
Small Spleen Peptides (SSPs) Shape Dendritic Cell Differentiation through Modulation of Extracellular ATP Synthesis Profile

 , , and
, , and 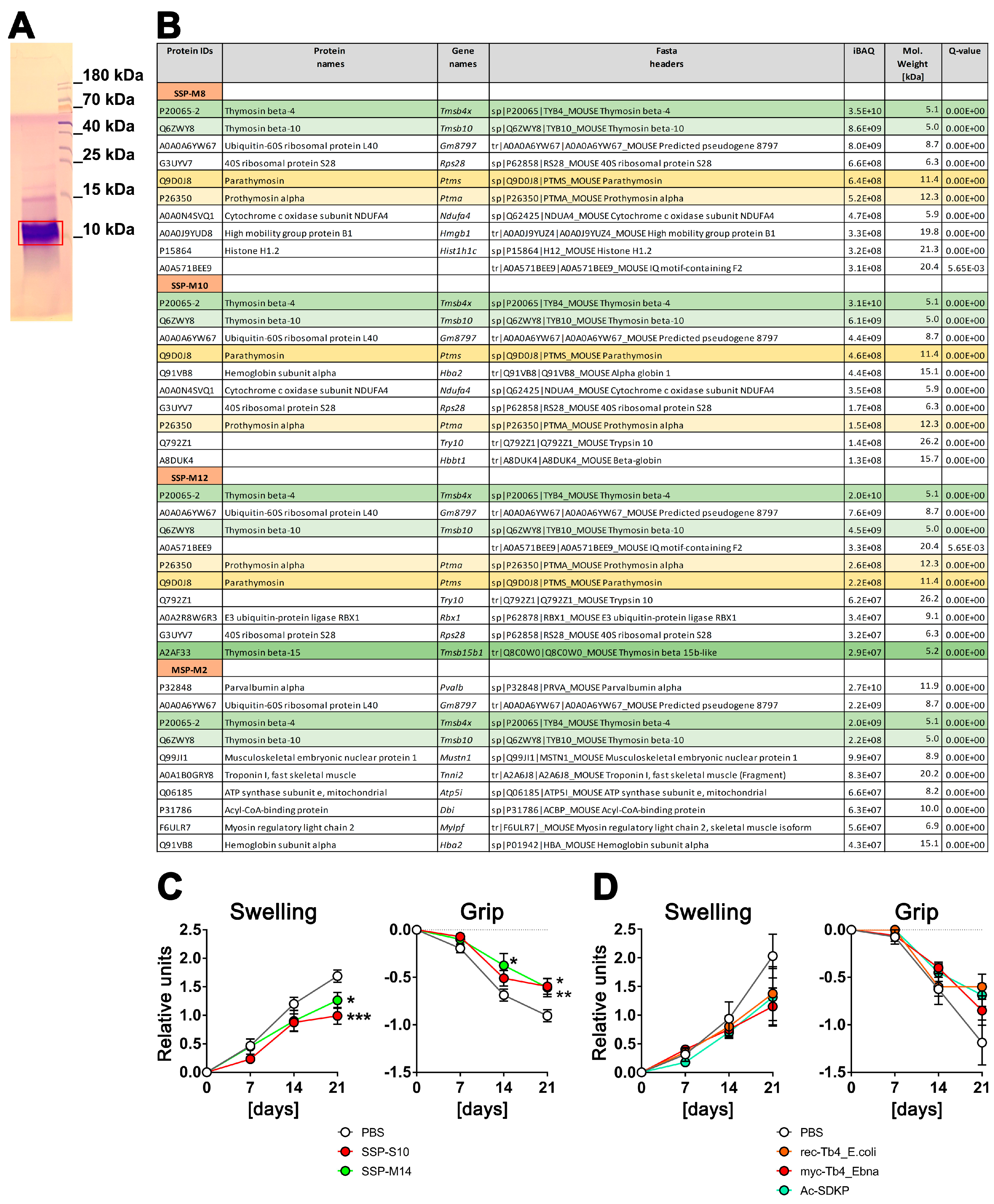{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Studies
2.2. Isolation and Differentiation of Primary Mouse Bone Marrow-Derived Macrophages and DCs
2.3. LC–MS/MS Analysis of SSP Probes
2.4. Cell Lines and Reagents
2.5. Mixed Lymphocyte Reaction and Flow Cytometry
2.6. RealTime-GloTM Extracellular ATP Assay
3. Results
3.1. LC–MS/MS Analysis of SSP Probes
3.2. Spleen-Derived Peptides with High Thymosin Content Provide Better Protection against Arthritis In Vivo Compared to Synthetically Derived Thymosins
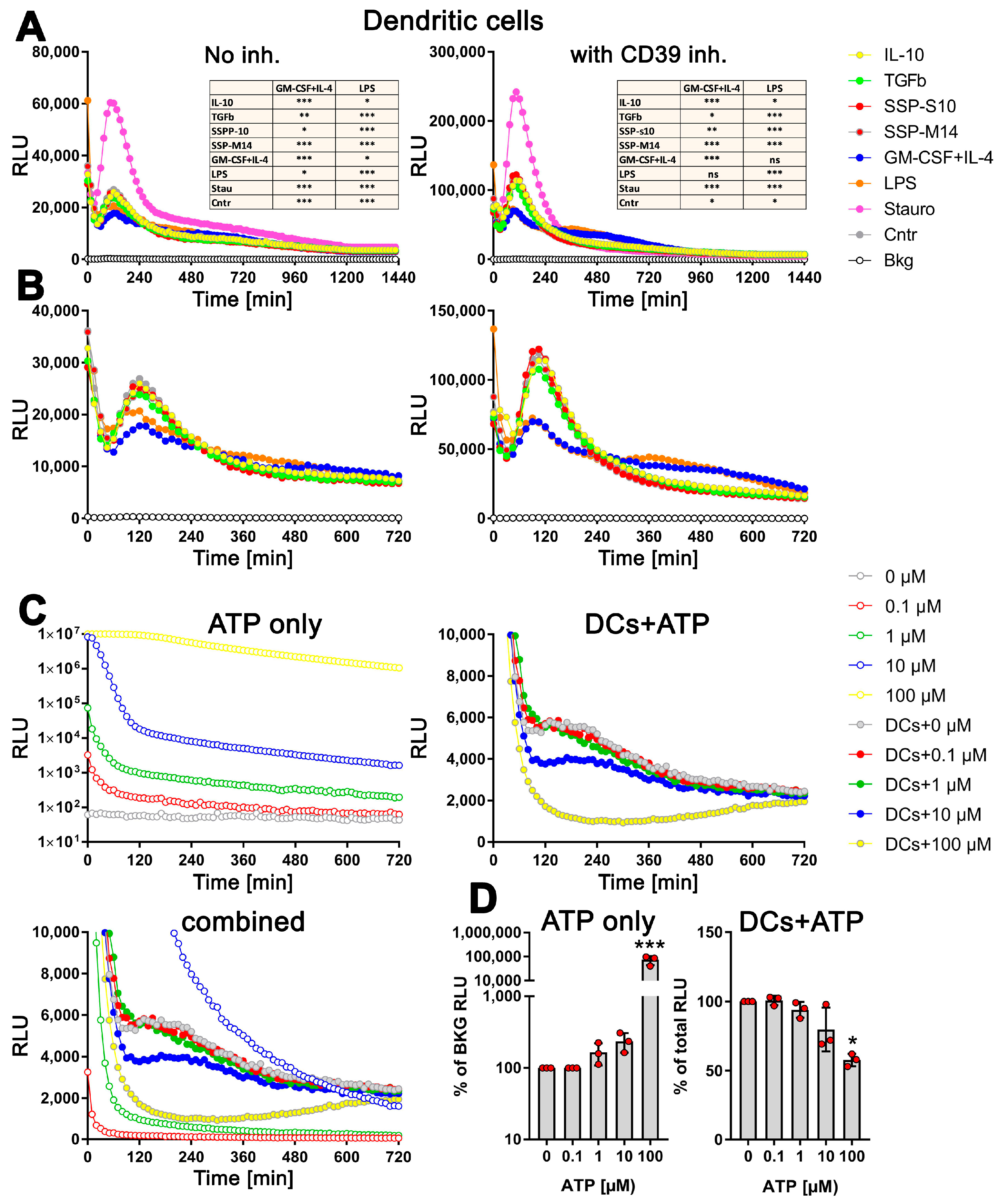
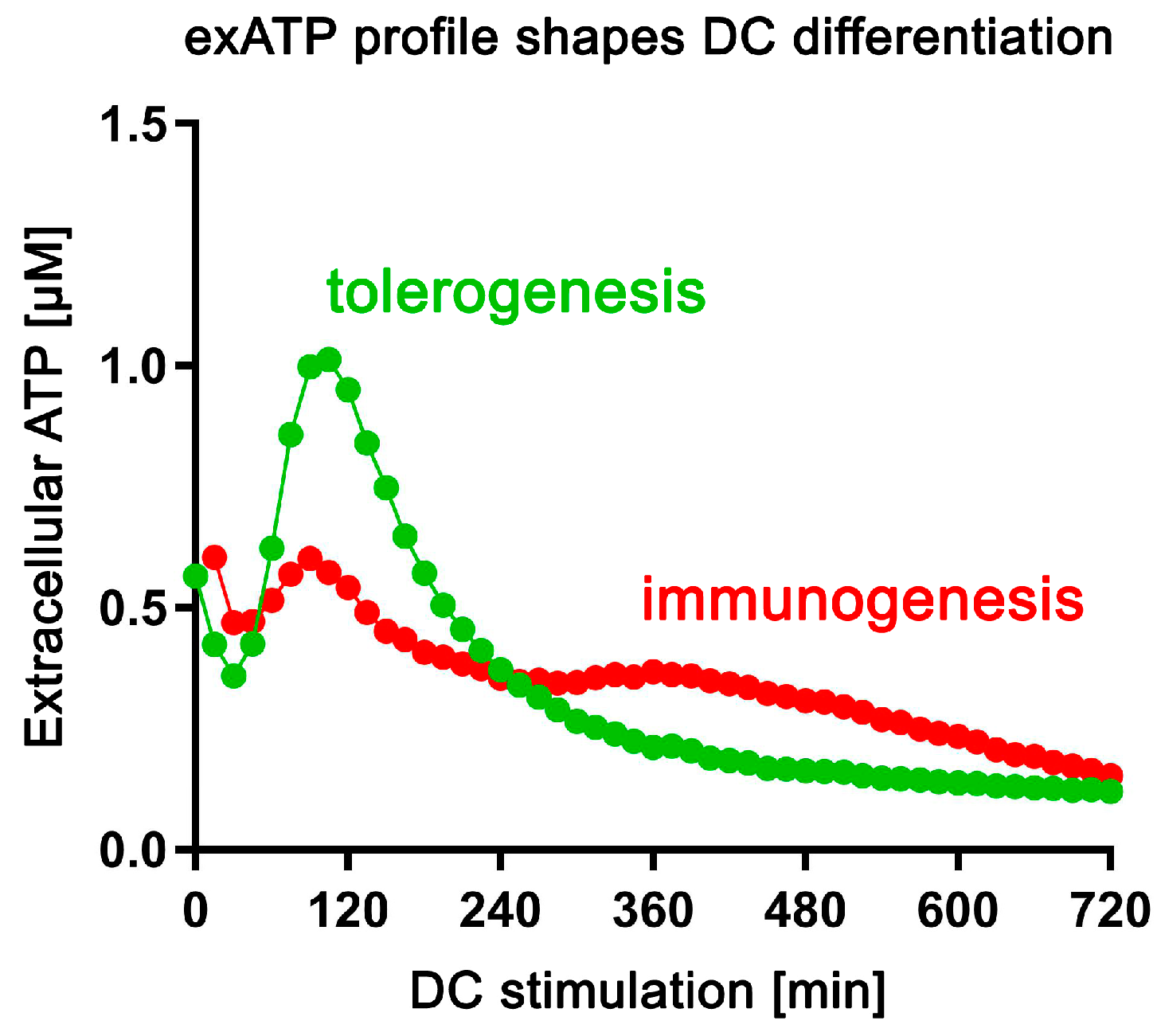
3.3. Stimulation of DCs with SSPs Results in a Distinctive Change in the Amount of exATP
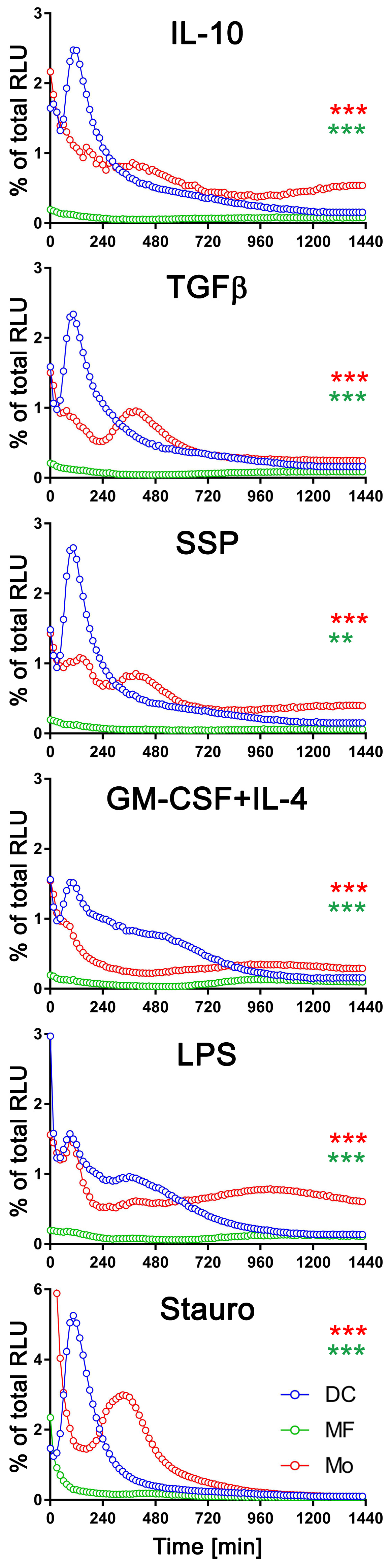
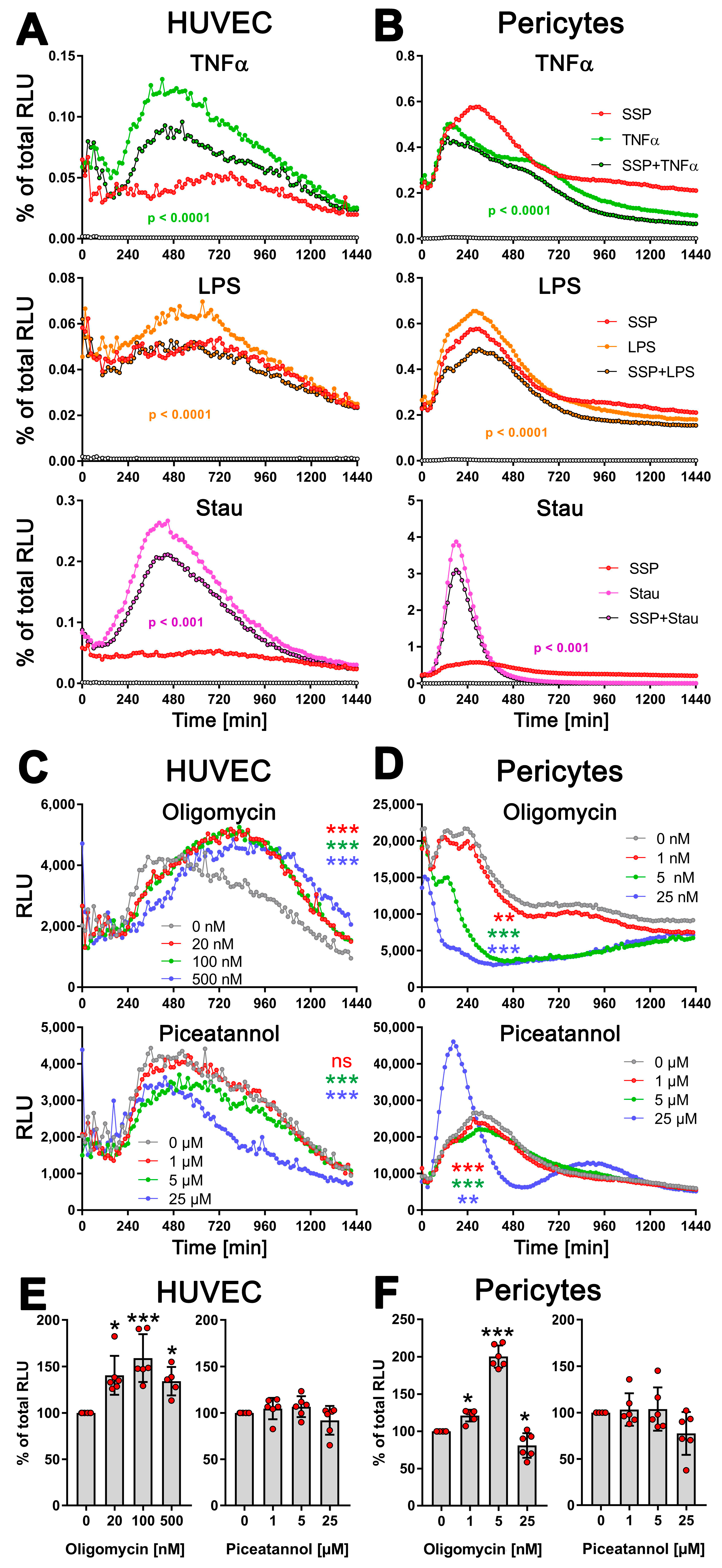
3.4. The exATP Profile Is Cell Type-Specific
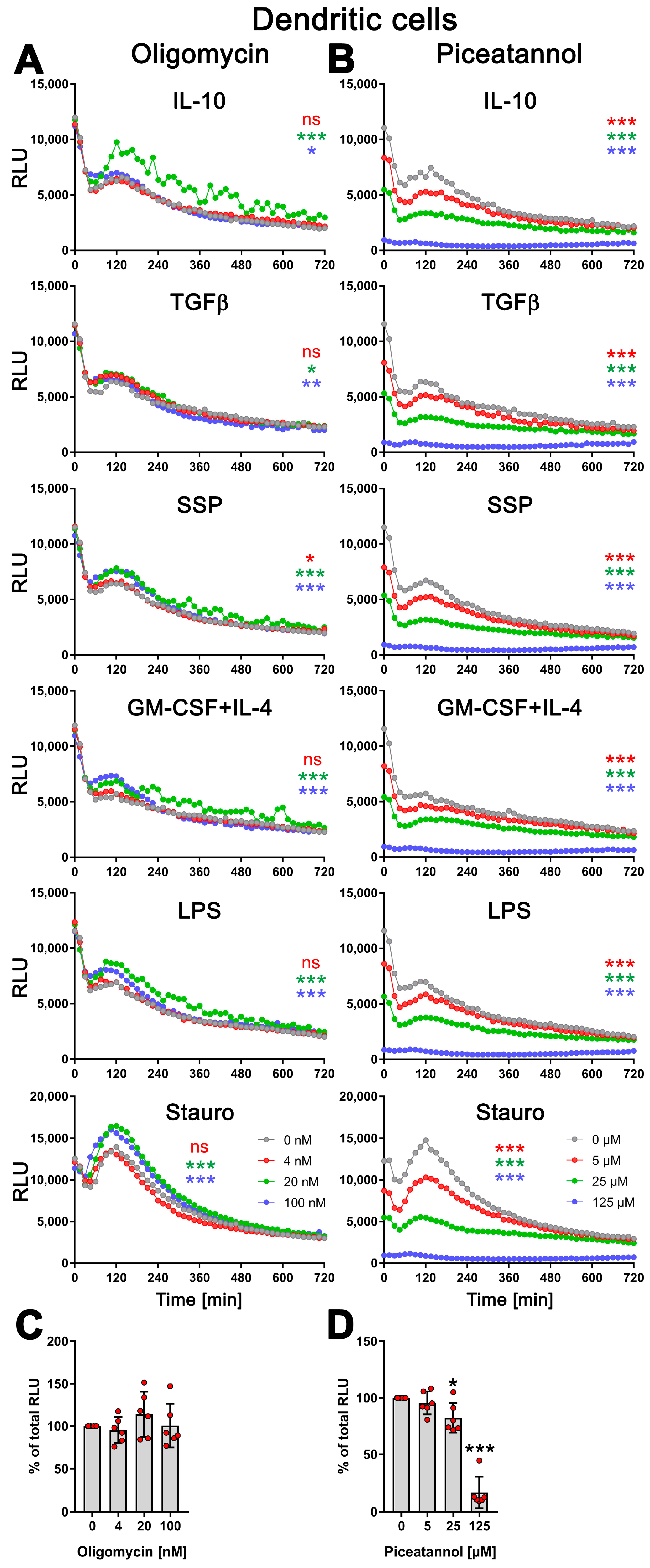
3.5. exATP Increase in DCs following Stimulation with Tolerogenic or Immunogenic Stimuli Is Attributed to the Synthesis of ATP
3.6. SSPs Suppress the Synthesis of exATP Mediated by Pro-Inflammatory Agents
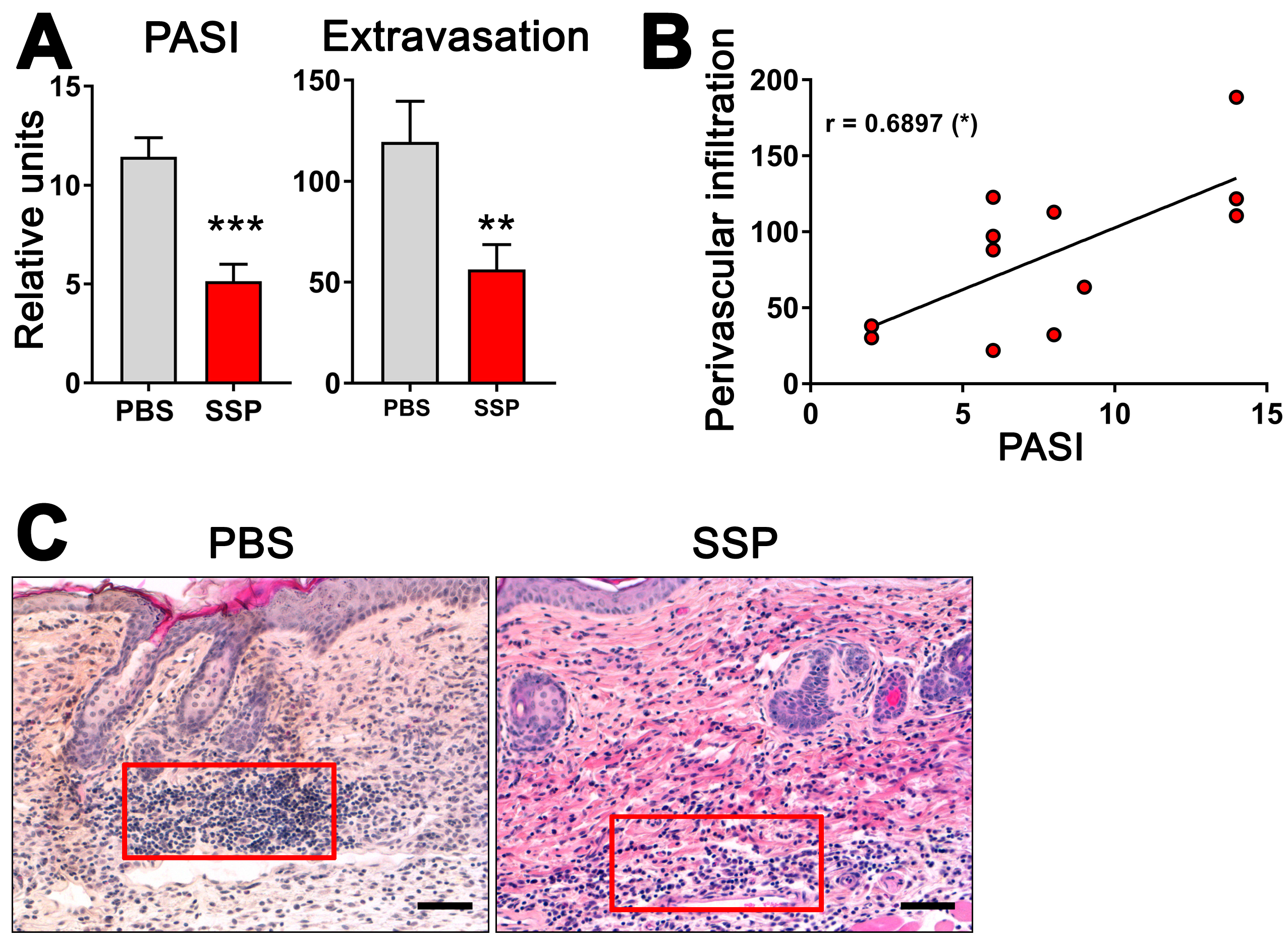
3.7. SSPs Reduce the Extravasation of Immune Cells into Psoriatic Skin
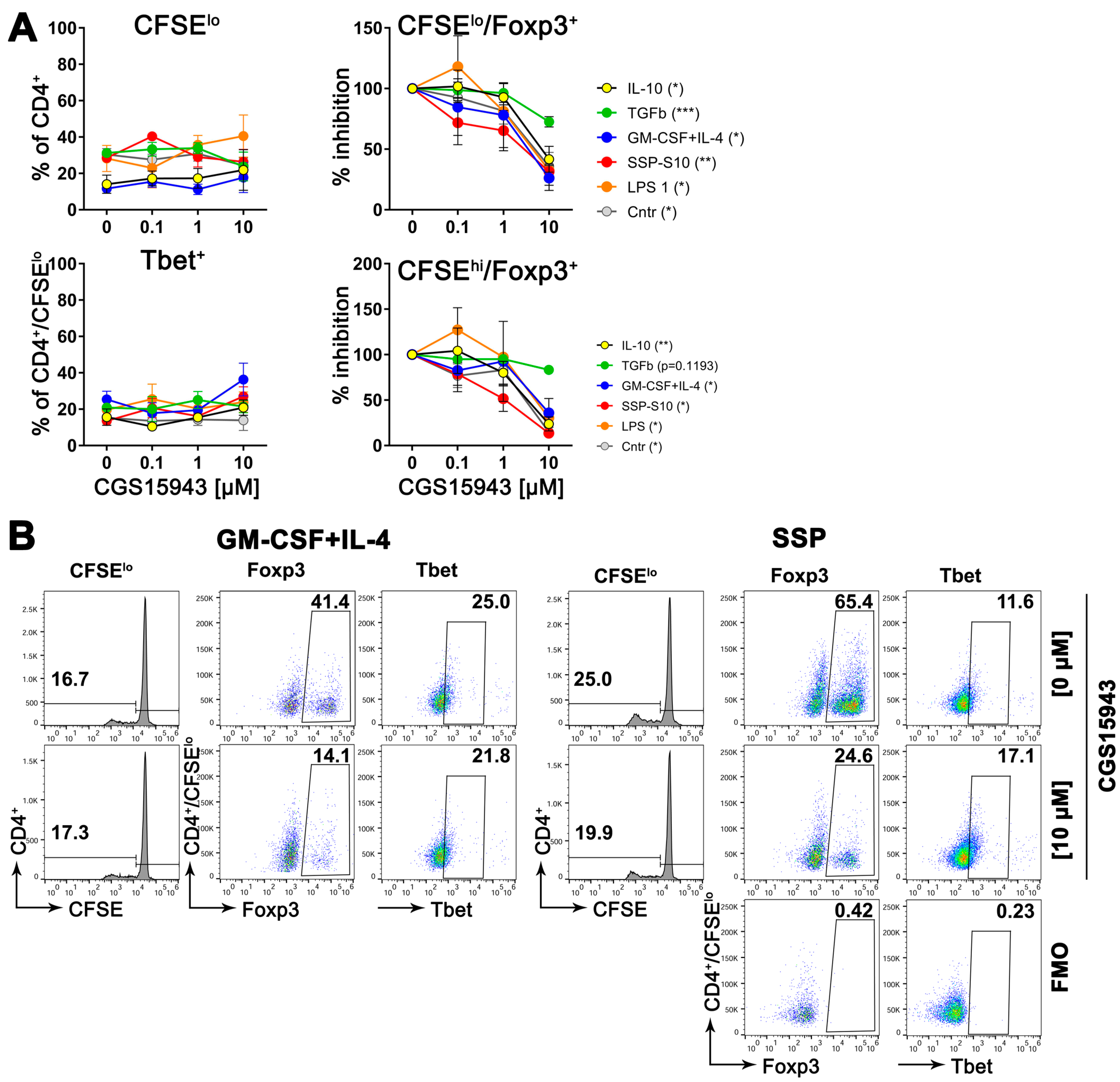
3.8. Inhibition of Adenosine Receptors Reduces the Formation of Treg Cells
4. Discussion
4.1. The Major Component of SSPs Is Tβ4
4.2. SSPs Induce De Novo Synthesis of exATP
4.3. Immunogenic and Tolerogenic Stimulation of DCs Elicit Divergent Effects on the Synthesis and Degradation of exATP
