
Leaflet Tensions Control the Spatio-Temporal Remodeling of Lipid Bilayers and Nanovesicles

 ,
,  , , and
, , and {kind=link}
{kind=link}
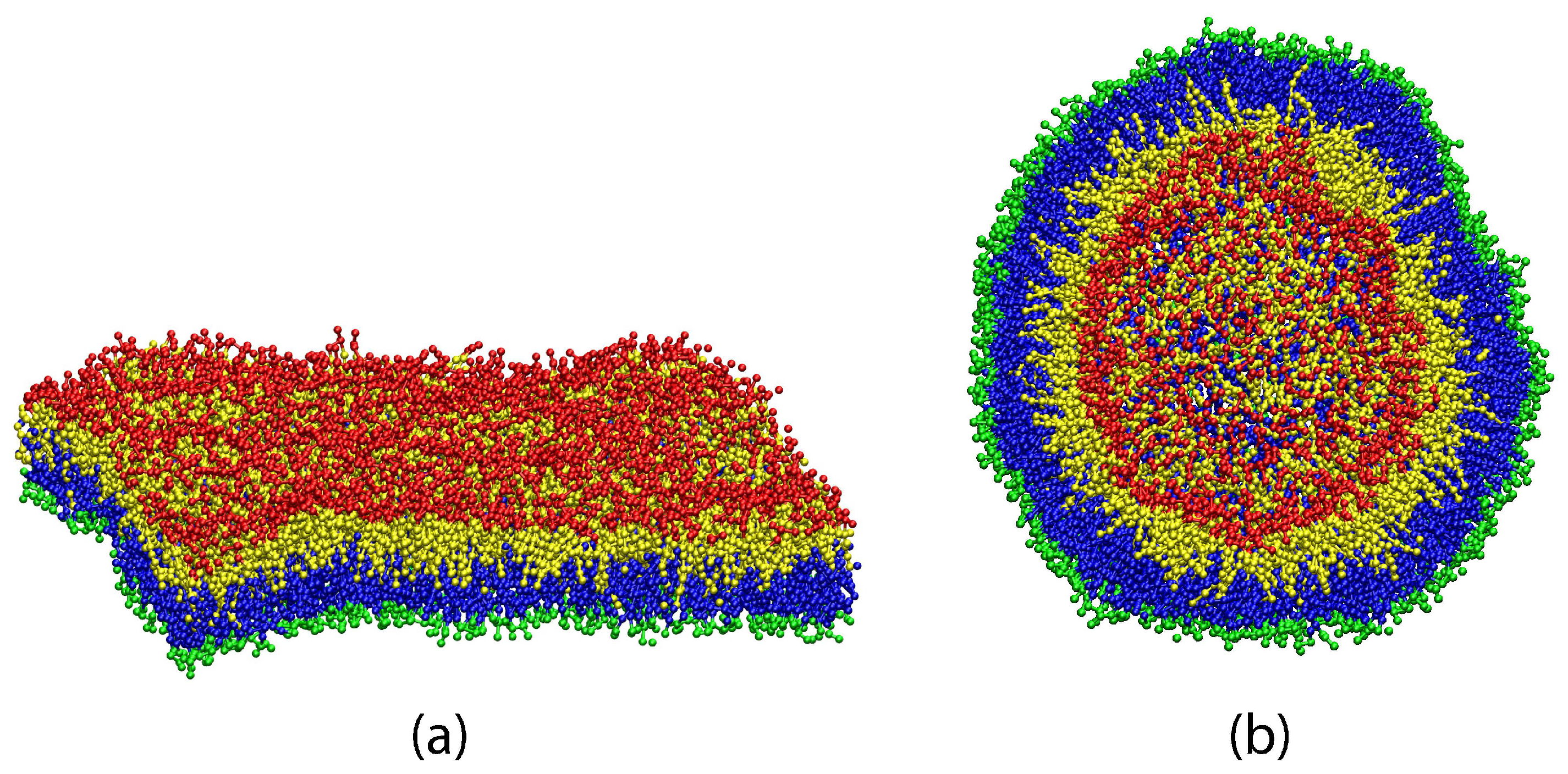{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
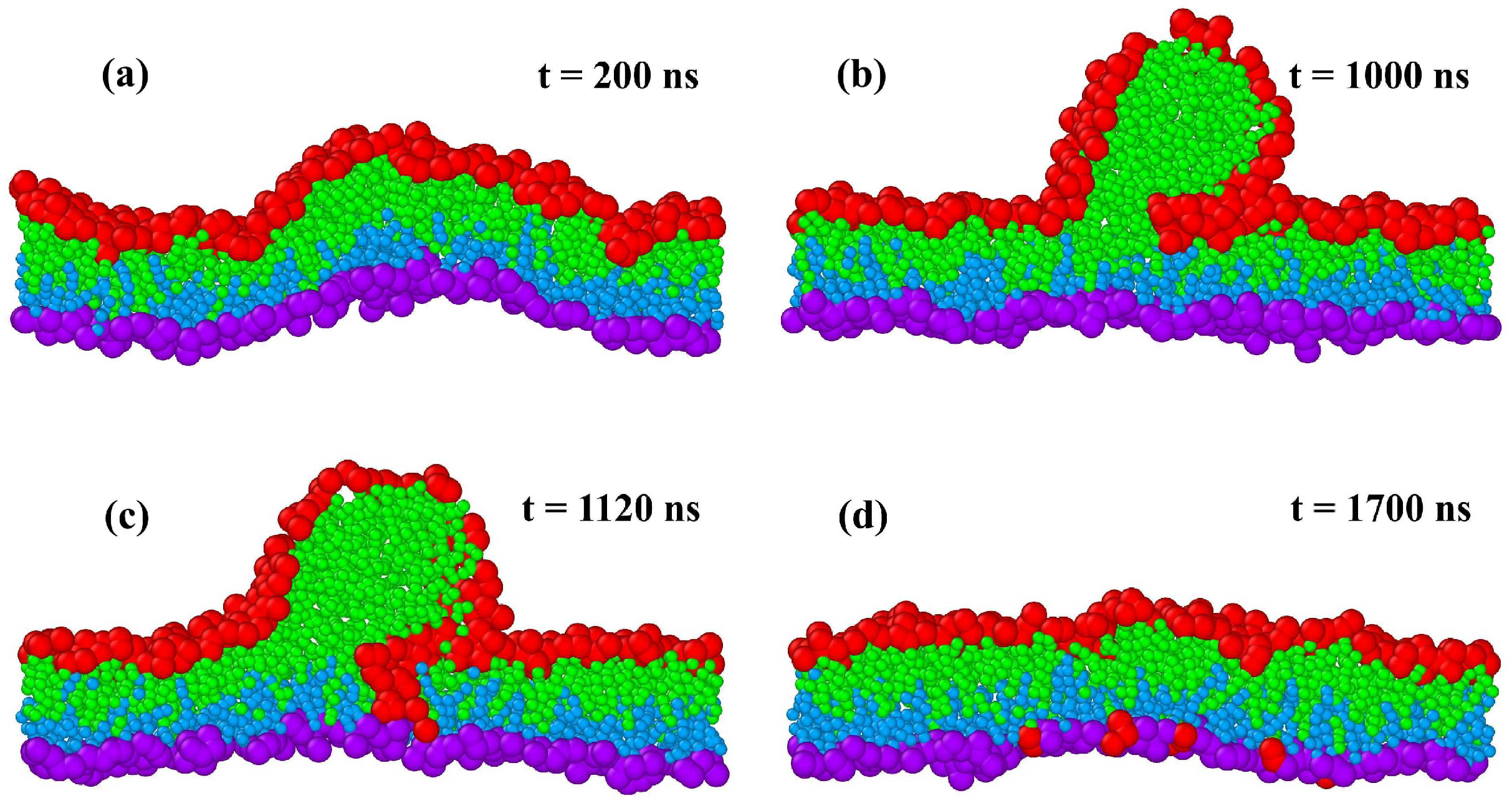{kind=link}
{kind=link}
{kind=link}
{kind=link}
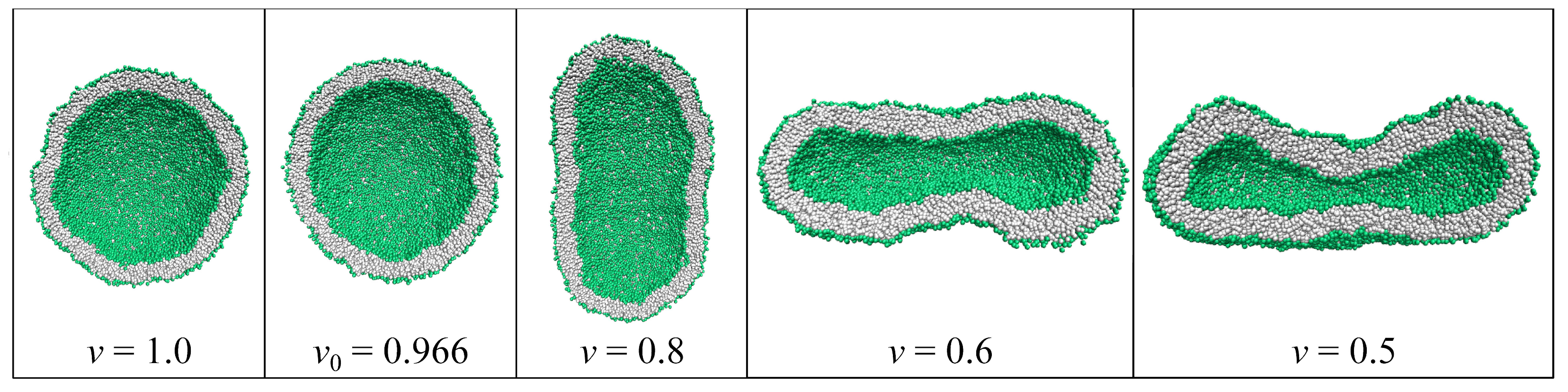{kind=link}
{kind=link}
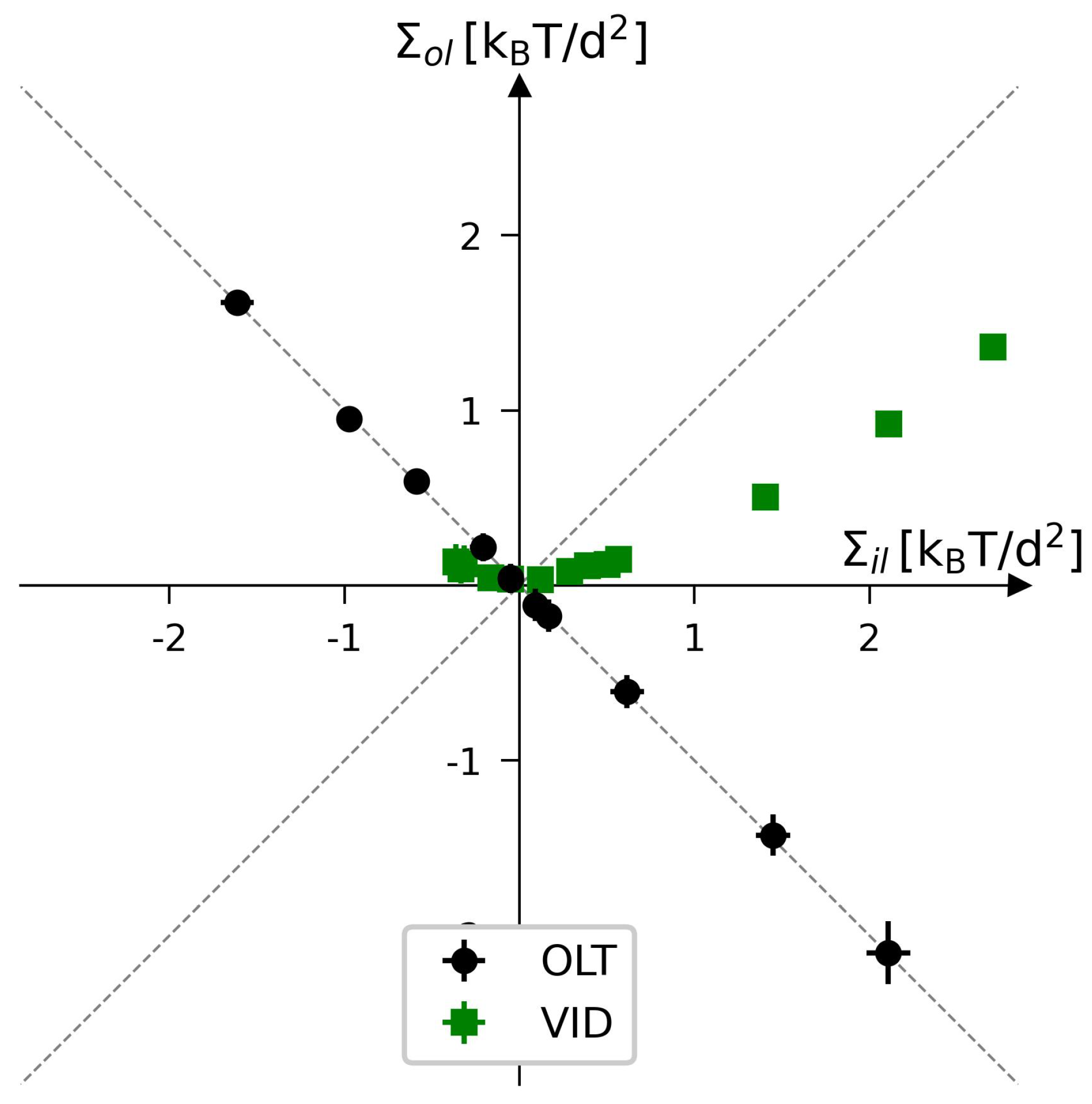{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
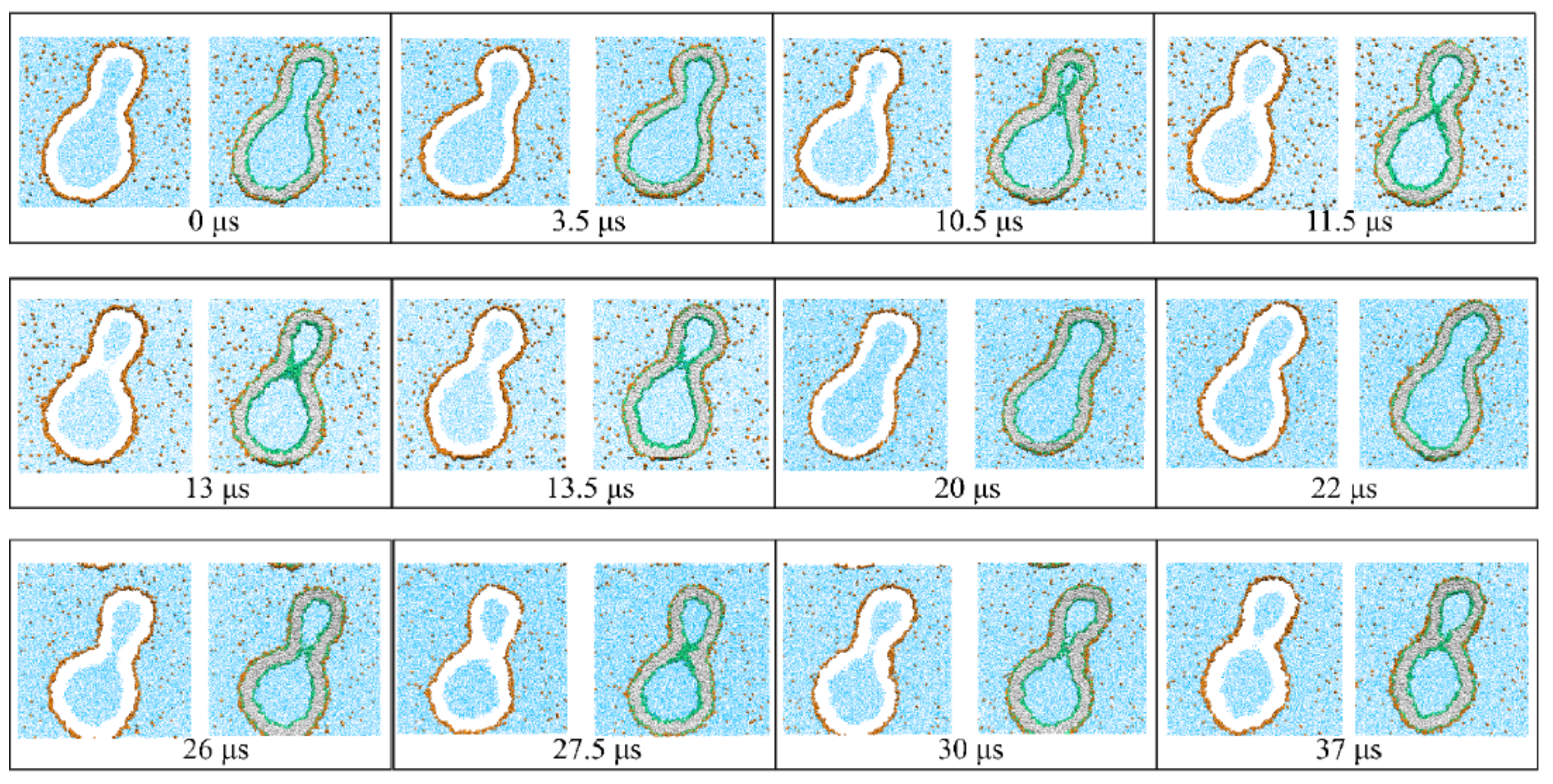{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
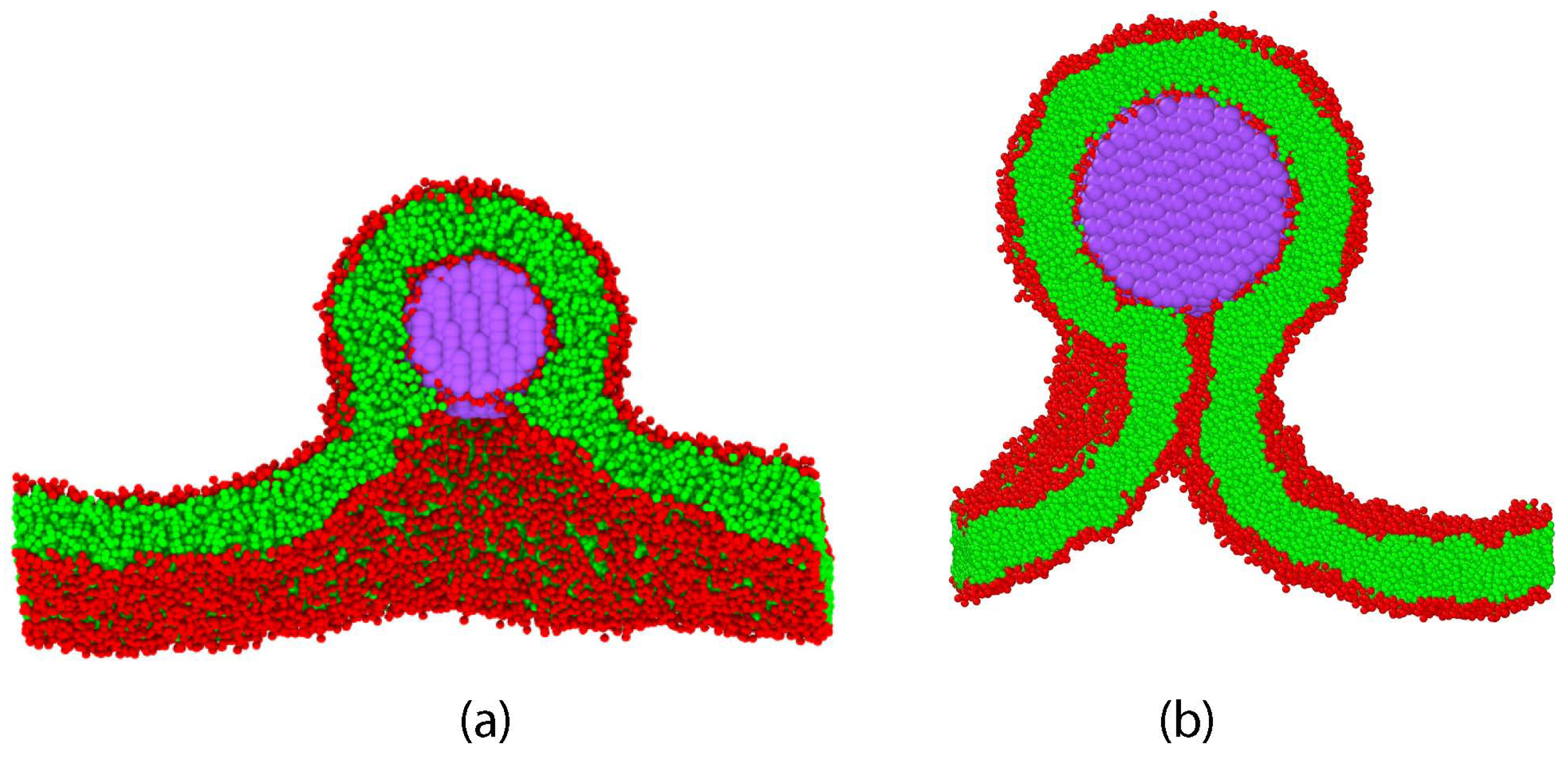{kind=link}
{kind=link}
{kind=link}
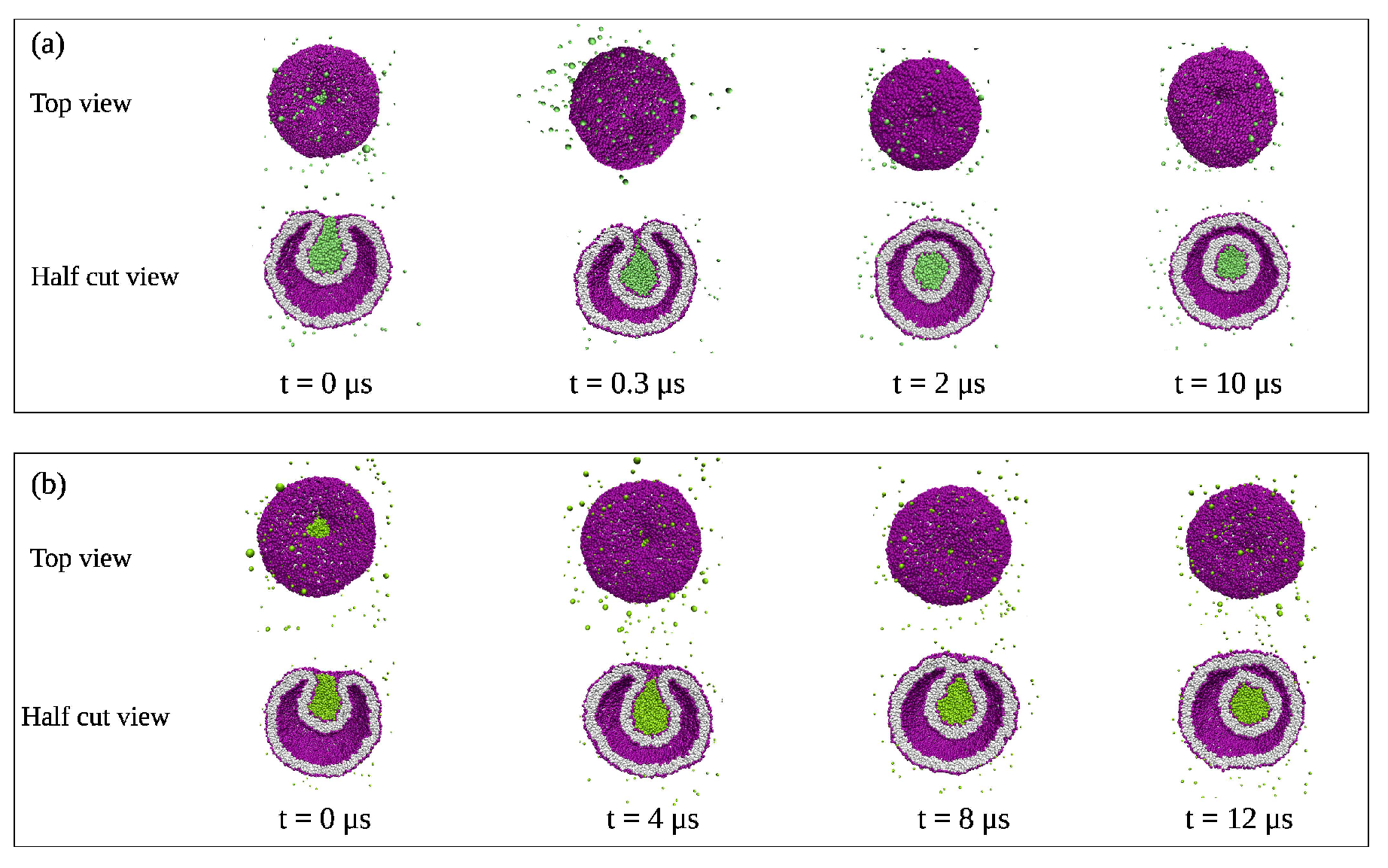{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
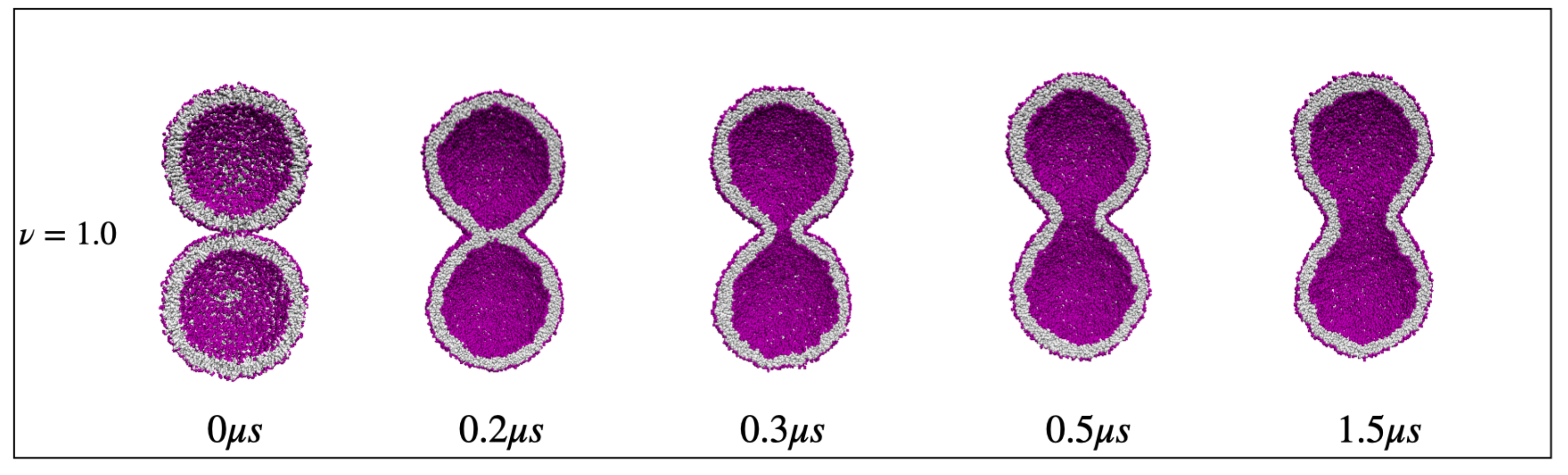{kind=link}
Abstract
:1. Introduction
2. Basic Aspects of Lipid Bilayers and Nanovesicles
2.1. Assembly and Molecular Dynamics of Lipids
2.1.1. Properties of Individual Lipid Molecules
2.1.2. Assembly and Molecular Dynamics of Lipids
2.1.3. Lateral Diffusion and Interleaflet Flip-Flops of Lipids
2.1.4. Fluid-Elastic Behavior of Molecular Bilayers and Nanovesicles
2.2. Different Notions of Membrane Tension
2.2.1. Bilayer Tension and Leaflet Tensions
2.2.2. Average Position of Leaflet-Leaflet Interface
2.2.3. Lipid Numbers and Membrane Tensions
2.3. Emergence of Membrane Curvature at the Nanoscale
2.4. Alternative Notions of Membrane Curvature
2.4.1. Intrinsic Lipid Curvature from Inverted Hexagonal Phases
2.4.2. Limitations of the Intrinsic Curvature Concept
2.4.3. Spontaneous Curvature of Bilayer Membranes
2.4.4. Spontaneous Curvature of Reference States
2.5. Dependence of Leaflet Tensions on Bilayer Geometry
2.5.1. Geometric Control Parameters for Planar Bilayers
2.5.2. Geometric Control Parameters for Vesicle Bilayers
2.6. Topological Transformations of Closed Membrane Compartments
3. Planar Bilayers with One Lipid Component
3.1. Assembly of Lipids into Planar Bilayers
3.2. Density and Stress Profiles
3.3. Bilayer Tension and Leaflet Tensions
3.4. Midplane of Leaflet-Leaflet Interface for Planar Bilayers
3.4.1. Midplane of Symmetric Planar Bilayers
3.4.2. Midplane from Density Profile of Hydrophobic Chains
3.4.3. Midplane from Volume per Lipid via Voronoi Tessellation
3.5. Two-Dimensional Leaflet Tension Space for Planar Bilayers
3.6. Area Compressibility Modulus of Symmetric Planar Bilayers
3.7. Stress Asymmetry between Leaflets of Planar Bilayers
3.8. Spontaneous Curvature of Planar Bilayers
4. Planar Bilayers with Several Lipid Components
4.1. Two Lipid Components with Small and Large Headgroups
4.1.1. Bilayer and Leaflet Compositions
4.1.2. Leaflet Tensions and Reference States
4.1.3. Small Compositional Asymmetries Generate Large Spontaneous curvatures
4.1.4. Bending Rigidity and Fluctuation Spectrum of Bending Undulations
4.2. Three-Component Bilayer with Flip-Flopping Lipid Component
4.2.1. Relaxation of Leaflet Tensions
4.2.2. Spontaneous Curvature of Two-Component Bilayers with Tensionless Leaflets
5. Instabilities of Planar Bilayers with Large Stress Asymmetries
5.1. Stability Regime for Planar Bilayers with One Lipid Component
5.2. Stress-Induced Flip-Flops of Lipids in Planar Bilayers
5.3. Stress-Induced Instability and Self-Healing of Planar Bilayers
6. Nanovesicles with One Lipid Component
6.1. Assembly of Lipids into Spherical Nanovesicles
6.2. Bilayer Tension and Leaflet Tensions of Vesicle Bilayers
6.3. Midsurface of Leaflet-Leaflet Interface for Nanovesicles
6.3.1. Midsurface from Density Profile of Hydrophobic Chains
6.3.2. Midsurface from Volume per Lipid via Voronoi Tesselation
6.4. Shape Transformations of Nanovesicles by Changes of Vesicle Volume
6.5. Fission of Membrane Neck for In-Budded Nanovesicles
6.6. Two-Dimensional Leaflet Tension Space for Nanovesicles
6.7. Stress Asymmetry of Vesicle Bilayers
6.8. Spontaneous Curvature of Vesicle Bilayers
7. Instabilities of Nanovesicles with Large Stress Asymmetries
7.1. Stability Regimes of Spherical Nanovesicles
7.2. Stress-Induced Flip-Flops between Leaflets of Nanovesicles
7.3. Stress-Induced Instability and Self-Healing of Nanovesicles
8. Remodeling of Nanovesicles via the Adsorption of Small Solutes
8.1. Phase Behavior of a Binary Solute-Water Mixture
8.2. Remodeling of Vesicle Shape for Good Solvent Conditions
8.3. Recurrent Shape Changes for Bad Solvent Conditions
8.4. Division of Nanovesicles via Solute-Mediated Adhesion
8.5. Fusion of Daughter Vesicles after Solute Removal
9. Engulfment of Condensate Droplets by Planar Bilayers
9.1. Formation of Condensate Droplets In Vitro and In Vivo
9.2. Interactions of Condensate Droplets with Biomembranes
9.3. Geometry of Partial and Complete Engulfment
9.4. Complete Engulfment of Droplets with Tight-Lipped Membrane Necks
9.5. Negative Line Tension of Contact Line
10. Engulfment of Rigid Nanoparticles by Planar Bilayers
10.1. Complete Engulfment of Nanoparticles with Tight-Lipped Membrane Necks
10.2. Other Simulation Studies of Nanoparticle Engulfment
11. Engulfment and Endocytosis of Condensate Droplets by Nanovesicles
11.1. Partial Engulfment of Condensate Droplets
11.2. Different Pathways for Engulfment and Endocytosis
11.3. Contact Line Tensions and Stress Asymmetry of Vesicle Bilayers
12. Adhesion and Fusion of Nanovesicles Controlled by Leaflet Tensions
13. Summary and Outlook
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations and Glossary of Mathematical Symbols
| C beads | chain beads |
| CHAIN | Protocol to determine the bilayer midsurface from the C bead density profile |
