
The Effects of an Osteoarthritic Joint Environment on ACL Damage and Degeneration: A Yucatan Miniature Pig Model

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Surgical Technique
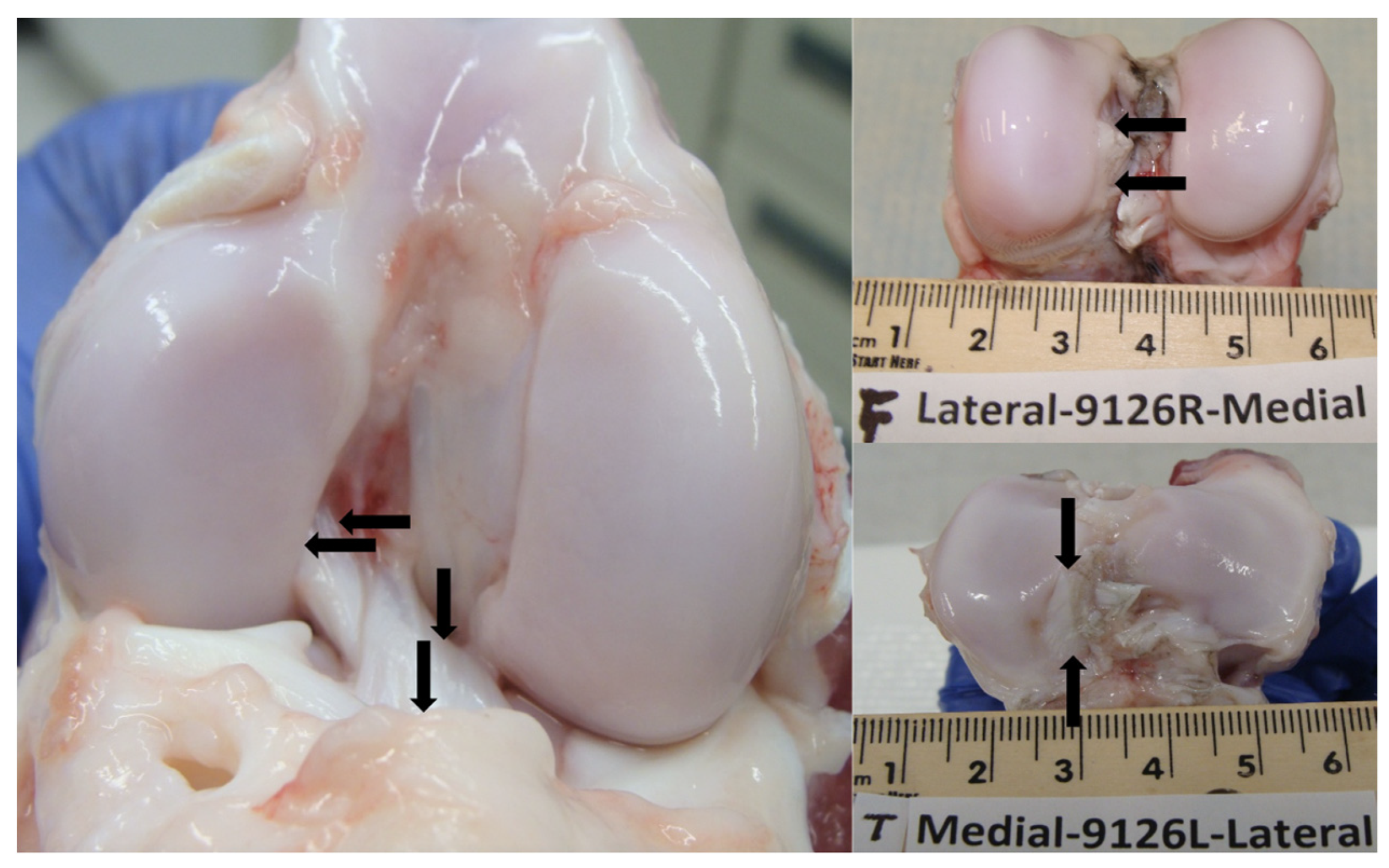
2.3. Macroscopic Cartilage Assessment
2.4. Cartilage Histological Preparation
2.5. Microscopic Cartilage Assessment
2.6. ACL Histological Preparation
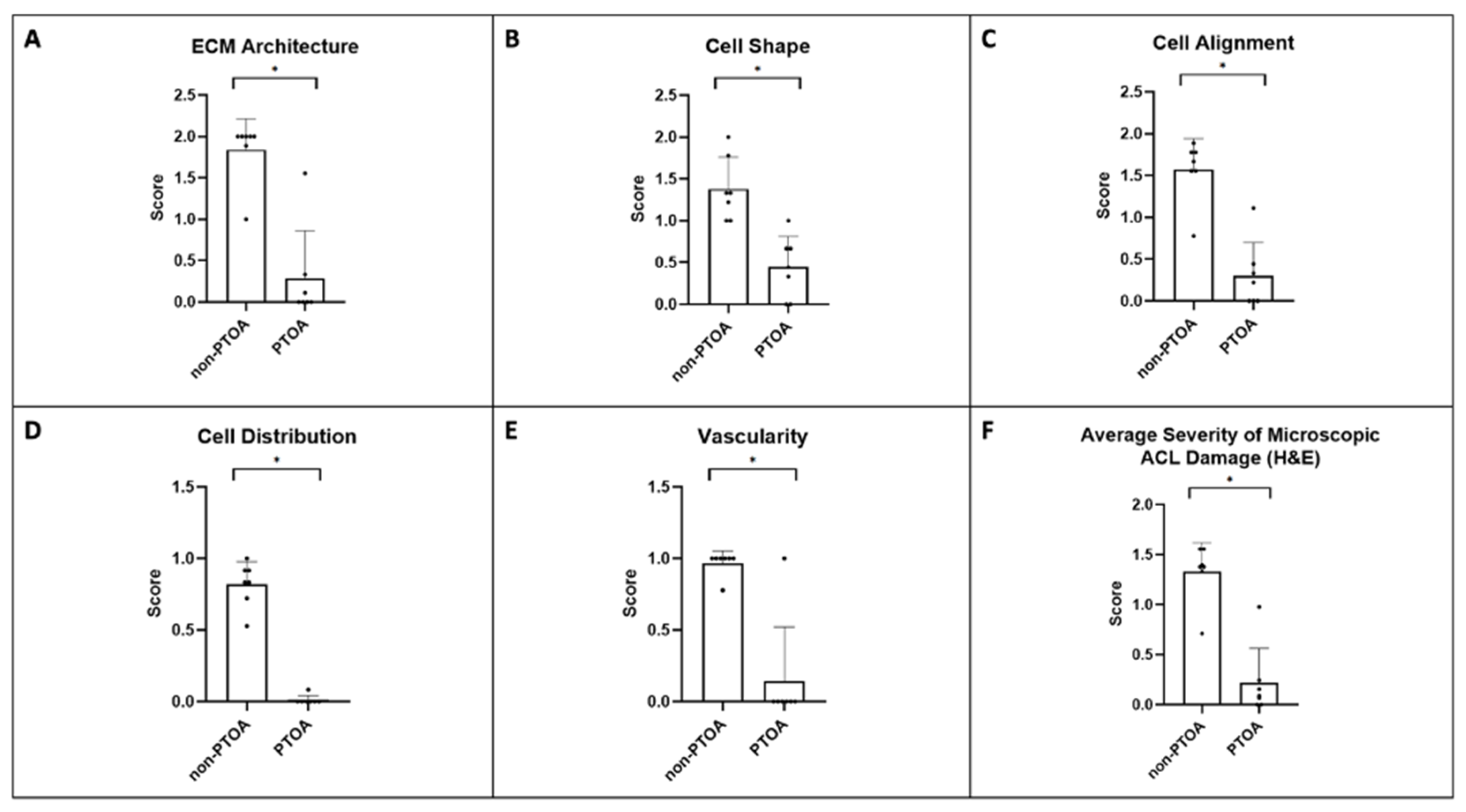
2.7. Microscopic ACL Assessment
2.8. Quantitative Real-Time PCR for Inflammatory Mediators
2.9. Immunohistochemistry
2.10. Statistical Methods
3. Results
3.1. Macroscopic Cartilage Assessment
3.2. Microscopic Cartilage Assessment
3.3. Microscopic ACL Assessment
3.4. Inflammatory Mediators
3.5. Immunohistochemistry
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheung, E.C.; DiLallo, M.; Feeley, B.T.; Lansdown, D.A. Osteoarthritis and ACL Reconstruction—Myths and Risks. Curr. Rev. Musculoskelet. Med. 2020, 13, 115–122. [Google Scholar] [CrossRef]
- Carbone, A.; Rodeo, S. Review of Current Understanding of Post-Traumatic Osteoarthritis Resulting from Sports Injuries. J. Orthop. Res. 2017, 35, 397–405. [Google Scholar] [CrossRef]
- Curado, J.; Hulet, C.; Hardy, P.; Jenny, J.-Y.; Rousseau, R.; Lucet, A.; Steltzlen, C.; Morin, V.; Grimaud, O.; Bouguennec, N.; et al. Very Long-Term Osteoarthritis Rate after Anterior Cruciate Ligament Reconstruction: 182 Cases with 22-Year’ Follow-up. Orthop. Traumatol. Surg. Res. 2020, 106, 459–463. [Google Scholar] [CrossRef]
- Alonso, B.; Bravo, B.; Mediavilla, L.; Gortazar, A.R.; Forriol, F.; Vaquero, J.; Guisasola, M.C. Osteoarthritis-Related Biomarkers Profile in Chronic Anterior Cruciate Ligament Injured Knee. Knee 2020, 27, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Huebner, K.D.; Shrive, N.G.; Frank, C.B. New Surgical Model of Post-Traumatic Osteoarthritis: Isolated Intra-Articular Bone Injury in the Rabbit. J. Orthop. Res. 2013, 31, 914–920. [Google Scholar] [CrossRef]
