
TRAF1 from a Structural Perspective



{kind=link}
{kind=link}
Abstract
:1. Introduction

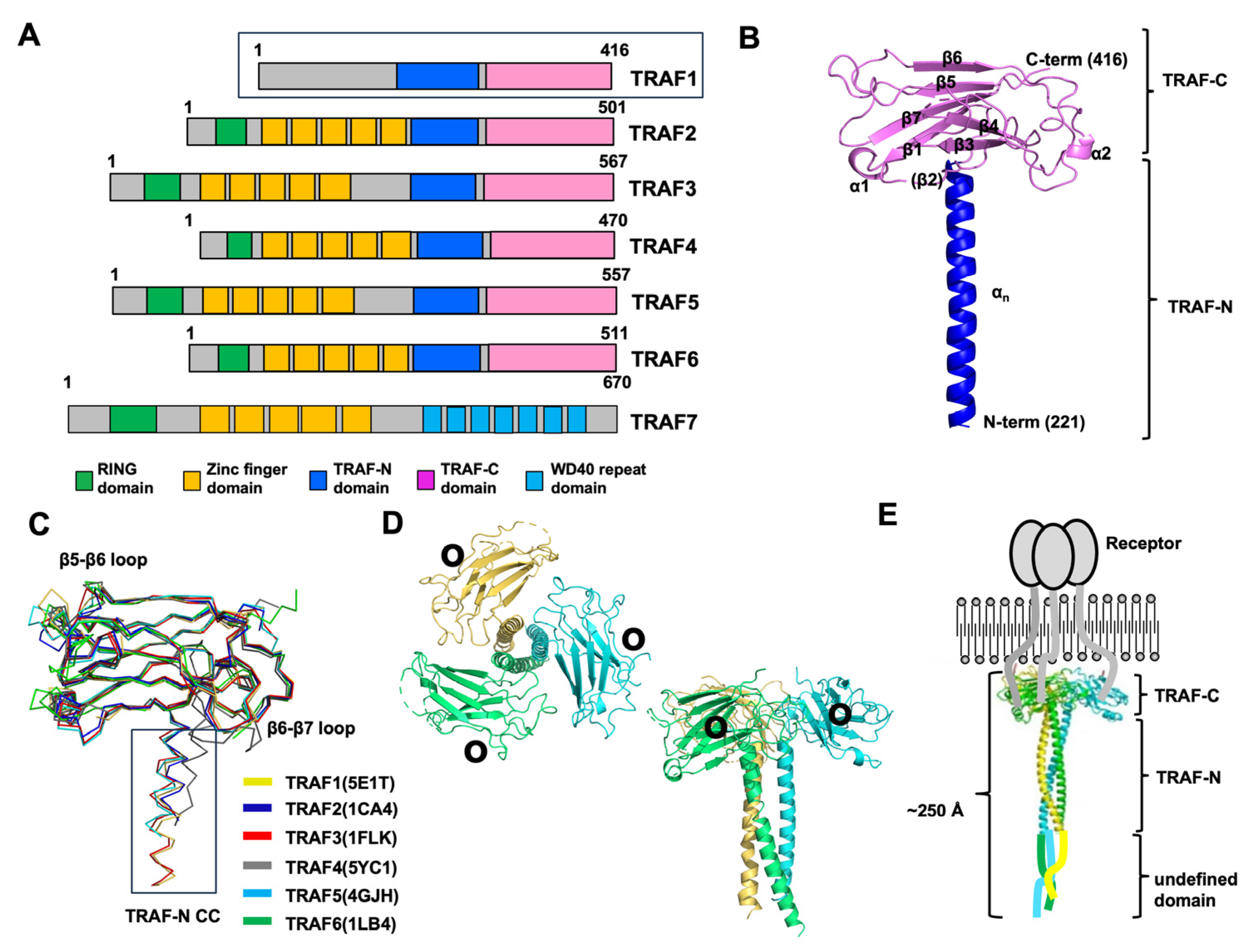
2. TRAF Domains and Structure
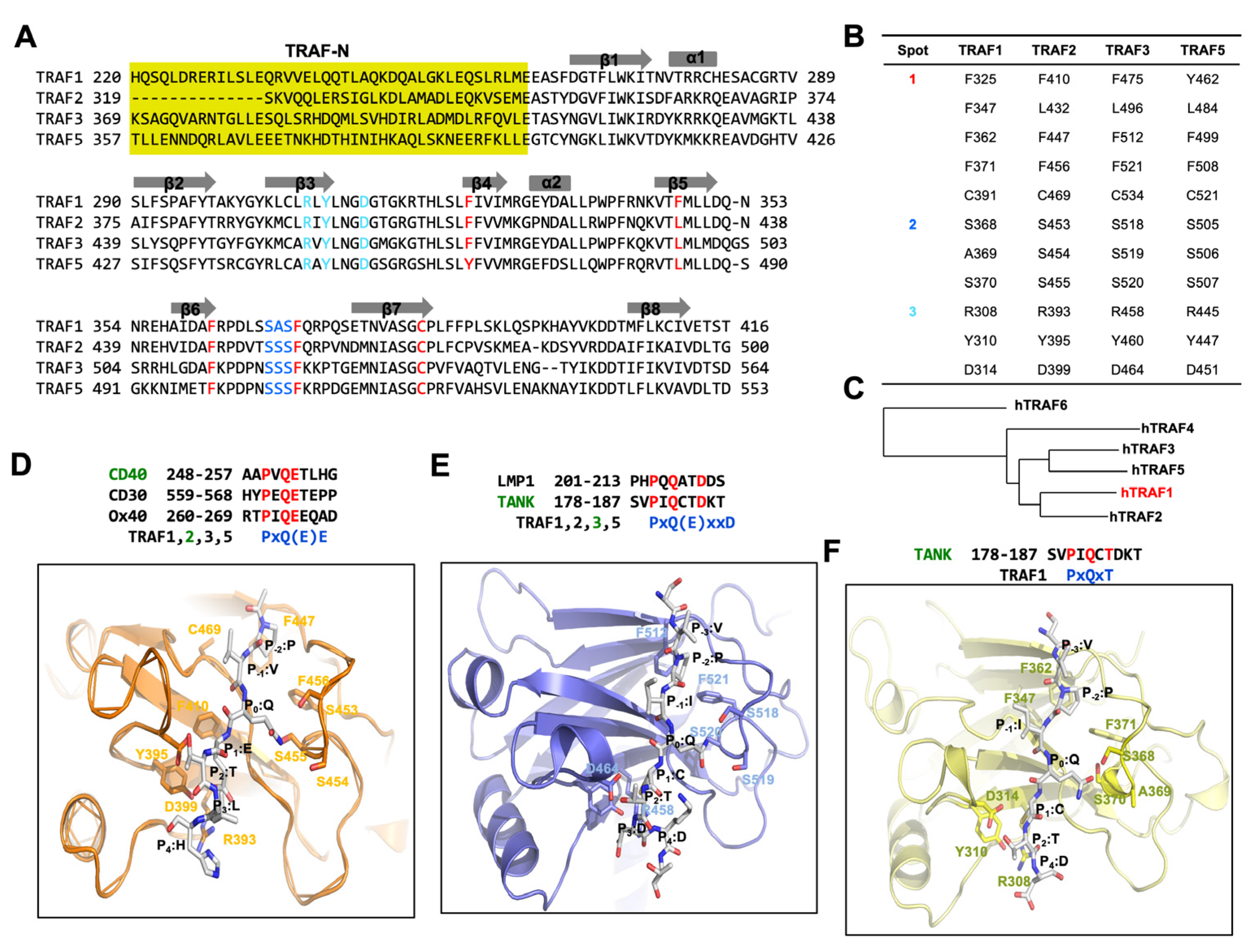
3. Understanding Receptor Interaction with TRAF Proteins
4. TRAF1 and Its Receptor Recognition
5. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arkee, T.; Bishop, G.A. TRAF family molecules in T cells: Multiple receptors and functions. J. Leukoc. Biol. 2020, 107, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Bishop, G.A. The multifaceted roles of TRAFs in the regulation of B-cell function. Nat. Rev. Immunol. 2004, 4, 775–786. [Google Scholar] [CrossRef]
- Wajant, H.; Henkler, F.; Scheurich, P. The TNF-receptor-associated factor family: Scaffold molecules for cytokine receptors, kinases and their regulators. Cell Signal 2001, 13, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Bradley, J.R.; Pober, J.S. Tumor necrosis factor receptor-associated factors (TRAFs). Oncogene 2001, 20, 6482–6491. [Google Scholar] [CrossRef] [PubMed]
- Deshaies, R.J.; Joazeiro, C.A. RING domain E3 ubiquitin ligases. Annu. Rev. Biochem. 2009, 78, 399–434. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.D.; Sun, S.C. Targeting signaling factors for degradation, an emerging mechanism for TRAF functions. Immunol. Rev. 2015, 266, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Rothe, M.; Wong, S.C.; Henzel, W.J.; Goeddel, D.V. A novel family of putative signal transducers associated with the cytoplasmic domain of the 75 kDa tumor necrosis factor receptor. Cell 1994, 78, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Zapata, J.M.; Pawlowski, K.; Haas, E.; Ware, C.F.; Godzik, A.; Reed, J.C. TEFs: A diverse family of proteins containing TRAF domains. J. Biol. Chem. 2001, 276, 24242–24252. [Google Scholar] [CrossRef] [PubMed]
- Arch, R.H.; Gedrich, R.W.; Thompson, C.B. Tumor necrosis factor receptor-associated factors (TRAFs)—A family of adapter proteins that regulates life and death. Genes Dev. 1998, 12, 2821–2830. [Google Scholar] [CrossRef]