4.4. Multiple Functions of SSPs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wixler, V.; Zaytsev, I.Z.; Leite Dantas, R.; Schied, T.; Boergeling, Y.; Luhrmann, V.; Varga, G.; Masemann, D.; Ludwig, S. Small spleen peptides prevent development of psoriatic arthritis via restoration of peripheral tolerance. Mol. Ther. 2022, 30, 745–762. [Google Scholar] [CrossRef] [PubMed]
- Wixler, V.; Boergeling, Y.; Leite Dantas, R.; Varga, G.; Ludwig, S. Conversion of dendritic cells into tolerogenic or inflammatory cells depends on the activation threshold and kinetics of the mTOR signaling pathway. Cell Commun. Signal. 2024; in revision. [Google Scholar]
- Faas, M.M.; Saez, T.; de Vos, P. Extracellular ATP and adenosine: The Yin and Yang in immune responses? Mol. Asp. Med. 2017, 55, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.L.; Cai, Y.Q.; Zhu, M.C.; Xing, L.J.; Wang, X. The yin and yang functions of extracellular ATP and adenosine in tumor immunity. Cancer Cell Int. 2020, 20, 110. [Google Scholar] [CrossRef] [PubMed]
- Pasquini, S.; Contri, C.; Borea, P.A.; Vincenzi, F.; Varani, K. Adenosine and Inflammation: Here, There and Everywhere. Int. J. Mol. Sci. 2021, 22, 7685. [Google Scholar] [CrossRef] [PubMed]
- Silva-Vilches, C.; Ring, S.; Mahnke, K. ATP and Its Metabolite Adenosine as Regulators of Dendritic Cell Activity. Front. Immunol. 2018, 9, 2581. [Google Scholar] [CrossRef] [PubMed]
- Yegutkin, G.G. Nucleotide- and nucleoside-converting ectoenzymes: Important modulators of purinergic signalling cascade. Biochim. Biophys. Acta 2008, 1783, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Taurino, F.; Gnoni, A. Systematic review of plasma-membrane ecto-ATP synthase: A new player in health and disease. Exp. Mol. Pathol. 2018, 104, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Antonioli, L.; Pacher, P.; Vizi, E.S.; Hasko, G. CD39 and CD73 in immunity and inflammation. Trends Mol. Med. 2013, 19, 355–367. [Google Scholar] [CrossRef]
