, R.; Klungland, A.; et al. Differential m6A, m6Am, and m1A Demethylation Mediated by FTO in the Cell Nucleus and Cytoplasm. Mol. Cell 2018, 71, 973–985.e5. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, L.H.; Wang, Y.; Xiao, Y.; Liu, J.; Zhang, W.; Yan, N.; Amu, G.; Tang, X.; Zhang, L.; et al. Structural insights into FTO’s catalytic mechanism for the demethylation of multiple RNA substrates. Proc. Natl. Acad. Sci. USA 2019, 116, 2919–2924. [Google Scholar] [CrossRef]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N(6)-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef]
- Alarcón, C.R.; Goodarzi, H.; Lee, H.; Liu, X.; Tavazoie, S.; Tavazoie, S.F. HNRNPA2B1 Is a Mediator of m(6)A-Dependent Nuclear RNA Processing Events. Cell 2015, 162, 1299–1308. [Google Scholar] [CrossRef]
- Han, D.; Liu, J.; Chen, C.; Dong, L.; Liu, Y.; Chang, R.; Huang, X.; Liu, Y.; Wang, J.; Dougherty, U.; et al. Anti-tumour immunity controlled through mRNA m6A methylation and YTHDF1 in dendritic cells. Nature 2019, 566, 270–274. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef]
- Liu, T.; Wei, Q.; Jin, J.; Luo, Q.; Liu, Y.; Yang, Y.; Cheng, C.; Li, L.; Pi, J.; Si, Y.; et al. The m6A reader YTHDF1 promotes ovarian cancer progression via augmenting EIF3C translation. Nucleic Acids Res. 2020, 48, 3816–3831. [Google Scholar] [CrossRef]
- Yang, S.; Wei, J.; Cui, Y.H.; Park, G.; Shah, P.; Deng, Y.; Aplin, A.E.; Lu, Z.; Hwang, S.; He, C.; et al. m6A mRNA demethylase FTO regulates melanoma tumorigenicity and response to anti-PD-1 blockade. Nat. Commun. 2019, 10, 2782. [Google Scholar] [CrossRef]
- Shen, F.; Huang, W.; Huang, J.T.; Xiong, J.; Yang, Y.; Wu, K.; Jia, G.F.; Chen, J.; Feng, Y.Q.; Yuan, B.F.; et al. Decreased N(6)-methyladenosine in peripheral blood RNA from diabetic patients is associated with FTO expression rather than ALKBH5. J. Clin. Endocrinol. Metab. 2015, 100, E148–E154. [Google Scholar] [CrossRef]
- Huo, Y.B.; Gao, X.; Peng, Q.; Nie, Q.; Bi, W. Dihydroartemisinin alleviates AngII-induced vascular smooth muscle cell proliferation and inflammatory response by blocking the FTO/NR4A3 axis. Inflamm. Res. 2022, 71, 243–253. [Google Scholar] [CrossRef]
- Frazer, K.A.; Ballinger, D.G.; Cox, D.R.; Hinds, D.A.; Stuve, L.L.; Gibbs, R.A.; Belmont, J.W.; Boudreau, A.; Hardenbol, P.; Leal, S.M.; et al. A second generation human haplotype map of over 3.1 million SNPs. Nature 2007, 449, 851–861. [Google Scholar]
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and cardiovascular disease: Pathophysiology, evaluation, and effect of weight loss: An update of the 1997 American Heart Association Scientific Statement on Obesity and Heart Disease from the Obesity Committee of the Council on Nutrition, Physical Activity, and Metabolism. Circulation 2006, 113, 898–918. [Google Scholar]
- Ahmad, T.; Chasman, D.I.; Mora, S.; Pare, G.; Cook, N.R.; Buring, J.E.; Ridker, P.M.; Lee, I.M. The fat-mass and obesity-associated (FTO) gene, physical activity, and risk of incident cardiovascular events in white women. Am. Heart J. 2010, 160, 1163–1169. [Google Scholar] [CrossRef]