- Hsu, H.; Huang, J.; Shu, H.B.; Baichwal, V.; Goeddel, D.V. TNF-dependent recruitment of the protein kinase RIP to the TNF receptor-1 signaling complex. Immunity 1996, 4, 387–396. [Google Scholar] [CrossRef]
- Song, H.Y.; Regnier, C.H.; Kirschning, C.J.; Goeddel, D.V.; Rothe, M. Tumor necrosis factor (TNF)-mediated kinase cascades: Bifurcation of nuclear factor-kappaB and c-jun N-terminal kinase (JNK/SAPK) pathways at TNF receptor-associated factor 2. Proc. Natl. Acad. Sci. USA 1997, 94, 9792–9796. [Google Scholar] [CrossRef] [PubMed]
- Hoeflich, K.P.; Yeh, W.C.; Yao, Z.; Mak, T.W.; Woodgett, J.R. Mediation of TNF receptor-associated factor effector functions by apoptosis signal-regulating kinase-1 (ASK1). Oncogene 1999, 18, 5814–5820. [Google Scholar] [CrossRef] [PubMed]
- Hacker, H.; Tseng, P.H.; Karin, M. Expanding TRAF function: TRAF3 as a tri-faced immune regulator. Nat. Rev. Immunol. 2011, 11, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Sundar, R.; Thakur, N.; Ekman, M.; Gudey, S.K.; Yakymovych, M.; Hermansson, A.; Dimitriou, H.; Bengoechea-Alonso, M.T.; Ericsson, J.; et al. TRAF6 ubiquitinates TGFbeta type I receptor to promote its cleavage and nuclear translocation in cancer. Nat. Commun. 2011, 2, 330. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, S.E.; Harikumar, K.B.; Hait, N.C.; Allegood, J.; Strub, G.M.; Kim, E.Y.; Maceyka, M.; Jiang, H.; Luo, C.; Kordula, T.; et al. Sphingosine-1-phosphate is a missing cofactor for the E3 ubiquitin ligase TRAF2. Nature 2010, 465, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Vallabhapurapu, S.; Matsuzawa, A.; Zhang, W.; Tseng, P.H.; Keats, J.J.; Wang, H.; Vignali, D.A.; Bergsagel, P.L.; Karin, M. Nonredundant and complementary functions of TRAF2 and TRAF3 in a ubiquitination cascade that activates NIK-dependent alternative NF-kappaB signaling. Nat. Immunol. 2008, 9, 1364–1370. [Google Scholar] [CrossRef]
- Zapata, J.M.; Reed, J.C. TRAF1: Lord without a RING. Sci. STKE 2002, 2002, PE27. [Google Scholar] [CrossRef] [PubMed]
- Duckett, C.S.; Gedrich, R.W.; Gilfillan, M.C.; Thompson, C.B. Induction of nuclear factor kappaB by the CD30 receptor is mediated by TRAF1 and TRAF2. Mol. Cell Biol. 1997, 17, 1535–1542. [Google Scholar] [CrossRef]