- Sun, C.; Chang, K.; Fleming, B.C.; Owens, B.D.; Beveridge, J.E.; Gage, A.; Talley-Bruns, R.C.; McAllister, S.; Costa, M.Q.; Pinette, M.P.; et al. A Novel Large Animal Model of Posttraumatic Osteoarthritis Induced by Inflammation with Mechanical Stability. Am. J. Transl. Res. 2023, 15, 4573. [Google Scholar] [PubMed]
- Otani, S.; Kanamoto, T.; Oyama, S.; Yamakawa, S.; Shi, W.; Miyazaki, R.; Aihara, M.; Oka, S.; Kuroda, S.; Nakai, T.; et al. Meniscus Surface Texture Is Associated with Degenerative Changes in Biological and Biomechanical Properties. Sci. Rep. 2022, 12, 11977. [Google Scholar] [CrossRef] [PubMed]
- Narama, I.; Masuoka-Nishiyama, M.; Matsuura, T.; Ozaki, K.; Nagatani, M.; Morishima, T. Morphogenesis of Degenerative Changes Predisposing Dogs to Rupture of the Cranial Cruciate Ligament. J. Vet. Med. Sci. 1996, 58, 1091–1097. [Google Scholar] [CrossRef]
- Hayashi, K.; Frank, J.D.; Hao, Z.; Schamberger, G.M.; Markel, M.D.; Manley, P.A.; Muir, P. Evaluation of Ligament Fibroblast Viability in Ruptured Cranial Cruciate Ligament of Dogs. Am. J. Vet. Res. 2003, 64, 1010–1016. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE Guidelines 2.0: Updated Guidelines for Reporting Animal Research. BMJ Open Sci. 2020, 4, e100115. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Wei, L.; Duan, Z.; Zhang, Z.; Chen, T.; Lu, J.; Zhao, R.; Cao, X.; Li, P.; Lv, Z.; et al. Contribution of IL-1β, 6 and TNF-α to the Form of Post-Traumatic Osteoarthritis Induced by “Idealized” Anterior Cruciate Ligament Reconstruction in a Porcine Model. Int. Immunopharmacol. 2018, 65, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Kiapour, A.M.; Fleming, B.C.; Murray, M.M. Structural and Anatomic Restoration of the Anterior Cruciate Ligament Is Associated with Less Cartilage Damage 1 Year after Surgery: Healing Ligament Properties Affect Cartilage Damage. Orthop. J. Sports Med. 2017, 5, 2325967117723886. [Google Scholar] [CrossRef]
- Little, C.B.; Smith, M.M.; Cake, M.A.; Read, R.A.; Murphy, M.J.; Barry, F.P. The OARSI Histopathology Initiative—Recommendations for Histological Assessments of Osteoarthritis in Sheep and Goats. Osteoarthr. Cartil. 2010, 18 (Suppl. S3), S80–S92. [Google Scholar] [CrossRef] [PubMed]