- Deaglio, S.; Dwyer, K.M.; Gao, W.; Friedman, D.; Usheva, A.; Erat, A.; Chen, J.F.; Enjyoji, K.; Linden, J.; Oukka, M.; et al. Adenosine generation catalyzed by CD39 and CD73 expressed on regulatory T cells mediates immune suppression. J. Exp. Med. 2007, 204, 1257–1265. [Google Scholar] [CrossRef]
- Silva-Vilches, C.; Ring, S.; Schrader, J.; Clausen, B.E.; Probst, H.C.; Melchior, F.; Schild, H.; Enk, A.; Mahnke, K. Production of Extracellular Adenosine by CD73(+) Dendritic Cells Is Crucial for Induction of Tolerance in Contact Hypersensitivity Reactions. J. Investig. Dermatol. 2019, 139, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Pantely, G.A.; Bristow, J.D. Adenosine. Renewed interest in an old drug. Circulation 1990, 82, 1854–1856. [Google Scholar] [CrossRef] [PubMed]
- Panther, E.; Corinti, S.; Idzko, M.; Herouy, Y.; Napp, M.; la Sala, A.; Girolomoni, G.; Norgauer, J. Adenosine affects expression of membrane molecules, cytokine and chemokine release, and the T-cell stimulatory capacity of human dendritic cells. Blood 2003, 101, 3985–3990. [Google Scholar] [CrossRef] [PubMed]
- Borea, P.A.; Gessi, S.; Merighi, S.; Varani, K. Adenosine as a Multi-Signalling Guardian Angel in Human Diseases: When, Where and How Does it Exert its Protective Effects? Trends Pharmacol. Sci. 2016, 37, 419–434. [Google Scholar] [CrossRef] [PubMed]