- Wang, C.Y.; Mayo, M.W.; Korneluk, R.G.; Goeddel, D.V.; Baldwin, A.S., Jr. NF-kappaB antiapoptosis: Induction of TRAF1 and TRAF2 and c-IAP1 and c- IAP2 to suppress caspase-8 activation. Science 1998, 281, 1680–1683. [Google Scholar] [CrossRef]
- Zheng, C.; Kabaleeswaran, V.; Wang, Y.; Cheng, G.; Wu, H. Crystal structures of the TRAF2: cIAP2 and the TRAF1: TRAF2: cIAP2 complexes: Affinity, specificity, and regulation. Mol. Cell 2010, 38, 101–113. [Google Scholar] [CrossRef]
- Lu, Y.Y.; Li, Z.Z.; Jiang, D.S.; Wang, L.; Zhang, Y.; Chen, K.; Zhang, X.F.; Liu, Y.; Fan, G.C.; Chen, Y.; et al. TRAF1 is a critical regulator of cerebral ischaemia-reperfusion injury and neuronal death. Nat. Commun. 2013, 4, 2852. [Google Scholar] [CrossRef]
- Xiang, M.; Wang, P.X.; Wang, A.B.; Zhang, X.J.; Zhang, Y.; Zhang, P.; Mei, F.H.; Chen, M.H.; Li, H. Targeting hepatic TRAF1-ASK1 signaling to improve inflammation, insulin resistance, and hepatic steatosis. J. Hepatol. 2016, 64, 1365–1377. [Google Scholar] [CrossRef]
- Zapata, J.M.; Lefebvre, S.; Reed, J.C. Targeting TRAfs for therapeutic intervention. Adv. Exp. Med. Biol. 2007, 597, 188–201. [Google Scholar] [CrossRef]
- Arthur, J.F.; Shen, Y.; Gardiner, E.E.; Coleman, L.; Murphy, D.; Kenny, D.; Andrews, R.K.; Berndt, M.C. TNF receptor-associated factor 4 (TRAF4) is a novel binding partner of glycoprotein Ib and glycoprotein VI in human platelets. J. Thromb. Haemost. JTH 2011, 9, 163–172. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, F.; Garcia de Vinuesa, A.; de Kruijf, E.M.; Mesker, W.E.; Hui, L.; Drabsch, Y.; Li, Y.; Bauer, A.; Rousseau, A.; et al. TRAF4 promotes TGF-beta receptor signaling and drives breast cancer metastasis. Mol. Cell 2013, 51, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Starczynowski, D.T.; Lockwood, W.W.; Delehouzee, S.; Chari, R.; Wegrzyn, J.; Fuller, M.; Tsao, M.S.; Lam, S.; Gazdar, A.F.; Lam, W.L.; et al. TRAF6 is an amplified oncogene bridging the RAS and NF-kappaB pathways in human lung cancer. J. Clin. Investig. 2011, 121, 4095–4105. [Google Scholar] [CrossRef] [PubMed]