- Borglykke, A.; Grarup, N.; Sparso, T.; Linneberg, A.; Fenger, M.; Jeppesen, J.; Hansen, T.; Pedersen, O.; Jorgensen, T. Genetic variant SLC2A2 [corrected] Is associated with risk of cardiovascular disease-assessing the individual and cumulative effect of 46 type 2 diabetes related genetic variants. PLoS ONE 2012, 7, e50418. [Google Scholar] [CrossRef]
- He, M.; Cornelis, M.C.; Franks, P.W.; Zhang, C.; Hu, F.B.; Qi, L. Obesity genotype score and cardiovascular risk in women with type 2 diabetes mellitus. Arter. Thromb. Vasc. Biol. 2010, 30, 327–332. [Google Scholar] [CrossRef]
- Lappalainen, T.; Kolehmainen, M.; Schwab, U.S.; Tolppanen, A.M.; Stancakova, A.; Lindstrom, J.; Eriksson, J.G.; Keinanen-Kiukaanniemi, S.; Aunola, S.; Ilanne-Parikka, P.; et al. Association of the FTO gene variant (rs9939609) with cardiovascular disease in men with abnormal glucose metabolism—The Finnish Diabetes Prevention Study. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Aijala, M.; Ronkainen, J.; Huusko, T.; Malo, E.; Savolainen, E.R.; Savolainen, M.J.; Salonurmi, T.; Bloigu, R.; Kesaniemi, Y.A.; Ukkola, O. The fat mass and obesity-associated (FTO) gene variant rs9939609 predicts long-term incidence of cardiovascular disease and related death independent of the traditional risk factors. Ann. Med. 2015, 47, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Timpson, N.J.; Harbord, R.; Davey Smith, G.; Zacho, J.; Tybjaerg-Hansen, A.; Nordestgaard, B.G. Does greater adiposity increase blood pressure and hypertension risk?: Mendelian randomization using the FTO/MC4R genotype. Hypertension 2009, 54, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Xi, B.; Zhang, M.; Wang, C.; Shen, Y.; Zhao, X.; Wang, X.; Mi, J. The common SNP (rs9939609) in the FTO gene modifies the association between obesity and high blood pressure in Chinese children. Mol. Biol. Rep. 2013, 40, 773–778. [Google Scholar] [CrossRef]
- Xi, B.; Zhao, X.; Shen, Y.; Wu, L.; Hotta, K.; Hou, D.; Cheng, H.; Wang, X.; Mi, J. Associations of obesity susceptibility loci with hypertension in Chinese children. Int. J. Obes. 2013, 37, 926–930. [Google Scholar] [CrossRef]
- Garcia-Solis, P.; Reyes-Bastidas, M.; Flores, K.; Garcia, O.P.; Rosado, J.L.; Mendez-Villa, L.; Garcia, G.C.; Garcia-Gutierrez, D.; Kuri-Garcia, A.; Hernandez-Montiel, H.L.; et al. Fat mass obesity-associated (FTO) (rs9939609) and melanocortin 4 receptor (MC4R) (rs17782313) SNP are positively associated with obesity and blood pressure in Mexican school-aged children. Br. J. Nutr. 2016, 116, 1834–1840. [Google Scholar] [CrossRef]
- Sun, Y.; Sun, J.; Wu, J.; Yang, M. Combined effects of FTO rs9939609 and MC4R rs17782313 on elevated nocturnal blood pressure in the Chinese Han population. Cardiovasc. J. Afr. 2016, 27, 21–24. [Google Scholar] [CrossRef]
- He, D.; Fu, M.; Miao, S.; Hotta, K.; Chandak, G.R.; Xi, B. FTO gene variant and risk of hypertension: A meta-analysis of 57,464 hypertensive cases and 41,256 controls. Metabolism 2014, 63, 633–639. [Google Scholar] [CrossRef]
- Pausova, Z.; Syme, C.; Abrahamowicz, M.; Xiao, Y.; Leonard, G.T.; Perron, M.; Richer, L.; Veillette, S.; Smith, G.D.; Seda, O.; et al. A common variant of the FTO gene is associated with not only increased adiposity but also elevated blood pressure in French Canadians. Circ. Cardiovasc. Genet. 2009, 2, 260–269. [Google Scholar] [CrossRef]