- Kharaz, Y.A.; Canty-Laird, E.G.; Tew, S.R.; Comerford, E.J. Variations in Internal Structure, Composition and Protein Distribution between Intra- and Extra-Articular Knee Ligaments and Tendons. J. Anat. 2018, 232, 943–955. [Google Scholar] [CrossRef] [PubMed]
- Kuchipudi, S.V.; Tellabati, M.; Nelli, R.K.; White, G.A.; Perez, B.B.; Sebastian, S.; Slomka, M.J.; Brookes, S.M.; Brown, I.H.; Dunham, S.P.; et al. 18S RRNA Is a Reliable Normalisation Gene for Real Time PCR Based on Influenza Virus Infected Cells. Virol. J. 2012, 9, 230. [Google Scholar] [CrossRef] [PubMed]
- Lohmander, L.S.; Englund, P.M.; Dahl, L.L.; Roos, E.M. The Long-Term Consequence of Anterior Cruciate Ligament and Meniscus Injuries: Osteoarthritis. Am. J. Sports Med. 2007, 35, 1756–1769. [Google Scholar] [CrossRef]
- Fleming, B.C.; Hulstyn, M.J.; Oksendahl, H.L.; Fadale, P.D. Ligament Injury, Reconstruction and Osteoarthritis. Curr. Opin. Orthop. 2005, 16, 354–362. [Google Scholar] [CrossRef]
- Kulkarni, P.; Koppikar, S.; Deshpande, S.; Wagh, N.; Harsulkar, A. Meniscal Tear as Potential Steering Factor for Inflammation May Aggravate Arthritis: Two Case Reports. J. Med. Case Rep. 2014, 8, 137. [Google Scholar] [CrossRef]
- McKinley, T.O.; Borrelli, J.; D’Lima, D.D.; Furman, B.D.; Giannoudis, P.V. Basic Science of Intraarticular Fractures and Posttraumatic Osteoarthritis. J. Orthop. Trauma 2010, 24, 567–570. [Google Scholar] [CrossRef]
- Hunt, E.R.; Jacobs, C.A.; Conley, C.E.-W.; Ireland, M.L.; Johnson, D.L.; Lattermann, C. Anterior Cruciate Ligament Reconstruction Reinitiates an Inflammatory and Chondrodegenerative Process in the Knee Joint. J. Orthop. Res. 2021, 39, 1281–1288. [Google Scholar] [CrossRef]
- Amano, K.; Huebner, J.L.; Stabler, T.V.; Tanaka, M.; McCulloch, C.E.; Lobach, I.; Lane, N.E.; Kraus, V.B.; Ma, C.B.; Li, X. Synovial Fluid Profile at the Time of Anterior Cruciate Ligament Reconstruction and Its Association with Cartilage Matrix Composition 3 Years after Surgery. Am. J. Sports Med. 2018, 46, 890–899. [Google Scholar] [CrossRef]