| DOPC | Dioleoylphosphatidylcholine, a phospholipid |
| DPPC | Dipalmitoylphosphatidylcholine, a phospholipid |
| ELT | Equal leaflet tension |
| GM1 | Monosialotetrahexosylganglioside, a glycolipid |
| GUV | Giant unilamellar vesicle |
| H beads | Headgroup beads |
| OLT | Oppositie leaflet tension |
| PEG | Polyethylene glycol |
| POPC | Palmitoyloleoylphosphatidylcholine, a phospholipid |
| S beads | Solute beads |
| STED | Simulated emission and depletion |
| SUV | Small unilamellar vesicle |
| VORON | Protocol to determine the bilayer midsurface from volume per lipid |
| W beads | water beads |
| a | Molecular area per lipid |
| a0 | Optimal molecular area per lipid |
| α, β | Two coexisting liquid phases α and β as in Figure 34 and Figure 38 |
| A‖ | Base area of cuboid simulation box |
| d | Diameter of beads in coarse-grained molecular dynamics simulations |
| ΔΣ | Stress asymmetry between two leaflets of planar bilayer, ΔΣ = Σul − Σll |
| ΔΣve | Stress asymmetry between two leaflets of vesicle bilayer, ΔΣve = Σol − Σil |
| Ddr | Diameter of spherical condensate droplet |
| kB | Boltzmann constant |
| KA | Area compressibility modulus of membrane |
| κ | Bending rigidity of membrane, which provides the basic energy scale |
| λ | Line tension of contact line between membrane and condensate droplet |
| L | Linear dimension of cuboid simulation box |
| Lz | Linear dimension of cuboid simulation box perpendicular to planar bilayer |
| m | Spontaneous (or preferred) curvature of membrane |
| M | Mean curvature of membrane surface |
| ν | Vesicle volume as in Equation (19), not to be confused with volume per lipid, υ |
| Nil | Lipid number in inner leaflet of vesicle bilayer |
| Nll | Lipid number in lower leaflet of planar bilayer |
| Nol | Lipid number in outer leaflet of vesicle bilayer |
| Nul | Lipid number in upper leaflet of planar bilayer |
| Pcdf | Cumulative distribution function for onset of flip-flops |
| Pexp | Exponential distribution function for onset of flip-flops in planar bilayers |
| PWei | Weibull distribution function for onset of flip-flops in vesicle bilayers |
| ΦS | Mole fraction of solute molecules as in Figure 27 |
| q | Wavenumber of undulation modes as in Figure 11 |
| r | Radial coordinate for spherical nanovesicle |
| rmax | Upper integration limit for integration over radial coordinate r |
| rmid | Radial coordinate for midsurface of spherical nanovesicle |
| Rco | Radius of circular contact line |
| ρC | Density profile of hydrophobic chain (C) beads |
| Σ | Bilayer tension |
| Σil | Inner leaflet tension of nanovesicle |
| Σll | Lower leaflet tension of planar bilayer |
| Σol | Outer leaflet tension of nanovesicle |
| Σul | Upper leaflet tension of planar bilayer |
| t | Time |
| T | Temperature |
| υ | Molecular volume per lipid, not to be confused with the rescaled vesicle volume ν |
| υ0 | Optimal molecular volume per lipid |
| V | Volume of vesicle |
| Vll | Volume of lower leaflet of planar bilayer |
| Vul | Volume of upper leaflet of planar bilayer |
| Vil | Volume of inner leaflet of vesicle bilayer |
| Vol | Volume of outer leaflet of vesicle bilayer |
| VlW | Volume of lower water compartment for planar bilayer |
| VuW | Volume of upper water compartment for planar bilayer |
| ViW | Volume of inner water compartment for vesicle bilayer |
| VoW | Volume of outer water compartment for vesicle bilayer |
| z | Cartesian coordinate perpendicular to planar bilayer |
| zmid | Midplane position for planar bilayer |
| ξ | Solubility of solute molecules in water as in Figure 27 |
Appendix A. Simulation Method of Dissipative Particle Dynamics
Appendix B. Stress Profiles across Lipid Bilayers
Appendix B.1. Anisotropic Pressure Tensor for Planar Bilayers
Appendix B.2. Anisotropic Pressure Tensor for Spherical Nanovesicles
References
- Gorter, E.; Grendel, F. On bimolecular layers of lipoids on the chromocytes of the blood. J. Exp. Med. 1925, 41, 439–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, J.D. The molecular structure and contact relationships of cell membranes. Prog. Biophys. Mol. Biol. 1960, 10, 343–418. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.D. The ultrastructure of cell membranes and their derivatives. Biochem. Soc. Symp. 1959, 16, 3–43. [Google Scholar] [PubMed]
- Bangham, A.; Horne, R. Negative Staining of Phospholipids and their Structural Modification by Surface-active Agents as observed in the Electron Microscope. J. Mol. Biol. 1964, 8, 660–668. [Google Scholar] [CrossRef]
- Jahn, A.; Stavis, S.M.; Hong, J.S.; Vreeland, W.N.; DeVoe, D.L.; Gaitan, M. Microfluidic Mixing and the Formation of Nanoscale Lipid Vesicles. ACS Nano 2010, 4, 2077–2087. [Google Scholar] [CrossRef]
- Pattni, B.S.; Chupin, V.V.; Torchilin, V.P. New Developments in Liposomal Drug Delivery. Chem. Rev. 2015, 115, 10938–10966. [Google Scholar] [CrossRef]
- Cevc, G.; Blume, G. Lipid vesicles penetrate into intact skin owing to the transdermal osmotic gradients and hydration force. Biochim. Biophys. Acta 1992, 1104, 226–232. [Google Scholar] [CrossRef]
- Touitou, E.; Dayan, N.; Bergelson, L.; Godin, B.; Eliaz, M. Ethosomes-novel vesicular carriers for enhanced delivery: Characterization and skin penetration properties. J. Control. Release 2000, 65, 403–418. [Google Scholar] [CrossRef]
- Bhasin, B.; Londhe, V.Y. An Overview of Transfersomal Drug Delivery. Int. J. Pharm. Sci. Res. 2018, 9, 2175–2184. [Google Scholar]
- Lai, F.; Caddeo, C.; Manca, M.L.; Manconi, M.; Sinico, C.; Fadda, A.M. What is new in the field of phospholipid vesicular nanocarriers for skin drug delivery. Int. J. Pharm. 2020, 583, 119398. [Google Scholar] [CrossRef]