- Falbova, D.; Vorobelova, L.; Sivakova, D.; Benus, R. Association between FTO (rs17817449) genetic variant, gamma-glutamyl transferase, and hypertension in Slovak midlife women. Am. J. Hum. Biol. 2022, 34, e23672. [Google Scholar] [CrossRef]
- Kumar, R.; Kohli, S.; Alam, P.; Barkotoky, R.; Gupta, M.; Tyagi, S.; Jain, S.K.; Pasha, M.A. Interactions between the FTO and GNB3 genes contribute to varied clinical phenotypes in hypertension. PLoS ONE 2013, 8, e63934. [Google Scholar] [CrossRef]
- Goulet, D.; O’Loughlin, J.; Sylvestre, M.P. Association of Genetic Variants With Body-Mass Index and Blood Pressure in Adolescents: A Replication Study. Front. Genet. 2021, 12, 690335. [Google Scholar] [CrossRef]
- Hubacek, J.A.; Stanek, V.; Gebauerova, M.; Pilipcincova, A.; Dlouha, D.; Poledne, R.; Aschermann, M.; Skalicka, H.; Matouskova, J.; Kruger, A.; et al. A FTO variant and risk of acute coronary syndrome. Clin. Chim. Acta 2010, 411, 1069–1072. [Google Scholar] [CrossRef]
- Thron, C.; Akhyari, P.; Godehardt, E.; Lichtenberg, A.; Ruther, U.; Seehaus, S. FTO Is Associated with Aortic Valve Stenosis in a Gender Specific Manner of Heterozygote Advantage: A Population-Based Case-Control Study. PLoS ONE 2015, 10, e0139419. [Google Scholar] [CrossRef]
- Small, A.M.; Peloso, G.M.; Linefsky, J.; Aragam, J.; Galloway, A.; Tanukonda, V.; Wang, L.C.; Yu, Z.; Sunitha Selvaraj, M.; Farber-Eger, E.H.; et al. Multiancestry Genome-Wide Association Study of Aortic Stenosis Identifies Multiple Novel Loci in the Million Veteran Program. Circulation 2023, 147, 942–955. [Google Scholar] [CrossRef]
- Klovaite, J.; Benn, M.; Nordestgaard, B.G. Obesity as a causal risk factor for deep venous thrombosis: A Mendelian randomization study. J. Intern. Med. 2015, 277, 573–584. [Google Scholar] [CrossRef]
- Lindstrom, S.; Germain, M.; Crous-Bou, M.; Smith, E.N.; Morange, P.E.; van Hylckama Vlieg, A.; de Haan, H.G.; Chasman, D.; Ridker, P.; Brody, J.; et al. Assessing the causal relationship between obesity and venous thromboembolism through a Mendelian Randomization study. Hum. Genet. 2017, 136, 897–902. [Google Scholar] [CrossRef]
- Hubacek, J.A.; Vrablik, M.; Dlouha, D.; Stanek, V.; Gebauerova, M.; Adamkova, V.; Ceska, R.; Dostalova, G.; Linhart, A.; Vitek, L.; et al. Gene variants at FTO, 9p21, and 2q36.3 are age-independently associated with myocardial infarction in Czech men. Clin. Chim. Acta 2016, 454, 119–123. [Google Scholar] [CrossRef]
- Berzuini, C.; Vansteelandt, S.; Foco, L.; Pastorino, R.; Bernardinelli, L. Direct genetic effects and their estimation from matched case-control data. Genet. Epidemiol. 2012, 36, 652–662. [Google Scholar] [CrossRef]
- Doney, A.S.; Dannfald, J.; Kimber, C.H.; Donnelly, L.A.; Pearson, E.; Morris, A.D.; Palmer, C.N. The FTO gene is associated with an atherogenic lipid profile and myocardial infarction in patients with type 2 diabetes: A Genetics of Diabetes Audit and Research Study in Tayside Scotland (Go-DARTS) study. Circ. Cardiovasc. Genet. 2009, 2, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Nordestgaard, B.G.; Palmer, T.M.; Benn, M.; Zacho, J.; Tybjaerg-Hansen, A.; Davey Smith, G.; Timpson, N.J. The effect of elevated body mass index on ischemic heart disease risk: Causal estimates from a Mendelian randomisation approach. PLoS Med. 2012, 9, e1001212. [Google Scholar] [CrossRef] [PubMed]