- Sieker, J.T.; Proffen, B.L.; Waller, K.A.; Chin, K.; Karamchedu, N.P.; Akelman, M.R.; Perrone, G.S.; Kiapour, A.M.; Konrad, J.; Murray, M.M.; et al. Transcriptional Profiling of Articular Cartilage in a Porcine Model of Early Post-Traumatic Osteoarthritis. J. Orthop. Res. 2018, 36, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Ojanen, S.P.; Finnilä, M.A.J.; Mäkelä, J.T.A.; Saarela, K.; Happonen, E.; Herzog, W.; Saarakkala, S.; Korhonen, R.K. Anterior Cruciate Ligament Transection of Rabbits Alters Composition, Structure and Biomechanics of Articular Cartilage and Chondrocyte Deformation 2 weeks Post-Surgery in a Site-Specific Manner. J. Biomech. 2020, 98, 109450. [Google Scholar] [CrossRef] [PubMed]
- Nagai-Tanima, M.; Ishikawa, A.; Kawai, H.; Ishida, K.; Kuroki, H.; Takakuwa, T.; Aoyama, T. The Opposing Effectongastrocnemius and Soleus Induced by Anterior Cruciate Ligament Transection: Gait Analysis and Muscle Assessment. Osteoarthr. Cartil. 2023, 31, 678–679. [Google Scholar] [CrossRef]
- Kim-Wang, S.Y.; Holt, A.G.; McGowan, A.M.; Danyluk, S.T.; Goode, A.P.; Lau, B.C.; Toth, A.P.; Wittstein, J.R.; DeFrate, L.E.; Yi, J.S.; et al. Immune Cell Profiles in Synovial Fluid after Anterior Cruciate Ligament and Meniscus Injuries. Arthritis Res. Ther. 2021, 23, 280. [Google Scholar] [CrossRef]
- Hasegawa, A.; Nakahara, H.; Kinoshita, M.; Asahara, H.; Koziol, J.; Lotz, M.K. Cellular and Extracellular Matrix Changes in Anterior Cruciate Ligaments during Human Knee Aging and Osteoarthritis. Arthritis Res. Ther. 2013, 15, R29. [Google Scholar] [CrossRef]
- Mendias, C.L.; Lynch, E.B.; Davis, M.E.; Sibilsky Enselman, E.R.; Harning, J.A.; Dewolf, P.D.; Makki, T.A.; Bedi, A. Changes in Circulating Biomarkers of Muscle Atrophy, Inflammation, and Cartilage Turnover in Patients Undergoing Anterior Cruciate Ligament Reconstruction and Rehabilitation. Am. J. Sports Med. 2013, 41, 1819–1826. [Google Scholar] [CrossRef]
- Beekhuizen, M.; Gierman, L.M.; van Spil, W.E.; Van Osch, G.J.V.M.; Huizinga, T.W.J.; Saris, D.B.F.; Creemers, L.B.; Zuurmond, A.-M. An Explorative Study Comparing Levels of Soluble Mediators in Control and Osteoarthritic Synovial Fluid. Osteoarthr. Cartil. 2013, 21, 918–922. [Google Scholar] [CrossRef]