- Limongi, T.; Susa, F.; Marini, M.; Allione, M.; Torre, B.; Pisano, R.; di Fabrizio, E. Lipid-Based Nanovesicular Drug Delivery Systems. J. Nanomater. 2021, 11, 3391. [Google Scholar] [CrossRef]
- Rai, S.; Pandey, V.; Rai, G. Transfersomes as versatile and flexible nano-vesicular carriers in skin cancer therapy: The state of the art. Nano Rev. Exp. 2017, 8, 1325708–1325720. [Google Scholar] [CrossRef]
- Damiani, G.; Pacifico, A.; Linder, D.M.; Pigatto, P.D.M.; Conic, R.; Grada, A.; Bragazzi, N.L. Nanodermatology-based solutions for psoriasis: State-of-the art and future prospects. Dermatol. Ther. 2019, 32, e13113. [Google Scholar] [CrossRef]
- Kulkarni, J.A.; Witzigmann, D.; Chen, S.; Cullis, P.R.; van der Meel, R. Lipid Nanoparticle Technology for Clinical Translation of siRNA Therapeutics. Acc. Chem. Res. 2019, 52, 2435–2444. [Google Scholar] [CrossRef]
- Suzuki, Y.; Ishihara, H. Difference in the lipid nanoparticle technology employed in three approved siRNA (Patisiran) and mRNA (COVID-19 vaccine) drugs. Drug Metab. Pharmacokinet. 2021, 41, 100424–100431. [Google Scholar] [CrossRef]
- Pengnam, S.; Plianwong, S.; Yingyongnarongkul, B.E.; Patrojanasophon, P.; Opanasopit, P. Delivery of small interfering RNAs by nanovesicles for cancer therapy. Drug Metab. Pharmacokinet. 2022, 42, 100425. [Google Scholar] [CrossRef]
- Chen, M.; Hu, X.; Liu, S. Next-Generation Nonviral Vectors for mRNA Vaccine Delivery. Macromol. Chem. Phys. 2022, 223, 2100443. [Google Scholar] [CrossRef]
- Ramachandran, S.; Satapathy, S.R.; Dutta, T. Delivery Strategies for mRNA Vaccines. J. Pharm. Med. 2022, 36, 11–20. [Google Scholar] [CrossRef]
- Mui, B.L.S.; Cullis, P.R.; Evans, E.A.; Madden, T.D. Osmotic properties of large unilamellar vesicles prepared by extrusion. Biophys. J. 1993, 64, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Mui, B.L.S.; Döbereiner, H.G.; Madden, T.D.; Cullis, P.R. Influence of Transbilayer Area Asymmetry on the Morphology of Large Unilamellar Vesicles. Biophys. J. 1995, 69, 930–941. [Google Scholar] [CrossRef] [Green Version]
- Frederik, P.M.; Hubert, D.H.W. Cryoelectron Microscopy of Liposomes. Meth. Enzymol. 2005, 391, 431–448. [Google Scholar]
- Dragicevic-Curic, N.; Scheglmann, D.; Albrecht, V.; Fahr, A. Temoporfin-loaded invasomes: Development, characterization and in vitro skin penetration studies. J. Control. Release 2008, 127, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Danino, D. Cryo-TEM of soft molecular assemblies. Curr. Opin. Colloid Interface Sci. 2012, 17, 316–329. [Google Scholar] [CrossRef]
- Różycki, B.; Lipowsky, R. Spontaneous curvature of bilayer membranes from molecular simulations: Asymmetric lipid densities and asymmetric adsorption. J. Chem. Phys. 2015, 142, 054101. [Google Scholar] [CrossRef] [PubMed]
- Sreekumari, A.; Lipowsky, R. Lipids with Bulky Head Groups Generate Large Membrane Curvatures by Small Compositional Asymmetries. J. Chem. Phys. 2018, 149, 084901. [Google Scholar] [CrossRef] [Green Version]
- Satarifard, V.; Grafmüller, A.; Lipowsky, R. Nanodroplets at Membranes Create Tight-Lipped Membrane Necks via Negative Line Tension. ACS Nano 2018, 12, 12424–12435. [Google Scholar] [CrossRef] [Green Version]
- Miettinen, M.; Lipowsky, R. Bilayer Membranes with Frequent Flip Flops have Tensionless Leaflets. Nano Lett. 2019, 19, 5011–5016. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, R.; Satarifard, V.; Grafmüller, A.; Lipowsky, R. Spherical Nanovesicles Transform into a Multitude of Nonspherical Shapes. Nano Lett. 2019, 19, 7703–7711. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, R.; Satarifard, V.; Grafmüller, A.; Lipowsky, R. Budding and Fission of Nanovesicles induced by membrane adsorption of small solutes. ACS Nano 2021, 15, 7237–7248. [Google Scholar] [CrossRef]
- Sreekumari, A.; Lipowsky, R. Large stress asymmetries of lipid bilayers and nanovesicles generate lipid flip-flops and bilayer instabilities. Soft Matter 2022, 18, 6066–6078. [Google Scholar] [CrossRef]
- Ghosh, R.; Satarifard, V.; Lipowsky, R. Different pathways for engulfment and endocytosis of droplets by nanovesicles. Nat. Commun. 2023, 14, 615. [Google Scholar] [CrossRef]
- Zamaletdinov, M.; Miettinen, M.; Lipowsky, R. Probing the elastic response of lipid bilayers and nanovesicles to leaflet tensions via volume per lipid. Soft Matter. (under review).
- Hossein, A.; Deserno, M. Spontaneous Curvature, Differential Stress, and Bending Modulus of Asymmetric Lipid Membranes. Biophys. J. 2020, 118, 624–642. [Google Scholar] [CrossRef]
- Varma, M.; Deserno, M. Distribution of cholesterol in asymmetric membranes driven by composition and differential stress. Biophys. J. 2022, 121, 4001–4018. [Google Scholar] [CrossRef]
- Goetz, R.; Lipowsky, R. Computer simulations of bilayer membranes: Self-assembly and interfacial tension. J. Chem. Phys. 1998, 108, 7397–7409. [Google Scholar] [CrossRef] [Green Version]
- Goetz, R.; Gompper, G.; Lipowsky, R. Mobilitiy and elasticity of self-assembled membranes. Phys. Rev. Lett. 1999, 82, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Hoogebrugge, P.; Koelman, J. Simulating nicroscopic hydrodynamic phenomena with dissipative particle dynamics. Europhys. Lett. 1992, 19, 155–160. [Google Scholar] [CrossRef]
- Español, P.; Warren, P. Statistical Mechanics of Dissipative Particle Dynamics. Europhys. Lett. 1995, 19, 190–196. [Google Scholar] [CrossRef] [Green Version]