- Bahreyni, A.; Khazaei, M.; Rajabian, M.; Ryzhikov, M.; Avan, A.; Hassanian, S.M. Therapeutic potency of pharmacological adenosine receptor agonist/antagonist in angiogenesis, current status and perspectives. J. Pharm. Pharmacol. 2018, 70, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Ohta, A.; Sitkovsky, M. Extracellular adenosine-mediated modulation of regulatory T cells. Front. Immunol. 2014, 5, 304. [Google Scholar] [CrossRef] [PubMed]
- Retser, E.; Schied, T.; Skryabin, B.V.; Vogl, T.; Kanczler, J.M.; Hamann, N.; Niehoff, A.; Hermann, S.; Eisenblatter, M.; Wachsmuth, L.; et al. Doxycycline-induced expression of transgenic human tumor necrosis factor alpha in adult mice results in psoriasis-like arthritis. Arthritis Rheum. 2013, 65, 2290–2300. [Google Scholar] [CrossRef] [PubMed]
- Leite Dantas, R.; Brachvogel, B.; Schied, T.; Bergmeier, V.; Skryabin, B.; Vogl, T.; Ludwig, S.; Wixler, V. The LIM-Only Protein Four and a Half LIM Domain Protein 2 Attenuates Development of Psoriatic Arthritis by Blocking Adam17-Mediated Tumor Necrosis Factor Release. Am. J. Pathol. 2017, 187, 2388–2398. [Google Scholar] [CrossRef] [PubMed]
- Kall, L.; Storey, J.D.; MacCoss, M.J.; Noble, W.S. Posterior error probabilities and false discovery rates: Two sides of the same coin. J. Proteome Res. 2008, 7, 40–44. [Google Scholar] [CrossRef]
- Nordmann, A.; Wixler, L.; Ludwig, S.; Wixler, V. Monoclonal antibodies against the PB1-F2 protein of H1N1 influenza A virus. Hybridoma 2010, 29, 321–326. [Google Scholar] [CrossRef]
- Dominari, A.; Hathaway Iii, D.; Pandav, K.; Matos, W.; Biswas, S.; Reddy, G.; Thevuthasan, S.; Khan, M.A.; Mathew, A.; Makkar, S.S.; et al. Thymosin alpha 1: A comprehensive review of the literature. World J. Virol. 