- Shahid, S.U.; Shabana; Rehman, A.; Hasnain, S. Role of a common variant of Fat Mass and Obesity associated (FTO) gene in obesity and coronary artery disease in subjects from Punjab, Pakistan: A case control study. Lipids Health Dis. 2016, 15, 29. [Google Scholar] [CrossRef] [PubMed]
- Shahid, S.U.; Shabana; Cooper, J.A.; Beaney, K.E.; Li, K.; Rehman, A.; Humphries, S.E. Genetic risk analysis of coronary artery disease in Pakistani subjects using a genetic risk score of 21 variants. Atherosclerosis 2017, 258, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Qiu, L.; Hu, Z.; Liu, J.; Liu, D.; Hou, D. Evaluation of the Obesity Genes FTO and MC4R for Contribution to the Risk of Large Artery Atherosclerotic Stroke in a Chinese Population. Obes. Facts 2016, 9, 353–362. [Google Scholar] [CrossRef]
- Winter, Y.; Back, T.; Scherag, A.; Linseisen, J.; Rohrmann, S.; Lanczik, O.; Hinney, A.; Scherag, S.; Neumaier, M.; Ringleb, P.A.; et al. Evaluation of the obesity genes FTO and MC4R and the type 2 diabetes mellitus gene TCF7L2 for contribution to stroke risk: The Mannheim-Heidelberg Stroke Study. Obes. Facts 2011, 4, 290–296. [Google Scholar] [CrossRef]
- Gulati, P.; Cheung, M.K.; Antrobus, R.; Church, C.D.; Harding, H.P.; Tung, Y.C.; Rimmington, D.; Ma, M.; Ron, D.; Lehner, P.J.; et al. Role for the obesity-related FTO gene in the cellular sensing of amino acids. Proc. Natl. Acad. Sci. USA 2013, 110, 2557–2562. [Google Scholar] [CrossRef]
- Cecil, J.E.; Tavendale, R.; Watt, P.; Hetherington, M.M.; Palmer, C.N.A. An obesity-associated FTO gene variant and increased energy intake in children. N. Engl. J. Med. 2008, 359, 2558–2566. [Google Scholar] [CrossRef]
- Udali, S.; Guarini, P.; Moruzzi, S.; Choi, S.-W.; Friso, S. Cardiovascular epigenetics: From DNA methylation to microRNAs. Mol. Asp. Med. 2013, 34, 883–901. [Google Scholar] [CrossRef]
- Aslibekyan, S.; Claas, S.A.; Arnett, D.K. Clinical applications of epigenetics in cardiovascular disease: The long road ahead. Transl. Res. 2015, 165, 143–153. [Google Scholar] [CrossRef]
- Bell, C.G.; Finer, S.; Lindgren, C.M.; Wilson, G.A.; Rakyan, V.K.; Teschendorff, A.E.; Akan, P.; Stupka, E.; Down, T.A.; Prokopenko, I.; et al. Integrated genetic and epigenetic analysis identifies haplotype-specific methylation in the FTO type 2 diabetes and obesity susceptibility locus. PLoS ONE 2010, 5, e14040. [Google Scholar] [CrossRef]
- Gluckman, P.D. Epigenetics and metabolism in 2011: Epigenetics, the life-course and metabolic disease. Nat. Rev. Endocrinol. 2011, 8, 74–76. [Google Scholar] [CrossRef]
- Li, X.X.; Mu, B.; Li, X.; Bie, Z.D. circCELF1 Inhibits Myocardial Fibrosis by Regulating the Expression of DKK2 Through FTO/m(6)A and miR-636. J. Cardiovasc. Transl. Res. 2022, 15, 998–1009. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, Y.; Liu, J.; Li, X.; Sang, Y.; Zhou, G.; Xue, J.; Jing, L.; Shi, Z.; Wei, J.; et al. Fat mass and obesity-associated gene (FTO) hypermethylation induced by decabromodiphenyl ethane causing cardiac dysfunction via glucolipid metabolism disorder. Ecotoxicol. Environ. Saf. 2022, 237, 113534. [Google Scholar] [CrossRef]