- Groot, R.D.; Warren, P.B. Dissipative particle dynamics: Bridging the gap between atomistic and mesoscopic simulation. J. Chem. Phys. 1997, 107, 4423–4435. [Google Scholar] [CrossRef] [Green Version]
- Shillcock, J.C.; Lipowsky, R. Equilibrium structure and lateral stress distribution of amphiphilic bilayers from dissipative particle dynamics simulations. J. Chem. Phys. 2002, 117, 5048–5061. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.P.; Aktulga, H.M.; Berger, R.; Bolintineanu, D.S.; Brown, W.M.; Crozier, P.S.; in ’t Veld, P.J.; Kohlmeyer, A.; Moore, S.G.; Nguyen, T.D.; et al. LAMMPS-a flexible simulation tool for particle-based materials modeling at the atomic, meso, and continuum scales. Comput. Phys. Commun. 2022, 271, 108171. [Google Scholar] [CrossRef]
- Nakamura, T.; Kawamoto, S.; Shinoda, W. Precise Calculation of the Local Pressure Tensor in Cartesian and Spherical Coordinates in LAMMPS. Comput. Phys. Commun. 2015, 190, 120–128. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Marrink, S.J.; Risselada, H.J.; Yefimov, S.; Tieleman, D.P.; de Vries, A.H. The MARTINI Force Field: Coarse Grained Model for Biomolecular Simulations. J. Phys. Chem. B 2007, 111, 7812–7824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shillcock, J.; Lipowsky, R. Tension-induced fusion of bilayer membranes and vesicles. Nat. Mater. 2005, 4, 225–228. [Google Scholar] [CrossRef] [Green Version]
- Grafmüller, A.; Shillcock, J.; Lipowsky, R. Pathway of Membrane Fusion with Two Tension-Dependent Energy Barriers. Phys. Rev. Lett. 2007, 98, 218101. [Google Scholar] [CrossRef] [Green Version]
- Lipowsky, R. Remodeling of Membrane Shape and Topology by Curvature Elasticity and Membrane Tension. Adv. Biol. 2022, 6, 2101020. [Google Scholar] [CrossRef]
- Ortiz, V.; Nielsen, S.O.; Discher, D.E.; Klein, M.L.; Lipowsky, R.; Shillcock, J. Dissipative Particle Dynamics Simulations of Polymersomes. J. Phys. Chem. B 2005, 109, 17708–17714. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Li, X.; Gao, L.; Fang, W. Theoretical Insight into the Relationship between the Structures of Antimicrobial Peptides and Their Actions on Bacterial Membranes. J. Phys. Chem. B 2015, 110, 850–860. [Google Scholar] [CrossRef]
- Choudhury, C.K.; Kuksenok, O. Native-Based Dissipative Particle Dynamics Approach for α-Helical Folding. J. Phys. Chem. B 2020, 124, 11379–11386. [Google Scholar] [CrossRef]
- Lavagnini, E.; Cook, J.L.; Warren, P.B.; Hunter, C.A. Translation of Chemical Structure into Dissipative Particle Dynamics Parameters for Simulation of Surfactant Self-Assembly. J. Phys. Chem. B 2021, 125, 3942–3952. [Google Scholar] [CrossRef] [PubMed]
- Sgouros, A.P.; Knippenberg, S.; Guillaume, M.; Theodorou, D.N. Multiscale simulations of polyzwitterions in aqueous bulk solutions and brush array configurations. Soft Matter 2021, 17, 10873–10890. [Google Scholar] [CrossRef] [PubMed]
- Vaiwala, R.; Ayappa, K.G. A generic force field for simulating native protein structures using dissipative particle dynamics. Soft Matter 2021, 17, 9772–9785. [Google Scholar] [CrossRef] [PubMed]
- Marsh, D. Molecular volumes of phospholipids and glycolipids in membranes. Chem. Phys. Lipids 2010, 163, 667–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberts, B.; Bray, D.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Essential Cell Biology: An Introduction to the Molecular Biology of the Cell; Garland: New York, NY, USA, 1998. [Google Scholar]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Woith, E.; Fuhrmann, G.; Melzig, M.F. Extracellular Vesicles–Connecting Kingdoms. Int. J. Mol. Sci. 2019, 20, 5695. [Google Scholar] [CrossRef] [Green Version]
- Barile, L.; Vassalli, G. Exosomes: Therapy delivery tools and biomarkers of diseases. Pharmacol. Ther. 2017, 174, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Jalaludin, I.; Lubman, D.M.; Kim, J. A guide to mass spectrometric analysis of extracellular vesicle proteins for biomarker discovery. Mass Spec. Rev. 2021, 42, e21749. [Google Scholar] [CrossRef]
- Herrmann, I.K.; Wood, M.J.A.; Fuhrmann, G. Extracellular vesicles as a next-generation drug delivery platform. Nat. Nanotechnol. 2021, 16, 748–759. [Google Scholar] [CrossRef]
- Van der Koog, L.; Gandek, T.B.; Nagelkerke, A. Liposomes and Extracellular Vesicles as Drug Delivery Systems: A Comparison of Composition, Pharmacokinetics, and Functionalization. Adv. Healthc. Mater. 2022, 11, 2100639. [Google Scholar] [CrossRef]
- Kornberg, R.D.; McConnell, H.M. Inside-outside transitions of phospholipids in vesicle membranes. Biochemistry 1971, 10, 1111–1120. [Google Scholar]
- Devaux, P.; McConnell, H.M. Lateral Diffusion in Spin-Labeled Phosphatidylcholine Multilayers. J. Am. Chem. Soc. 1972, 94, 4475–4481. [Google Scholar] [CrossRef]
- Sackmann, E.; Träuble, H. Studies of the Crystalline-Liquid Crystalline Phase Transition of Lipid Model Membranes II: Analysis of Electron Spin Resonance Spectra of Steroid Labels Incorporated into Lipid Membranes. J. Am. Chem. Soc. 1972, 94, 4492–4498. [Google Scholar] [CrossRef]
- Träuble, H.; Sackmann, E. Studies of the Crystalline-Liquid Crystalline Phase Transition of Lipid Model Membranes III: Structure of a Steroid-Lecithin System below and above the Lipid-Phase Transition. J. Am. Chem. Soc. 1972, 94, 4499–4510. [Google Scholar] [CrossRef]
- Almeida, P.F.F.; Vaz, W.L.C. Lateral Diffusion in Membranes. In Structure and Dynamics of Membranes; Lipowsky, R., Sackmann, E., Eds.; Elsevier: Amsterdam, The Netherlands, 1995; Volume 1A. [Google Scholar]