- Mehana, E.-S.E.; Khafaga, A.F.; El-Blehi, S.S. The Role of Matrix Metalloproteinases in Osteoarthritis Pathogenesis: An Updated Review. Life Sci. 2019, 234, 116786. [Google Scholar] [CrossRef]
- Bell, R.D.; Shultz, S.J.; Wideman, L.; Henrich, V.C. Collagen Gene Variants Previously Associated with Anterior Cruciate Ligament Injury Risk Are Also Associated with Joint Laxity. Sports Health 2012, 4, 312–318. [Google Scholar] [CrossRef]
- Sanchez-Lopez, E.; Coras, R.; Torres, A.; Lane, N.E.; Guma, M. Synovial Inflammation in Osteoarthritis Progression. Nat. Rev. Rheumatol. 2022, 18, 258–275. [Google Scholar] [CrossRef] [PubMed]
- Haslauer, C.M.; Elsaid, K.A.; Fleming, B.C.; Proffen, B.L.; Johnson, V.M.; Murray, M.M. Loss of Extracellular Matrix from Articular Cartilage Is Mediated by the Synovium and Ligament after Anterior Cruciate Ligament Injury. Osteoarthr. Cartil. 2013, 21, 1950–1957. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Mucci, L.; Javaheri, B.; van’t Hof, R.; Bou-Gharios, G.; Pitsillides, A.A.; Comerford, E.; Poulet, B. Meniscal and Ligament Modifications in Spontaneous and Post-Traumatic Mouse Models of Osteoarthritis. Arthritis Res. Ther. 2020, 22, 171. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.A.; Humpherys, J.L.; Stannard, J.P.; Cook, J.L. Impact of Medial Meniscotibial Ligament Disruption Compared to Peripheral Medial Meniscal Tear on Knee Biomechanics. J. Knee Surg. 2021, 34, 784–792. [Google Scholar] [CrossRef]
- Spang, J.T.; Dang, A.B.C.; Mazzocca, A.; Rincon, L.; Obopilwe, E.; Beynnon, B.; Arciero, R.A. The Effect of Medial Meniscectomy and Meniscal Allograft Transplantation on Knee and Anterior Cruciate Ligament Biomechanics. Arthrosc. J. Arthrosc. Relat. Surg. 2010, 26, 192–201. [Google Scholar] [CrossRef]
- Griffin, T.M.; Guilak, F. The Role of Mechanical Loading in the Onset and Progression of Osteoarthritis. Exerc. Sports Sci. Rev. 2005, 33, 195. [Google Scholar] [CrossRef]
- Goldring, M.B.; Tsuchimochi, K.; Ijiri, K. The Control of Chondrogenesis. J. Cell Biochem. 2006, 97, 33–44. [Google Scholar] [CrossRef]