- Mathiyalagan, P.; Adamiak, M.; Mayourian, J.; Sassi, Y.; Liang, Y.; Agarwal, N.; Jha, D.; Zhang, S.; Kohlbrenner, E.; Chepurko, E.; et al. FTO-Dependent N(6)-Methyladenosine Regulates Cardiac Function During Remodeling and Repair. Circulation 2019, 139, 518–532. [Google Scholar] [CrossRef]
- Yu, Y.; Pan, Y.; Fan, Z.; Xu, S.; Gao, Z.; Ren, Z.; Yu, J.; Li, W.; Liu, F.; Gu, J.; et al. LuHui Derivative, A Novel Compound That Inhibits the Fat Mass and Obesity-Associated (FTO), Alleviates the Inflammatory Response and Injury in Hyperlipidemia-Induced Cardiomyopathy. Front. Cell Dev. Biol. 2021, 9, 731365. [Google Scholar] [CrossRef]
- Dubey, P.K.; Patil, M.; Singh, S.; Dubey, S.; Ahuja, P.; Verma, S.K.; Krishnamurthy, P. Increased m6A-RNA methylation and FTO suppression is associated with myocardial inflammation and dysfunction during endotoxemia in mice. Mol. Cell. Biochem. 2022, 477, 129–141. [Google Scholar] [CrossRef]
- Ke, W.L.; Huang, Z.W.; Peng, C.L.; Ke, Y.P. m6A demethylase FTO regulates the apoptosis and inflammation of cardiomyocytes via YAP1 in ischemia-reperfusion injury. Bioengineered 2022, 13, 5443–5452. [Google Scholar] [CrossRef]
- Xu, K.; Mo, Y.; Li, D.; Yu, Q.; Wang, L.; Lin, F.; Kong, C.; Balelang, M.F.; Zhang, A.; Chen, S.; et al. N(6)-methyladenosine demethylases Alkbh5/Fto regulate cerebral ischemia-reperfusion injury. Ther. Adv. Chronic Dis. 2020, 11, 2040622320916024. [Google Scholar] [CrossRef]
- Hou, L.; Li, S.; Li, S.; Wang, R.; Zhao, M.; Liu, X. FTO inhibits oxidative stress by mediating m6A demethylation of Nrf2 to alleviate cerebral ischemia/reperfusion injury. J. Physiol. Biochem. 2023, 79, 133–146. [Google Scholar] [CrossRef]
- Du, Y.D.; Guo, W.Y.; Han, C.H.; Wang, Y.; Chen, X.S.; Li, D.W.; Liu, J.L.; Zhang, M.; Zhu, N.; Wang, X. N6-methyladenosine demethylase FTO impairs hepatic ischemia-reperfusion injury via inhibiting Drp1-mediated mitochondrial fragmentation. Cell. Death Dis. 2021, 12, 442. [Google Scholar] [CrossRef] [PubMed]
- Chokkalla, A.K.; Jeong, S.; Mehta, S.L.; Davis, C.K.; Morris-Blanco, K.C.; Bathula, S.; Qureshi, S.S.; Vemuganti, R. Cerebroprotective Role of N6-Methyladenosine Demethylase FTO (Fat Mass and Obesity-Associated Protein) After Experimental Stroke. Stroke 2023, 54, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Xi, W.; Bai, Y.; Liu, X.; Zhang, Y.; Li, L.; Bian, L.; Liu, C.; Tang, Y.; Shen, L.; et al. FTO-dependent m6A modification of Plpp3 in circSCMH1-regulated vascular repair and functional recovery following stroke. Nat. Commun. 2023, 14, 489. [Google Scholar] [CrossRef] [PubMed]
- Berulava, T.; Buchholz, E.; Elerdashvili, V.; Pena, T.; Islam, M.R.; Lbik, D.; Mohamed, B.A.; Renner, A.; von Lewinski, D.; Sacherer, M.; et al. Changes in m6A RNA methylation contribute to heart failure progression by modulating translation. Eur. J. Heart Fail. 2020, 22, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Xu, Y.; Cui, X.; Jiang, H.; Luo, W.; Weng, X.; Wang, Y.; Zhao, Y.; Sun, A.; Ge, J. Alteration of m6A RNA Methylation in Heart Failure With Preserved Ejection Fraction. Front. Cardiovasc. Med. 2021, 8, 647806. [Google Scholar] [CrossRef]