- Sako, Y.; Kusumi, A. Compartmentalized Structure of the Plasma Membrane for Receptor Movements as Revealed by a Nanometer-level Motion Analysis. J. Cell Biol. 1994, 125, 1251–1264. [Google Scholar] [CrossRef]
- Saxton, M.J.; Jacobson, K. Single-Particle Tracking: Applications to Membrane Dynamics. Annu. Rev. Biophys. Biomol. Struct. 1997, 26, 373–399. [Google Scholar] [CrossRef]
- Fujiwara, T.; Ritchie, K.; Murakoshi, H.; Jacobson, K.; Kusumi, A. Phospholipids undergo hop diffusion in compartmentalized cell membrane. J. Cell Biol. 2002, 157, 1071–1081. [Google Scholar] [CrossRef] [Green Version]
- Kusumi, A.; Nakada, C.; Ritchie, K.; Murase, K.; Suzuki, K.; Murakoshi, H.; Kasai, R.S.; Kondo, J.; Fujiwara, T. Paradigm Shift of the Plasma Membrane Concept from the Two-Dimensional Continuum Fluid to the Partitioned Fluid: High-Speed Single-Molecule Tracking of Membrane Molecules. Annu. Rev. Biophys. Biomol. Struct. 2005, 34, 351–378. [Google Scholar] [CrossRef] [Green Version]
- Van der Meer, B.W. Fluidity, Dynamics, and Order. In Biomembranes: Physical Aspects; Shinitzky, M., Ed.; VCH Verlagsgesellschaft: Weinheim, Germany, 1993; pp. 97–158. [Google Scholar]
- Victorov, A.V.; Janes, N.; Moehren, G.; Rubin, E.; Taraschi, T.F.; Hoek, J.B. Rapid Transbilayer Movement of Phosphatidylethanol in Unilamellar Phosphatidylcholine Vesicles. J. Am. Chem. Soc. 1994, 116, 4050–4052. [Google Scholar] [CrossRef]
- Liu, J.; Conboy, J.C. 1,2-Diacyl-Phosphatidylcholine Flip-Flop Measured Directly by Sum-Frequency Vibrational Spectroscopy. Biophys. J. 2005, 89, 2522–2532. [Google Scholar] [CrossRef] [Green Version]
- Nakano, M.; Fukuda, M.; Kudo, T.; Matsuzaki, N.; Azuma, T.; Sekine, K.; Endo, H.; Handa, T. Flip-flop of phospholipids in vesicles: Kinetic analysis with time-resolved small-angle neutron scattering. J. Phys. Chem. B 2009, 113, 6745–6748. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Brown, K.L.; Conboy, J.C. The effect of cholesterol on the intrinsic rate of lipid flip-flop as measured by sum-frequency vibrational spectroscopy. Faraday Discuss. 2013, 161, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, D.; Geier, B.; Pabst, G. Asymmetric Lipid Membranes: Towards More Realistic Model Systems. Membranes 2015, 5, 180–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquardt, D.; Heberle, F.A.; Miti, T.; Eicher, B.; London, E.; Katsaras, J.; Pabst, G. 1H NMR shows slow phospholipid flip-flop in gel and fluid bilayers. Langmuir 2017, 33, 3731–3741. [Google Scholar] [CrossRef]
- Sperotto, M.M.; Ferrarini, A. Spontaneous lipid flip-flop in membranes: A still unsettled picture from experiments and simulations. In The Biophysics of Cell Membranes; Springer Series in Biophysics 19; Epand, R., Ruysschaert, J.M., Eds.; Springer Nature: Singapore, 2017; pp. 29–60. [Google Scholar]
- Porcar, L.; Gerelli, Y. On the lipid flip-flop and phase transition coupling. Soft Matter 2020, 16, 7696–7703. [Google Scholar] [CrossRef]
- Steck, T.L.; Ye, J.; Lange, Y. Probing Red Cell Membrane Cholesterol Movement with Cyclodextrin. Biophys. J. 2002, 83, 2118–2125. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, J.A. Fast flip-flop of cholesterol and fatty acids in membranes: Implications for membrane transport proteins. Curr. Opin. Lipidol. 2003, 14, 263–271. [Google Scholar] [CrossRef]
- Bruckner, R.J.; Mansy, S.S.; Ricardo, A.; Mahadevan, L.; Szostak, J.W. Flip-Flop-Induced Relaxation of Bending Energy: Implications for Membrane Remodeling. Biophys. J. 2009, 97, 3113–3122. [Google Scholar] [CrossRef] [Green Version]
- Evans, E.; Needham, D. Physical Properties of Surfactant Bilayer Membranes: Thermal Transitions, Elasticity, Rigidity, Cohesion, and Colloidal Interactions. J. Phys. Chem. 1987, 91, 4219–4228. [Google Scholar] [CrossRef]
- Rawicz, W.; Olbrich, K.C.; McIntosh, T.; Needham, D.; Evans, E. Effect of Chain Length and Unsaturation on Elasticity of Lipid Bilayers. Biophys. J. 2000, 79, 328–339. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, T.; Agudo-Canalejo, J.; Dimova, R.; Lipowsky, R. Membrane Nanotubes Increase the Robustness of Giant Vesicles. ACS Nano 2018, 12, 4478–4485. [Google Scholar] [CrossRef] [Green Version]
- Callen, H. Thermodynamics and an Introduction to Thermostatistics, 2nd ed.; Wiley: New York, NY, USA, 1985. [Google Scholar]
- Israelachvili, J.; Marcelja, S.; Horn, R. Physical principles of membrane organization. Quart. Rev. Biophys. 1980, 13, 121–200. [Google Scholar] [CrossRef] [Green Version]
- Sadoc, J.F.; Charvolin, J. Frustration in bilayers and topologies of liquid crystals of amphiphilic molecules. J. Phys. 1986, 47, 683–691. [Google Scholar] [CrossRef]
- Semeraro, E.F.; Marx, L.; Frewein, M.P.K.; Pabst, G. Increasing complexity in small-angle X-ray and neutron scattering experiments: From biological membrane mimics to live cells. Soft Matter 2020, 17, 222–232. [Google Scholar] [CrossRef] [Green Version]
- Seddon, J.M.; Templer, R.H. Polymorphism of Lipid-Water Systems. In Structure and Dynamics of Membranes; Lipowsky, R., Sackmann, E., Eds.; Elsevier Science: Amsterdam, The Netherlands, 1995; Chapter 3; Volume 1A, pp. 97–160. [Google Scholar]
- Helfrich, W. Elastic properties of lipid bilayers: Theory and possible experiments. Z. Naturforsch. C 1973, 28, 693–703. [Google Scholar] [CrossRef]
- Frank, F.C. I. Liquid Crystals. On the Theory of Liquid Crystals. Discuss. Faraday Soc. 1958, 25, 19–28. [Google Scholar] [CrossRef]
- Seifert, U.; Berndl, K.; Lipowsky, R. Shape transformations of vesicles: Phase diagram for spontaneous curvature and bilayer coupling model. Phys. Rev. A 1991, 44, 1182–1202. [Google Scholar] [CrossRef] [Green Version]