2020, 9, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Hara, T. Thymosins and muscle regeneration. Vitam. Horm. 2011, 87, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Kuzan, A. Thymosin beta as an Actin-binding Protein with a Variety of Functions. Adv. Clin. Exp. Med. 2016, 25, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Philp, D.; Kleinman, H.K. Animal studies with thymosin beta, a multifunctional tissue repair and regeneration peptide. Ann. N. Y. Acad. Sci. 2010, 1194, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Hannappel, E. Thymosin beta4 and its posttranslational modifications. Ann. N. Y. Acad. Sci. 2010, 1194, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Freeman, K.W.; Bowman, B.R.; Zetter, B.R. Regenerative protein thymosin beta-4 is a novel regulator of purinergic signaling. Faseb J. 2011, 25, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Leist, M.; Single, B.; Castoldi, A.F.; Kuhnle, S.; Nicotera, P. Intracellular adenosine triphosphate (ATP) concentration: A switch in the decision between apoptosis and necrosis. J. Exp. Med. 1997, 185, 1481–1486. [Google Scholar] [CrossRef] [PubMed]
- Neupane, P.; Bhuju, S.; Thapa, N.; Bhattarai, H.K. ATP Synthase: Structure, Function and Inhibition. Biomol. Concepts 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cauwels, A.; Rogge, E.; Vandendriessche, B.; Shiva, S.; Brouckaert, P. Extracellular ATP drives systemic inflammation, tissue damage and mortality. Cell Death Dis. 2014, 5, e1102. [Google Scholar] [CrossRef]
- Dosch, M.; Gerber, J.; Jebbawi, F.; Beldi, G. Mechanisms of ATP Release by Inflammatory Cells. Int. J. Mol. Sci. 2018, 19, 1222. [Google Scholar] [CrossRef]