- Shen, W.; Li, H.; Su, H.; Chen, K.; Yan, J. FTO overexpression inhibits apoptosis of hypoxia/reoxygenation-treated myocardial cells by regulating m6A modification of Mhrt. Mol. Cell. Biochem. 2021, 476, 2171–2179. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Y.; Guo, R.; Zhao, L.; Yan, J.; Gao, C. Comprehensive Analysis of N6-Methyladenosine RNA Methylation Regulators in the Diagnosis and Subtype Classification of Acute Myocardial Infarction. J. Immunol. Res. 2022, 2022, 5173761. [Google Scholar] [CrossRef]
- Zhang, B.; Jiang, H.; Wu, J.; Cai, Y.; Dong, Z.; Zhao, Y.; Hu, Q.; Hu, K.; Sun, A.; Ge, J. m6A demethylase FTO attenuates cardiac dysfunction by regulating glucose uptake and glycolysis in mice with pressure overload-induced heart failure. Signal Transduct. Target. Ther. 2021, 6, 377. [Google Scholar] [CrossRef]
- Mo, C.; Yang, M.; Han, X.; Li, J.; Gao, G.; Tai, H.; Huang, N.; Xiao, H. Fat mass and obesity-associated protein attenuates lipid accumulation in macrophage foam cells and alleviates atherosclerosis in apolipoprotein E-deficient mice. J. Hypertens. 2017, 35, 810–821. [Google Scholar] [CrossRef]
- Mo, W.; Chen, Z.; Zhang, X.; Dai, G.; Ma, D.; Pan, J.; Zhang, X.; Wu, G.; Fan, W. N6-Methyladenosine Demethylase FTO (Fat Mass and Obesity-Associated Protein) as a Novel Mediator of Statin Effects in Human Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 644–658. [Google Scholar] [CrossRef]
- Kruger, N.; Biwer, L.A.; Good, M.E.; Ruddiman, C.A.; Wolpe, A.G.; DeLalio, L.J.; Murphy, S.; Macal, E.H., Jr.; Ragolia, L.; Serbulea, V.; et al. Loss of Endothelial FTO Antagonizes Obesity-Induced Metabolic and Vascular Dysfunction. Circ. Res. 2020, 126, 232–242. [Google Scholar] [CrossRef]
- He, Y.; Xing, J.; Wang, S.; Xin, S.; Han, Y.; Zhang, J. Increased m6A methylation level is associated with the progression of human abdominal aortic aneurysm. Ann. Transl. Med. 2019, 7, 797. [Google Scholar] [CrossRef]
- Carnevali, L.; Graiani, G.; Rossi, S.; Al Banchaabouchi, M.; Macchi, E.; Quaini, F.; Rosenthal, N.; Sgoifo, A. Signs of cardiac autonomic imbalance and proarrhythmic remodeling in FTO deficient mice. PLoS ONE 2014, 9, e95499. [Google Scholar] [CrossRef]
- Claussnitzer, M.; Dankel, S.N.; Kim, K.H.; Quon, G.; Meuleman, W.; Haugen, C.; Glunk, V.; Sousa, I.S.; Beaudry, J.L.; Puviindran, V.; et al. FTO Obesity Variant Circuitry and Adipocyte Browning in Humans. N. Engl. J. Med. 2015, 373, 895–907. [Google Scholar] [CrossRef]
- Postma, A.V.; Christoffels, V.M.; Bezzina, C.R. Developmental aspects of cardiac arrhythmogenesis. Cardiovasc. Res. 2011, 91, 243–251. [Google Scholar] [CrossRef]
- Zhang, S.S.; Kim, K.H.; Rosen, A.; Smyth, J.W.; Sakuma, R.; Delgado-Olguin, P.; Davis, M.; Chi, N.C.; Puviindran, V.; Gaborit, N.; et al. Iroquois homeobox gene 3 establishes fast conduction in the cardiac His-Purkinje network. Proc. Natl. Acad. Sci. USA 2011, 108, 13576–13581. [Google Scholar] [CrossRef]
- Stratigopoulos, G.; Martin Carli, J.F.; O’Day, D.R.; Wang, L.; Leduc, C.A.; Lanzano, P.; Chung, W.K.; Rosenbaum, M.; Egli, D.; Doherty, D.A.; et al. Hypomorphism for RPGRIP1L, a ciliary gene vicinal to the FTO locus, causes increased adiposity in mice. Cell. Metab. 2014, 19, 767–779. [Google Scholar] [CrossRef]
- Zhou, L.L.; Xu, H.; Huang, Y.; Yang, C.G. Targeting the RNA demethylase FTO for cancer therapy. RSC Chem. Biol. 2021, 2, 1352–1369. [Google Scholar] [CrossRef]