- Lipowsky, R. Understanding giant vesicles: A theoretical perspective. In The Giant Vesicle Book; Dimova, R., Marques, C., Eds.; CRC Press: Boca Raton, FL, USA, 2019; Chapter 5; pp. 73–168. [Google Scholar]
- Bhatia, T.; Christ, S.; Steinkühler, J.; Dimova, R.; Lipowsky, R. Simple sugars shape giant vesicles into multispheres with many membrane necks. Soft Matter 2020, 16, 1246–1258. [Google Scholar] [CrossRef] [Green Version]
- Steinkühler, J.; Knorr, R.L.; Zhao, Z.; Bhatia, T.; Bartelt, S.; Wegner, S.; Dimova, R.; Lipowsky, R. Controlled division of cell-sized vesicles by low densities of membrane-bound proteins. Nat. Commun. 2020, 11, 905. [Google Scholar] [CrossRef] [Green Version]
- Seifert, U.; Miao, L.; Döbereiner, H.G.; Wortis, M. Budding transition for bilayer fluid vesicles. In The Structure and Conformation of Amphiphilic Membranes; Lipowsky, R., Richter, D., Kremer, K., Eds.; Springer Proceedings in Physic; Springer: Berlin/Heidelberg, Germany, 1992; pp. 93–96. [Google Scholar]
- Döbereiner, H.G.; Evans, E.; Kraus, M.; Seifert, U.; Wortis, M. Mapping vesicle shapes into the phase diagram: A comparison of experiment and theory. Phys. Rev. E 1997, 55, 4458–4474. [Google Scholar] [CrossRef] [Green Version]
- Svetina, S.; Žekš, B. Shape Behavior of Lipid Vesicles as the Basis of Some Cellular Processes. Anat. Rec. 2002, 268, 215–225. [Google Scholar] [CrossRef] [PubMed]
- do Carmo, M. Differential Geometry of Curves and Surfaces; Prentice-Hall: Englewood Cliffs, NJ, USA, 1976. [Google Scholar]
- Helfrich, W. Amphiphilic mesophases made of defects. In Physics of Defects; Balian, R., Kleman, M., Poirier, J.P., Eds.; North-Holland Publishing Company: Amsterdam, The Netherlands, 1981; pp. 715–755. [Google Scholar]
- Lipowsky, R. Understanding and controlling the morphological complexity of biomembranes. In Multiresponsive Behavior of Biomembranes and Giant Vesicles; Academic Press: London, UK, 2019; Chapter 3; Volume 30, pp. 105–155. [Google Scholar]
- Dasgupta, R.; Miettinen, M.S.; Fricke, N.; Lipowsky, R.; Dimova, R. The glycolipid GM1 reshapes asymmetric biomembranes and giant vesicles by curvature generation. Proc. Nat. Acad. Sci. USA 2018, 115, 5756–5761. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Agudo-Canalejo, J.; Grafmüller, A.; Dimova, R.; Lipowsky, R. Patterns of flexible nanotubes formed by liquid-ordered and liquid-disordered membranes. ACS Nano 2016, 10, 463–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagle, J.F. Introductory Lecture: Basic quantities in model biomembranes. Faraday Discuss. 2013, 161, 11–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bancroft, W.D. The theory of emulsification, V. J. Phys. Chem. 1913, 17, 501–519. [Google Scholar] [CrossRef] [Green Version]
- Bancroft, W.; Tucker, C. Gibbs on emulsification. J. Phys. Chem. 1927, 31, 1681–1692. [Google Scholar] [CrossRef]
- Lipowsky, R. Remodeling of Biomembranes and Vesicles by Adhesion of Condensate Droplets. Membranes 2023, 13, 223. [Google Scholar] [CrossRef]
- Weibull, W. A Statistical Distribution Function of Wide Applicability. J. Appl. Mech. 1951, 18, 293–297. [Google Scholar] [CrossRef]
- Rinne, H. The Weibull Distribution—A Handbook; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Lai, C.D. Generalized Weibull Distributions; Springer Briefs in Statistics; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Cox, D.R. Renewal Theory; Methuen and Co., Ltd.: London, UK, 1962. [Google Scholar]
- Taylor, H.M.; Karlin, S. An Introduction to Stochastic Modelling; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Fesce, R.; Grohovaz, F.; Valtorta, F.; Meldolesi, J. Neurotransmitter Release: Fusion or ‘Kiss-and-run’? Trends Cell Biol. 1994, 4, 1–4. [Google Scholar] [CrossRef]
- Staal, R.G.W.; Mosharov, E.V.; Sulzer, D. Dopamine Neurons Release Transmitter via a Flickering Fusion Pore. Nat. Neurosci. 2004, 7, 341–346. [Google Scholar] [CrossRef]
- Wightman, R.M.; Haynes, C.L. Synaptic Vesicles Really Do Kiss and Run. Nat. Neurosci. 2004, 7, 321–322. [Google Scholar] [CrossRef]
- Albertsson, P.A. Partition of Cell Particles and Macromolecules: Separation and Purification of Biomolecules, Cell Organelles Membranes, and Cells in Aqueous Polymer Two-Phase Systems and Their Use in Biochemical Analysis and Biotechnology, 3rd ed.; Wiley: New York, NY, USA, 1986. [Google Scholar]
- Esquena, J. Water-in-water (W/W) emulsions. Curr. Opin. Colloid Interface Sci. 2016, 25, 109–119. [Google Scholar] [CrossRef]
- Crowe, C.D.; Keating, C.D. Liquid–liquid phase separation in artificial cells. Interface Focus 2018, 8, 20180032. [Google Scholar] [CrossRef] [Green Version]
- De Jong, H.G.B.; Kruyt, H.R. Coacervation (Partial miscibility in colloid systems)(Preliminary Communication). Proc. K. Ned. Akad. Wet. 1929, 32, 849–856. [Google Scholar]
- Sing, C.E. Development of the modern theory of polymeric complex coacervation. Adv. Colloid Interface Sci. 2017, 239, 2–16. [Google Scholar] [CrossRef]
- Li, L.; Srivastava, S.; Andreev, M.; Marciel, A.B.; de Pablo, J.J.; Tirrell, M.V. Phase Behavior and Salt Partitioning in Polyelectrolyte Complex Coacervates. Macromolecules 2018, 51, 2988–2995. [Google Scholar] [CrossRef]
- Brangwynne, C.P.; Eckmann, C.R.; Courson, D.S.; Rybarska, A.; Hoege, C.; Gharakhani, J.; Jülicher, F.; Hyman, A.A. Germline P Granules Are Liquid Droplets That Localize by Controlled Dissolution/Condensation. Science 2009, 324, 1729–1732. [Google Scholar] [CrossRef]