- Hasegawa, A.; Otsuki, S.; Pauli, C.; Miyaki, S.; Patil, S.; Steklov, N.; Kinoshita, M.; Koziol, J.; D’Lima, D.D.; Lotz, M.K. Anterior Cruciate Ligament Changes in Human Joint in Aging and Osteoarthritis. Arthritis Rheum. 2012, 64, 696–704. [Google Scholar] [CrossRef]
- Zhao, R.; Dong, Z.; Wei, X.; Gu, X.; Han, P.; Wu, H.; Yan, Y.; Huang, L.; Li, H.; Zhang, C.; et al. Inflammatory Factors Are Crucial for the Pathogenesis of Post-Traumatic Osteoarthritis Confirmed by a Novel Porcine Model: “Idealized” Anterior Cruciate Ligament Reconstruction and Gait Analysis. Int. Immunopharmacol. 2021, 99, 107905. [Google Scholar] [CrossRef]
- Khella, C.M.; Horvath, J.M.; Asgarian, R.; Rolauffs, B.; Hart, M.L. Anti-Inflammatory Therapeutic Approaches to Prevent or Delay Post-Traumatic Osteoarthritis (PTOA) of the Knee Joint with a Focus on Sustained Delivery Approaches. Int. J. Mol. Sci. 2021, 22, 8005. [Google Scholar] [CrossRef]
- Heard, B.J.; Barton, K.I.; Abubacker, S.; Chung, M.; Martin, C.R.; Schmidt, T.A.; Shrive, N.G.; Hart, D.A. Synovial and Cartilage Responsiveness to Peri-Operative Hyaluronic Acid ± Dexamethasone Administration Following a Limited Injury to the Rabbit Stifle Joint. J. Orthop. Res. 2022, 40, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Sarabia, F.; Coronel, P.; Collantes, E.; Navarro, F.J.; de la Serna, A.R.; Naranjo, A.; Gimeno, M.; Herrero-Beaumont, G. AMELIA study group A 40-Month Multicentre, Randomised Placebo-Controlled Study to Assess the Efficacy and Carry-over Effect of Repeated Intra-Articular Injections of Hyaluronic Acid in Knee Osteoarthritis: The AMELIA Project. Ann. Rheum. Dis. 2011, 70, 1957–1962. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wei, X.; Zhou, J.; Zhang, J.; Li, K.; Chen, Q.; Terek, R.; Fleming, B.C.; Goldring, M.B.; Ehrlich, M.G.; et al. Identification of Alpha 2 Macroglobulin (A2M) as a Master Inhibitor of Cartilage Degrading Factors That Attenuates Post-Traumatic Osteoarthritis Progression. Arthritis Rheumatol. 2014, 66, 1843–1853. [Google Scholar] [CrossRef]