- Xing, Y.; Ye, Y.; Zuo, H.; Li, Y. Progress on the Function and Application of Thymosin beta4. Front. Endocrinol. 2021, 12, 767785. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.L.; Hannappel, E.; Sosne, G.; Kleinman, H.K. Thymosin beta4: A multi-functional regenerative peptide. Basic properties and clinical applications. Expert Opin. Biol. Ther. 2012, 12, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Liao, T.D.; Romero, C.A.; Maheshwari, M.; Peterson, E.L.; Carretero, O.A. Thymosin beta4 Deficiency Exacerbates Renal and Cardiac Injury in Angiotensin-II-Induced Hypertension. Hypertension 2018, 71, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Yin, C. The anti-inflammatory peptide Ac-SDKP: Synthesis, role in ACE inhibition, and its therapeutic potential in hypertension and cardiovascular diseases. Pharmacol. Res. 2018, 134, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Yang, H.I. Thymosin beta4 in rheumatoid arthritis: Friend or foe. Biomed. Rep. 2017, 7, 205–208. [Google Scholar] [CrossRef]
- Gatto, C.; Grandi, M.; Solaini, G.; Baracca, A.; Giorgio, V. The F1Fo-ATPase inhibitor protein IF1 in pathophysiology. Front. Physiol. 2022, 13, 917203. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Aguilar, A.; Cuezva, J.M. A Review of the Inhibition of the Mitochondrial ATP Synthase by IF1 in vivo: Reprogramming Energy Metabolism and Inducing Mitohormesis. Front. Physiol. 2018, 9, 1322. [Google Scholar] [CrossRef] [PubMed]
- Gore, E.; Duparc, T.; Genoux, A.; Perret, B.; Najib, S.; Martinez, L.O. The Multifaceted ATPase Inhibitory Factor 1 (IF1) in Energy Metabolism Reprogramming and Mitochondrial Dysfunction: A New Player in Age-Associated Disorders? Antioxid. Redox Signal. 2022, 37, 370–393. [Google Scholar] [CrossRef]