- Xie, P. TRAF molecules in cell signaling and in human diseases. J. Mol. Signal. 2013, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.C.; Burkitt, V.; Villa, A.R.; Tong, L.; Wu, H. Structural basis for self-association and receptor recognition of human TRAF2. Nature 1999, 398, 533–538. [Google Scholar] [CrossRef]
- Ye, H.; Arron, J.R.; Lamothe, B.; Cirilli, M.; Kobayashi, T.; Shevde, N.K.; Segal, D.; Dzivenu, O.K.; Vologodskaia, M.; Yim, M.; et al. Distinct molecular mechanism for initiating TRAF6 signalling. Nature 2002, 418, 443–447. [Google Scholar] [CrossRef]
- Ni, C.Z.; Welsh, K.; Leo, E.; Chiou, C.K.; Wu, H.; Reed, J.C.; Ely, K.R. Molecular basis for CD40 signaling mediated by TRAF3. Proc. Natl. Acad. Sci. USA 2000, 97, 10395–10399. [Google Scholar] [CrossRef]
- Kim, C.M.; Choi, J.Y.; Bhat, E.A.; Jeong, J.H.; Son, Y.J.; Kim, S.; Park, H.H. Crystal structure of TRAF1 TRAF domain and its implications in the TRAF1-mediated intracellular signaling pathway. Sci. Rep. 2016, 6, 25526. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, A.; McEwen, A.G.; Poussin-Courmontagne, P.; Rognan, D.; Nomine, Y.; Rio, M.C.; Tomasetto, C.; Alpy, F. TRAF4 is a novel phosphoinositide-binding protein modulating tight junctions and favoring cell migration. PLoS Biol. 2013, 11, e1001726. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Cho, Y.J.; Park, H.H. Structure of the TRAF4 TRAF domain with a coiled-coil domain and its implications for the TRAF4 signalling pathway. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Niu, F.; Ru, H.; Ding, W.; Ouyang, S.; Liu, Z.J. Structural biology study of human TNF receptor associated factor 4 TRAF domain. Protein Cell 2013, 4, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Reichardt, A.; Liang, H.H.; Aliyari, R.; Cheng, D.; Wang, Y.Y.; Xu, F.; Cheng, G.H.; Liu, Y.F. Single Amino Acid Substitutions Confer the Antiviral Activity of the TRAF3 Adaptor Protein onto TRAF5. Sci. Signal 2012, 5, ra81. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.M.; Son, Y.J.; Kim, S.; Kim, S.Y.; Park, H.H. Molecular basis for unique specificity of human TRAF4 for platelets GPIbbeta and GPVI. Proc. Natl. Acad. Sci. USA 2017, 114, 11422–11427. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.Y.; Park, Y.C.; Ye, H.; Wu, H. All TRAFs are not created equal: Common and distinct molecular mechanisms of TRAF-mediated signal transduction. J. Cell Sci. 2002, 115, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H. Structure of TRAF Family: Current Understanding of Receptor Recognition. Front. Immunol. 2018, 9, 1999. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Park, Y.C.; Kreishman, M.; Kieff, E.; Wu, H. The structural basis for the recognition of diverse receptor sequences by TRAF2. Mol. Cell 1999, 4, 321–330. [Google Scholar] [CrossRef]
- Park, Y.C.; Ye, H.; Hsia, C.; Segal, D.; Rich, R.L.; Liou, H.-L.; Myszka, D.G.; Wu, H. A novel mechanism of TRAF signaling revealed by structural and functional analyses of the TRADD-TRAF2 interaction. Cell 2000, 101, 777–787. [Google Scholar] [CrossRef]
- Li, C.; Ni, C.Z.; Havert, M.L.; Cabezas, E.; He, J.; Kaiser, D.; Reed, J.C.; Satterthwait, A.C.; Cheng, G.; Ely, K.R. Downstream regulator TANK binds to the CD40 recognition site on TRAF3. Structure 2002, 10, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.M.; Jeong, J.H.; Son, Y.J.; Choi, J.H.; Kim, S.; Park, H.H. Molecular basis for TANK recognition by TRAF1 revealed by the crystal structure of TRAF1/TANK complex. FEBS Lett. 2017, 591, 810–821. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, I.; Beyaert, R. TRAF1 is a TNF inducible regulator of NF-kappaB activation. FEBS Lett. 1999, 460, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Choi, Y. TRAF1 and its biological functions. Adv. Exp. Med. Biol. 2007, 597, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Greenfeld, H.; Takasaki, K.; Walsh, M.J.; Ersing, I.; Bernhardt, K.; Ma, Y.; Fu, B.; Ashbaugh, C.W.; Cabo, J.; Mollo, S.B.; et al. TRAF1 Coordinates Polyubiquitin Signaling to Enhance Epstein-Barr Virus LMP1-Mediated Growth and Survival Pathway Activation. PLoS Pathog. 2015, 11, e1004890. [Google Scholar] [CrossRef] [PubMed]
- Sabbagh, L.; Pulle, G.; Liu, Y.; Tsitsikov, E.N.; Watts, T.H. ERK-dependent Bim modulation downstream of the 4-1BB-TRAF1 signaling axis is a critical mediator of CD8 T cell survival in vivo. J. Immunol. 2008, 180, 8093–8101. [Google Scholar] [CrossRef] [PubMed]