- Sun, C.; Cao, C.; Zhao, T.; Guo, H.; Fleming, B.C.; Owens, B.; Beveridge, J.; McAllister, S.; Wei, L. A2M Inhibits Inflammatory Mediators of Chondrocytes by Blocking IL-1β/NF-ΚB Pathway. J. Orthop. Res. 2023, 41, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wei, X.; Browning, S.; Scuderi, G.; Hanna, L.S.; Wei, L. Targeted Designed Variants of Alpha-2-Macroglobulin (A2M) Attenuate Cartilage Degeneration in a Rat Model of Osteoarthritis Induced by Anterior Cruciate Ligament Transection. Arthritis Res. Ther. 2017, 19, 175. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequences | |
|---|---|---|
| IL-1β | Forward | 5’-CTG CAA ACT CCA GGA CAA AGA-3 |
| Reverse | 5’-GGG TGG CAT CAC AGA AAA-3’ | |
| NF-κB | Forward | 5’-ACT TGC CAG ACA CAG ATG AC-3’ |
| Reverse | 5’-GTC GGT GGG TCC ATT GAA A-3’ | |
| TNF-α | Forward | 5’-CCT ACT GCA CTT CGA GGT TATC-3’ |
| Reverse | 5’-ACG GGC TTA TCT GAG GTT TG-3’ | |
| 18S | Forward | 5’-GTAACCCGTTGAACCCCATT-3’ |
| Reverse | 5’-CCATCCAATCGGTAGTAGCG-3’ | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwartz, E.; Chang, K.; Sun, C.; Zhang, F.; Peng, G.; Owens, B.; Wei, L. The Effects of an Osteoarthritic Joint Environment on ACL Damage and Degeneration: A Yucatan Miniature Pig Model. Biomolecules 2023, 13, 1416. https://doi.org/10.3390/biom13091416
Schwartz E, Chang K, Sun C, Zhang F, Peng G, Owens B, Wei L. The Effects of an Osteoarthritic Joint Environment on ACL Damage and Degeneration: A Yucatan Miniature Pig Model. Biomolecules. 2023; 13(9):1416. https://doi.org/10.3390/biom13091416
Chicago/Turabian StyleSchwartz, Elias, Kenny Chang, Changqi Sun, Fei Zhang, Guoxuan Peng, Brett Owens, and Lei Wei. 2023. "The Effects of an Osteoarthritic Joint Environment on ACL Damage and Degeneration: A Yucatan Miniature Pig Model" Biomolecules 13, no. 9: 1416. https://doi.org/10.3390/biom13091416
APA StyleSchwartz, E., Chang, K., Sun, C., Zhang, F., Peng, G., Owens, B., & Wei, L. (2023). The Effects of an Osteoarthritic Joint Environment on ACL Damage and Degeneration: A Yucatan Miniature Pig Model. Biomolecules, 13(9), 1416. https://doi.org/10.3390/biom13091416