- Ziganshin, A.U.; Ziganshina, L.E.; King, B.F.; Burnstock, G. Differential degradation of extracellular adenine nucleotides by folliculated oocytes of Xenopus laevis. Comp. Biochem. Physiol. A Physiol. 1996, 114, 335–340. [Google Scholar] [CrossRef]
- Greiner, J.V.; Glonek, T. Intracellular ATP Concentration and Implication for Cellular Evolution. Biology 2021, 10, 1166. [Google Scholar] [CrossRef]
- Bastid, J.; Regairaz, A.; Bonnefoy, N.; Dejou, C.; Giustiniani, J.; Laheurte, C.; Cochaud, S.; Laprevotte, E.; Funck-Brentano, E.; Hemon, P.; et al. Inhibition of CD39 enzymatic function at the surface of tumor cells alleviates their immunosuppressive activity. Cancer Immunol. Res. 2015, 3, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Bynoe, M.S.; Viret, C. Foxp3+CD4+ T cell-mediated immunosuppression involves extracellular nucleotide catabolism. Trends Immunol. 2008, 29, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Bo, C.; Kang, Y.; Li, H. What Else Can CD39 Tell Us? Front. Immunol. 2017, 8, 727. [Google Scholar] [CrossRef] [PubMed]
- Malinarich, F.; Duan, K.; Hamid, R.A.; Bijin, A.; Lin, W.X.; Poidinger, M.; Fairhurst, A.M.; Connolly, J.E. High mitochondrial respiration and glycolytic capacity represent a metabolic phenotype of human tolerogenic dendritic cells. J. Immunol. 2015, 194, 5174–5186. [Google Scholar] [CrossRef] [PubMed]
- Lohman, A.W.; Billaud, M.; Isakson, B.E. Mechanisms of ATP release and signalling in the blood vessel wall. Cardiovasc. Res. 2012, 95, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B. Adenosine, an endogenous distress signal, modulates tissue damage and repair. Cell Death Differ. 2007, 14, 1315–1323. [Google Scholar] [CrossRef] [PubMed]