- Nocentini, G.; Riccardi, C. GITR: A modulator of immune response and inflammation. Adv. Exp. Med. Biol. 2009, 647, 156–173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Zhang, R.; Huang, L.; Wang, P.X.; Zhang, Y.; Jiang, D.S.; Zhu, L.H.; Tian, S.; Zhang, X.D.; Li, H. TRAF1 is a key mediator for hepatic ischemia/reperfusion injury. Cell Death Dis. 2014, 5, e1467. [Google Scholar] [CrossRef] [PubMed]
- Abate, F.; Todaro, M.; van der Krogt, J.A.; Boi, M.; Landra, I.; Machiorlatti, R.; Tabbo, F.; Messana, K.; Abele, C.; Barreca, A.; et al. A novel patient-derived tumorgraft model with TRAF1-ALK anaplastic large-cell lymphoma translocation. Leukemia 2015, 29, 1390–1401. [Google Scholar] [CrossRef]
- Wu, S.; Xie, P.; Welsh, K.; Li, C.; Ni, C.Z.; Zhu, X.; Reed, J.C.; Satterthwait, A.C.; Bishop, G.A.; Ely, K.R. LMP1 protein from the Epstein-Barr virus is a structural CD40 decoy in B lymphocytes for binding to TRAF3. J. Biol. Chem. 2005, 280, 33620–33626. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, Z.; Zhang, Z.; Wang, Y.; Li, C.; Wang, X.; He, F.; Sun, L.; Jiao, S.; Shi, W.; et al. Structural Insights into mitochondrial antiviral signaling protein (MAVS)-tumor necrosis factor receptor-associated factor 6 (TRAF6) signaling. J. Biol. Chem. 2015, 290, 26811–26820. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, H.; Kim, S.; Kim, D.Y.; Han, J.H.; Park, H.H. TRAF1 from a Structural Perspective. Biomolecules 2024, 14, 510. https://doi.org/10.3390/biom14050510
Jang H, Kim S, Kim DY, Han JH, Park HH. TRAF1 from a Structural Perspective. Biomolecules. 2024; 14(5):510. https://doi.org/10.3390/biom14050510
Chicago/Turabian StyleJang, Hyunseok, Subin Kim, Do Yeon Kim, Ju Hee Han, and Hyun Ho Park. 2024. "TRAF1 from a Structural Perspective" Biomolecules 14, no. 5: 510. https://doi.org/10.3390/biom14050510
APA StyleJang, H., Kim, S., Kim, D. Y., Han, J. H., & Park, H. H. (2024). TRAF1 from a Structural Perspective. Biomolecules, 14(5), 510. https://doi.org/10.3390/biom14050510