- Fritsch, A.W.; Diaz-Delgadillob, A.F.; Adame-Arana, O.; Hoege, C.; Mittasch, M.; Kreysing, M.; Leaver, M.; Hyman, A.A.; Jülicher, F.; Weber, C.A. Local thermodynamics govern formation and dissolution of Caenorhabditis elegans P granule condensates. Proc. Natl. Acad. Sci. USA 2021, 118, e2102772118. [Google Scholar] [CrossRef]
- Berry, J.; Weber, S.C.; Vaidya, N.; Haatajaa, M.; Brangwynne, C.P. RNA transcription modulates phase transition-driven nuclear body assembly. Proc. Natl. Acad. Sci. USA 2015, 112, E5237–E5245. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Wheeler, J.R.; Walters, R.W.; Agrawal, A.; Barsic, A.; Parker, R. ATPase-Modulated Stress Granules Contain a Diverse Proteome and Substructure. Cell 2016, 164, 487–498. [Google Scholar] [CrossRef] [Green Version]
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular condensates: Organizers of cellular biochemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Banjade, S.; Cheng, H.C.; Kim, S.; Chen, B.; Guo, L.; Llaguno, M.; Hollingsworth, J.V.; King, D.S.; Banani, S.F.; et al. Phase transitions in the assembly of multivalent signalling proteins. Nature 2012, 483, 336–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, A.; Lee, H.O.; Jawerth, L.; Maharana, S.; Jahnel, M.; Hein, M.Y.; Stoynov, S.; Mahamid, J.; Saha, S.; Franzmann, T.M.; et al. A Liquid-to-Solid Phase Transition of the ALS Protein FUS Accelerated by Disease Mutation. Cell 2015, 162, 1066–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Protter, D.S.W.; Rosen, M.K.; Parker, R. Formation and Maturation of Phase-Separated Liquid Droplets by RNA-Binding Proteins. Mol. Cell 2015, 60, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Molliex, A.; Temirov, J.; Lee, J.; Kim, H.J.; Mittag, T.; Taylor, J.P. Phase Separation by Low Complexity Domains Promotes Stress Granule Assembly and Drives Pathological Fibrillization. Cell 2015, 163, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Han, T.W.; Kato, M.; Xie, S.; Wu, L.C.; Mirzaei, H.; Pei, J.; Chen, M.; Xie, Y.; Allen, J.; Xiao, G.; et al. Cell-free Formation of RNA Granules: Bound RNAs Identify Features and Components of Cellular Assemblies. Cell 2012, 149, 768–779. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Shorter, J. It’s Raining Liquids: RNA Tunes Viscoelasticity and Dynamics of Membraneless Organelles. Mol. Cell 2015, 60, 189–192. [Google Scholar] [CrossRef] [Green Version]
- Helfrich, M.; Mangeney-Slavin, L.; Long, M.; Djoko, K.; Keating, C. Aqueous Phase Separation in Giant Vesicles. J. Am. Chem. Soc. 2002, 124, 13374–13375. [Google Scholar] [CrossRef]
- Li, Y.; Lipowsky, R.; Dimova, R. Transition from complete to partial wetting within membrane compartments. J. Am. Chem. Soc. 2008, 130, 12252–12253. [Google Scholar] [CrossRef] [Green Version]
- Kusumaatmaja, H.; Li, Y.; Dimova, R.; Lipowsky, R. Intrinsic contact angle of aqueous phases at membranes and vesicles. Phys. Rev. Lett. 2009, 103, 238103. [Google Scholar] [CrossRef] [Green Version]
- Andes-Koback, M.; Keating, C.D. Complete budding and asymmetric division of primitive model cells to produce daughter vesicles with different interior and membrane compositions. J. Am. Chem. Soc. 2011, 133, 9545–9555. [Google Scholar] [CrossRef]
- Li, Y.; Kusumaatmaja, H.; Lipowsky, R.; Dimova, R. Wetting-induced budding of vesicles in contact with several aqueous phases. J. Phys. Chem. B 2012, 116, 1819–1823. [Google Scholar] [CrossRef]
- Love, C.; Steinkühler, J.; Gonzales, D.T.; Yandrapalli, N.; Robinson, T.; Dimova, R.; Tang, T.Y.D. Reversible pH-Responsive Coacervate Formation in Lipid Vesicles Activates Dormant Enzymatic Reactions. Angew. Chem. 2020, 132, 6006–6013. [Google Scholar] [CrossRef]
- Last, M.G.F.; Deshpande, S.; Dekker, C. pH-Controlled Coacervate-Membrane Interactions within Liposomes. ACS Nano 2020, 14, 4487–4498. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, Y.; Yang, X.; He, X.; Li, M.; Liu, S.; Wang, K.; Liu, J.; Mann, S. Giant Coacervate Vesicles As an Integrated Approach to Cytomimetic Modeling. J. Am. Chem. Soc. 2021, 143, 2866–2874. [Google Scholar] [CrossRef]
- Cakmak, F.P.; Marianelli, A.M.; Keating, C.D. Phospholipid Membrane Formation Templated by Coacervate Droplets. Langmuir 2021, 37, 10366–10375. [Google Scholar] [CrossRef]
- Lu, T.; Liese, S.; Schoenmakers, L.; Weber, C.A.; Suzuki, H.; Huck, W.T.S.; Spruijt, E. Endocytosis of Coacervates into Liposomes. J. Am. Chem. Soc. 2022, 144, 13451–13455. [Google Scholar] [CrossRef]
- Milovanovic, D.; Wu, Y.; Bian, X.; Camilli, P.D. A liquid phase of synapsin and lipid vesicles. Science 2018, 361, 604–607. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Mayr, C. A Membraneless Organelle Associated with the Endoplasmic Reticulum Enables 3’UTR-Mediated Protein-Protein Interactions. Cell 2018, 175, 1492–1506. [Google Scholar] [CrossRef] [Green Version]
- Bergeron-Sandoval, L.P.; Kumar, S.; Heris, H.K.; Chang, C.; Cornell, C.E.; Keller, S.L.; François, P.; Hendricks, A.; Ehrlicher, A.J.; Pappu, R.V.; et al. Proteins with prion-like domains form viscoelastic condensates that enable membrane remodeling and endocytosis. Proc. Nat. Acad. Sci. USA 2021, 118, e2113789118. [Google Scholar] [CrossRef]
- Day, K.J.; Kago, G.; Wang, L.; Richter, J.B.; Hayden, C.C.; Lafer, E.M.; Stachowiak, J.C. Liquid-like protein interactions catalyse assembly of endocytic vesicles. Nat. Cell Biol. 2021, 23, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Witkowska, A.; Haucke, V. Liquid-like protein assemblies initiate endocytosis. Nat. Cell Biol. 2021, 23, 301–302. [Google Scholar] [CrossRef]