- Chen, B.; Ye, F.; Yu, L.; Jia, G.; Huang, X.; Zhang, X.; Peng, S.; Chen, K.; Wang, M.; Gong, S.; et al. Development of cell-active N6-methyladenosine RNA demethylase FTO inhibitor. J. Am. Chem. Soc. 2012, 134, 17963–17971. [Google Scholar] [CrossRef]
- Huang, Y.; Su, R.; Sheng, Y.; Dong, L.; Dong, Z.; Xu, H.; Ni, T.; Zhang, Z.S.; Zhang, T.; Li, C.; et al. Small-Molecule Targeting of Oncogenic FTO Demethylase in Acute Myeloid Leukemia. Cancer Cell 2019, 35, 677–691.e10. [Google Scholar] [CrossRef]
- Su, R.; Dong, L.; Li, Y.; Gao, M.; Han, L.; Wunderlich, M.; Deng, X.; Li, H.; Huang, Y.; Gao, L.; et al. Targeting FTO Suppresses Cancer Stem Cell Maintenance and Immune Evasion. Cancer Cell 2020, 38, 79–96.e11. [Google Scholar] [CrossRef] [PubMed]
- Huff, S.; Tiwari, S.K.; Gonzalez, G.M.; Wang, Y.; Rana, T.M. m6A-RNA Demethylase FTO Inhibitors Impair Self-Renewal in Glioblastoma Stem Cells. ACS Chem. Biol. 2021, 16, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Mauer, J.; Luo, X.; Blanjoie, A.; Jiao, X.; Grozhik, A.V.; Patil, D.P.; Linder, B.; Pickering, B.F.; Vasseur, J.J.; Chen, Q.; et al. Reversible methylation of m6Am in the 5′ cap controls mRNA stability. Nature 2017, 541, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Dong, L.; Li, C.; Nachtergaele, S.; Wunderlich, M.; Qing, Y.; Deng, X.; Wang, Y.; Weng, X.; Hu, C.; et al. R-2HG Exhibits Anti-tumor Activity by Targeting FTO/m6A/MYC/CEBPA Signaling. Cell 2018, 172, 90–105.e23. [Google Scholar] [CrossRef]
- Akichika, S.; Hirano, S.; Shichino, Y.; Suzuki, T.; Nishimasu, H.; Ishitani, R.; Sugita, A.; Hirose, Y.; Iwasaki, S.; Nureki, O.; et al. Cap-specific terminal N 6-methylation of RNA by an RNA polymerase II-associated methyltransferase. Science 2019, 363, eaav0080. [Google Scholar] [CrossRef]
- Sendinc, E.; Valle-Garcia, D.; Dhall, A.; Chen, H.; Henriques, T.; Navarrete-Perea, J.; Sheng, W.; Gygi, S.P.; Adelman, K.; Shi, Y. PCIF1 Catalyzes m6Am mRNA Methylation to Regulate Gene Expression. Mol. Cell 2019, 75, 620–630.e9. [Google Scholar] [CrossRef]
- Zhuo, W.; Sun, M.; Wang, K.; Zhang, L.; Li, K.; Yi, D.; Li, M.; Sun, Q.; Ma, X.; Liu, W.; et al. m6Am methyltransferase PCIF1 is essential for aggressiveness of gastric cancer cells by inhibiting TM9SF1 mRNA translation. Cell Discov. 2022, 8, 48. [Google Scholar] [CrossRef]


{kind=link}
| Diseases | SNP | Population | Association | References |
|---|---|---|---|---|
| Incident CVD | rs8050136 | Caucasian, Danish | + | [19,20] |
| rs9939609 | American, Finnish and Finland | + | [21,22,23] | |
| Hypertension | rs9939609 | Danish, Japanese, Chinese, Mexican, Indian | + | [24,25,26,27,28,29] |
| rs9302652 | Canadian | + | [30] | |
| rs17817449 | Slovak | + | [31] | |
| rs8050136, rs9926289 | Indian | + | [32] | |
| rs9930333 | Canadian | - | [33] | |
| Acute coronary syndrome | rs17817449 | Czech | + | [34] |
| Aortic valve stenosis | rs9939609 | German | + | [35] |
| rs11647020 | White | + | [36] | |
| Venous thromboembolism | rs9939609 | Danish | + | [37] |
| rs1558902 | French, Dutch, American | + | [38] | |
| Myocardial infarction | rs17817449 | Czech | + | [39] |
| rs9939609 | Britisher | + | [40,41] | |
| Ischemic heart disease | rs9939609 | Danish | + | [42] |
| Coronary artery disease | rs9939609 | Pakistani | + | [43,44] |
| Stroke | rs8050136, rs9939609 | Chinese | - | [45] |