- Munshaw, S.; Bruche, S.; Redpath, A.N.; Jones, A.; Patel, J.; Dube, K.N.; Lee, R.; Hester, S.S.; Davies, R.; Neal, G.; et al. Thymosin beta4 protects against aortic aneurysm via endocytic regulation of growth factor signaling. J. Clin. Investig. 2021, 131, e127884. [Google Scholar] [CrossRef] [PubMed]
- Rossdeutsch, A.; Smart, N.; Dube, K.N.; Turner, M.; Riley, P.R. Essential role for thymosin beta4 in regulating vascular smooth muscle cell development and vessel wall stability. Circ. Res. 2012, 111, e89–e102. [Google Scholar] [CrossRef]
- Dube, K.N.; Bollini, S.; Smart, N.; Riley, P.R. Thymosin beta4 protein therapy for cardiac repair. Curr. Pharm. Des. 2012, 18, 799–806. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wixler, V.; Leite Dantas, R.; Varga, G.; Boergeling, Y.; Ludwig, S. Small Spleen Peptides (SSPs) Shape Dendritic Cell Differentiation through Modulation of Extracellular ATP Synthesis Profile. Biomolecules 2024, 14, 469. https://doi.org/10.3390/biom14040469
Wixler V, Leite Dantas R, Varga G, Boergeling Y, Ludwig S. Small Spleen Peptides (SSPs) Shape Dendritic Cell Differentiation through Modulation of Extracellular ATP Synthesis Profile. Biomolecules. 2024; 14(4):469. https://doi.org/10.3390/biom14040469
Chicago/Turabian StyleWixler, Viktor, Rafael Leite Dantas, Georg Varga, Yvonne Boergeling, and Stephan Ludwig. 2024. "Small Spleen Peptides (SSPs) Shape Dendritic Cell Differentiation through Modulation of Extracellular ATP Synthesis Profile" Biomolecules 14, no. 4: 469. https://doi.org/10.3390/biom14040469
APA StyleWixler, V., Leite Dantas, R., Varga, G., Boergeling, Y., & Ludwig, S. (2024). Small Spleen Peptides (SSPs) Shape Dendritic Cell Differentiation through Modulation of Extracellular ATP Synthesis Profile. Biomolecules, 14(4), 469. https://doi.org/10.3390/biom14040469