- Snead, W.T.; Jalihal, A.P.; Gerbich, T.M.; Seim, I.; Hu, Z.; Gladfelter, A.S. Membrane surfaces regulate assembly of ribonucleoprotein condensates. Nat. Cell Biol. 2022, 24, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.A.; Jasnow, D.; Balazs, A.C. Designing Synthetic Vesicles that Engulf Nanoscopic Particles. J. Chem. Phys. 2007, 127, 084703. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, H.; Takasu, M. Adhesion of Nanoparticles to Vesicles: A Brownian Dynamics Simulation. Biophys. J. 2002, 83, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Burgess, S.; Wang, Z.; Vishnyakov, A.; Neimark, A.V. Adhesion, intake, and release of nanoparticles by lipid bilayers. J. Colloid Interface Sci. 2020, 561, 58–70. [Google Scholar] [CrossRef]
- Feng, Y.H.; Zhang, X.P.; Zhao, Z.Q.; Guo, X.D. Dissipative Particle Dynamics Aided Design of Drug Delivery Systems: A Review. Mol. Pharm. 2020, 17, 1778–1799. [Google Scholar] [CrossRef]
- Bahrami, A.H.; Lipowsky, R.; Weikl, T.R. Tubulation and Aggregation of Spherical Nanoparticles Adsorbed on Vesicles. Phys. Rev. Lett. 2012, 109, 188102. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, S.; Auth, T.; Gompper, G. Shape and Orientation Matter for the Cellular Uptake of Nonspherical Particles. Nano Lett. 2014, 14, 687–693. [Google Scholar] [CrossRef]
- Yang, K.; Ma, Y.Q. Computer simulation of the translocation of nanoparticles with different shapes across a lipid bilayer. Nat. Nanotechnol. 2010, 5, 579–583. [Google Scholar] [CrossRef]
- Zhao, Z.; Roy, D.; Steinkühler, J.; Robinson, T.; Lipowsky, R.; Dimova, R. Super-resolution imaging of highly curved membrane structures in giant vesicles encapsulating molecular condensates. Adv. Mater. 2022, 34, 2106633. [Google Scholar] [CrossRef]
- Drechsler, C.; Markones, M.; Choi, J.Y.; Frieling, N.; Fiedler, S.; Voelker, D.R.; Schubert, R.; Heerklotz, H. Preparation of Asymmetric Liposomes Using a Phosphatidylserine Decarboxylase. Biophys. J. 2018, 115, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Takaoka, R.; Kurosaki, H.; Nakao, H.; Ikeda, K.; Nakano, M. Formation of asymmetric vesicles via phospholipase D-mediated transphosphatidylation. Biochim. Biophys. Acta Biomembr. 2018, 1860, 245–249. [Google Scholar] [CrossRef]
- Salo-Ahen, O.M.H.; Alanko, I.; Bhadane, R.; Bonvin, A.M.J.J.; Honorato, R.V.; Hossain, S.; Juffer, A.H.; Kabedev, A.; Lahtela-Kakkonen, M.; Larsen, A.S.; et al. Molecular Dynamics Simulations in Drug Discovery and Pharmaceutical Development. Processes 2021, 9, 71. [Google Scholar] [CrossRef]
- Smrt, S.T.; Escobar, C.A.; Dey, S.; Cross, T.A.; Zhou, H.X. An Arg/Ala-rich helix in the N-terminal region of M. tuberculosis FtsQ is a potential membrane anchor of the Z-ring. Commun. Biol. 2023, 6, 311. [Google Scholar] [CrossRef]
- Cantor, R.S. Lateral pressures in cell membranes: A mechanism for modulation of protein function. J. Phys. Chem. B 1997, 101, 1723–1725. [Google Scholar] [CrossRef]
- Szleifer, I.; Kramer, D.; Ben-Shaul, A.; Gelbart, W.M.; Safran, S.A. Molecular theory of curvature elasticity in surfactant films. J. Chem. Phys. 1990, 92, 6800–6817. [Google Scholar] [CrossRef]
- Gompper, G.; Klein, S. Ginzburg-Landau theory of aqueous surfactant solutions. J. Phys. II 1992, 2, 1725–1744. [Google Scholar] [CrossRef]
- Feller, S.; Zhang, Y.; Pastor, R. Computer simulation of liquid/liquid intrefaces. II. Surface tension–area dependence of a bilayer and monolayer. J. Chem. Phys. 1995, 103, 10267–10276. [Google Scholar] [CrossRef]
- Schofield, P.; Henderson, J.R. Statistical Mechanics of Inhomogeneous Fluids. Proc. R. Soc. Lond. Ser. A 1982, 379, 231–246. [Google Scholar]
- Rowlinson, J.; Widom, B. Molecular Theory Capillarity; Oxford University Press: Oxford, UK, 1989. [Google Scholar]
- Bakker, G. Kapillarität und Oberflächenspannung; Handbuch der Experimentalphysik; Akademische Verlagsgesellschaft: Leipzig, Germany, 1928; Volume 6. [Google Scholar]
- Kirkwood, J.G.; Buff, F.P. The Statistical Mechanical Theory of Surface Tension. J. Chem. Phys. 1949, 17, 338–343. [Google Scholar] [CrossRef]
- Irving, J.H.; Kirkwood, J.G. The Statistical Mechanical Theory of Transport Processes. J. Chem. Phys. 1950, 18, 817–829. [Google Scholar] [CrossRef]
- Harasima, A. Molecular Theory of Surface Tension. In Advances in Chemical Physics; Prigogine, I., Debye, P., Eds.; Wiley: Hoboken, NJ, USA, 1958; Volume 1, pp. 203–237. [Google Scholar]












































Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lipowsky, R.; Ghosh, R.; Satarifard, V.; Sreekumari, A.; Zamaletdinov, M.; Różycki, B.; Miettinen, M.; Grafmüller, A. Leaflet Tensions Control the Spatio-Temporal Remodeling of Lipid Bilayers and Nanovesicles. Biomolecules 2023, 13, 926. https://doi.org/10.3390/biom13060926
Lipowsky R, Ghosh R, Satarifard V, Sreekumari A, Zamaletdinov M, Różycki B, Miettinen M, Grafmüller A. Leaflet Tensions Control the Spatio-Temporal Remodeling of Lipid Bilayers and Nanovesicles. Biomolecules. 2023; 13(6):926. https://doi.org/10.3390/biom13060926
Chicago/Turabian StyleLipowsky, Reinhard, Rikhia Ghosh, Vahid Satarifard, Aparna Sreekumari, Miftakh Zamaletdinov, Bartosz Różycki, Markus Miettinen, and Andrea Grafmüller. 2023. "Leaflet Tensions Control the Spatio-Temporal Remodeling of Lipid Bilayers and Nanovesicles" Biomolecules 13, no. 6: 926. https://doi.org/10.3390/biom13060926
APA StyleLipowsky, R., Ghosh, R., Satarifard, V., Sreekumari, A., Zamaletdinov, M., Różycki, B., Miettinen, M., & Grafmüller, A. (2023). Leaflet Tensions Control the Spatio-Temporal Remodeling of Lipid Bilayers and Nanovesicles. Biomolecules, 13(6), 926. https://doi.org/10.3390/biom13060926