| rs9937053 | German | - | [46] |
| Diseases | Target RNA | Function | References |
|---|---|---|---|
| Myocardial fibrosis | DDK2 | Regulated the activation, viability, and migration of cardiac fibroblasts | [53] |
| - | Suppressed cardiomyocytes apoptosis and myocardial dysfunction | [54] | |
| Myocarditis | IL-6, IL-1β, TNF-α | Influenced the expression of proinflammatory factors to regulate myocarditis and dysfunction during endotoxemia | [56] |
| CD36 | Increased CD36 expression and suppressed anti-inflammatory effects | [57] | |
| Ischemia– reperfusion injury | YAP1 | Regulated the apoptosis and inflammation of H/R-induced cardiomyocytes | [58] |
| Bcl2 | Regulated I/R-induced neuronal apoptosis | [59] | |
| Nrf2 | Inhibited the oxidative stress response and ultimately alleviated cerebral I/R injury | [60] | |
| Drp1 | Contributed to the hepatic protective effect via impairing Drp1-mediated mitochondrial fragmentation | [61] | |
| - | Decreased poststroke gray and white matter damage and improved motor function recovery | [62] | |
| Plpp3 | Contributed to vessel regeneration and constituted a more mature vessel network after stroke | [63] | |
| Heart failure | - | Promoted heart recovery and alleviated HF | [64] |
| - | Upregulated in the peripheral blood samples of patients with heart failure and might serve as a novel diagnostic biomarker of heart failure | [65] | |
| Mhrt | Inhibited apoptosis of H/R-treated myocardial cells | [66] | |
| SERCA2A, MYH6, RYR2 | Restored cardiac contractile protein expression to increase the contractile function of cardiomyocytes, reduce cardiac fibrosis, and boost angiogenesis in ischemic myocardium | [55] | |
| - | Affected the contractile ability of hypoxic cardiomyocytes | [67] | |
| Pgam2, Akt-GLUT4 | Attenuated cardiomyocyte contractile dysfunction by regulating glucose uptake and glycolysis | [68] | |
| Atherosclerosis | PPARγ | Inhibited macrophage lipid uptake and accelerated intracellular cholesterol efflux | [69] |
| eNOS, KLF2 | Regulated the expression of atheroprotective genes and affected endothelial cell inflammatory responses | [70] | |
| Hypertension | PDG2 | Affected the myogenic tone in resistance arteries | [71] |
| Aneurysm | - | Associated with aneurysmal smooth muscle cells and might play an important role in the progression of aneurysm | [72] |
| Arrhythmia | - | Affected the autonomic neural modulation of cardiac function in the sympathetic direction | [73] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Z.-Y.; Jing, X.; Xiong, X.-D. Emerging Role and Mechanism of the FTO Gene in Cardiovascular Diseases. Biomolecules 2023, 13, 850. https://doi.org/10.3390/biom13050850
Xu Z-Y, Jing X, Xiong X-D. Emerging Role and Mechanism of the FTO Gene in Cardiovascular Diseases. Biomolecules. 2023; 13(5):850. https://doi.org/10.3390/biom13050850
Chicago/Turabian StyleXu, Zi-Yang, Xia Jing, and Xing-Dong Xiong. 2023. "Emerging Role and Mechanism of the FTO Gene in Cardiovascular Diseases" Biomolecules 13, no. 5: 850. https://doi.org/10.3390/biom13050850
APA StyleXu, Z.-Y., Jing, X., & Xiong, X.-D. (2023). Emerging Role and Mechanism of the FTO Gene in Cardiovascular Diseases. Biomolecules, 13(5), 850. https://doi.org/10.3390/biom13050850