
Caspase-5: Structure, Pro-Inflammatory Activity and Evolution

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction: Caspases Are Critical Regulators of Apoptosis and Pyroptosis
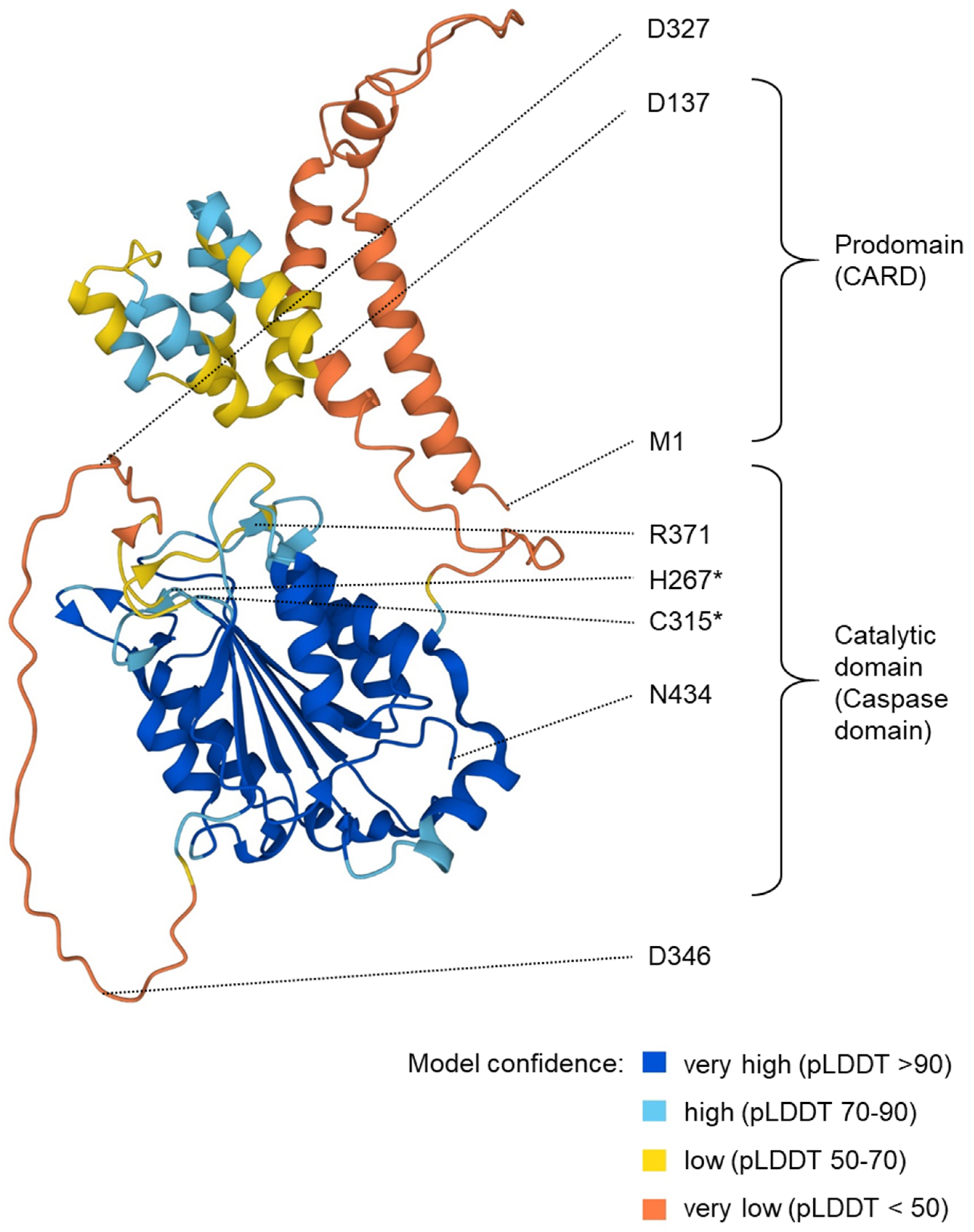
2. The Structure of Caspase-5 Gene and Proteins
3. The Function of Caspase-5 in the Response to Gram-Negative Bacteria
3.1. LPS Regulates the Expression of Caspase-5
3.2. LPS Activates Caspase-5 in the Non-Canonical Inflammasome
3.3. Mechanisms of Sensing Cytosolic LPS and Oxidized Phospholipids
3.4. Substrates of Caspase-5
4. The Evolution of Caspase-5 and Related Caspases
5. Roles of Caspase-5 in Human Diseases
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Salvesen, G.S.; Ashkenazi, A. Snapshot: Caspases. Cell 2011, 147, 1197. [Google Scholar] [CrossRef]
- Van Opdenbosch, N.; Lamkanfi, M. Caspases in cell death, inflammation, and disease. Immunity 2019, 50, 1352–1364. [Google Scholar] [CrossRef]
- Fuentes-Prior, P.; Salvesen, G.S. The protein structures that shape caspase activity, specificity, activation and inhibition. Biochem. J. 2004, 384 Pt 2, 201–232. [Google Scholar] [CrossRef]
- Green, D.R. Caspases and Their Substrates. Cold Spring Harb. Perspect. Biol. 2022, 14, a041012. [Google Scholar] [CrossRef]
- Eckhart, L.; Ballaun, C.; Hermann, M.; VandeBerg, J.L.; Sipos, W.; Uthman, A.; Fischer, H.; Tschachler, E. Identification of novel mammalian caspases reveals an important role of gene loss in shaping the human caspase repertoire. Mol. Biol. Evol. 2008, 25, 831–841. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Van de Craen, M.; Van Loo, G.; Pype, S.; Van Criekinge, W.; Van den brande, I.; Molemans, F.; Fiers, W.; Declercq, W.; Vandenabeele, P. Identification of a new caspase homologue: Caspase-14. Cell Death Differ. 1998, 5, 838–846. [Google Scholar] [CrossRef]
- Newton, K.; Strasser, A.; Kayagaki, N.; Dixit, V.M. Cell death. Cell 2024, 187, 235–256. [Google Scholar] [CrossRef]
- Fischer, H.; Rossiter, H.; Ghannadan, M.; Jaeger, K.; Barresi, C.; Declercq, W.; Tschachler, E.; Eckhart, L. Caspase-14 but not caspase-3 is processed during the development of fetal mouse epidermis. Differentiation 2005, 73, 406–413. [Google Scholar] [CrossRef]
- Denecker, G.; Hoste, E.; Gilbert, B.; Hochepied, T.; Ovaere, P.; Lippens, S.; Van den Broecke, C.; Van Damme, P.; D’Herde, K.; Hachem, J.P.; et al. Caspase-14 protects against epidermal UVB photodamage and water loss. Nat. Cell Biol. 2007, 9, 666–674. [Google Scholar] [CrossRef]
- Irmler, M.; Thome, M.; Hahne, M.; Schneider, P.; Hofmann, K.; Steiner, V.; Bodmer, J.L.; Schröter, M.; Burns, K.; Mattmann, C.; et al. Inhibition of death receptor signals by cellular FLIP. Nature 1997, 388, 190–195. [Google Scholar] [CrossRef]
- Julien, O.; Wells, J.A. Caspases and their substrates. Cell Death Differ. 2017, 24, 1380–1389. [Google Scholar] [CrossRef]
- Bibo-Verdugo, B.; Snipas, S.J.; Kolt, S.; Poreba, M.; Salvesen, G.S. Extended subsite profiling of the pyroptosis effector protein gasdermin D reveals a region recognized by inflammatory caspase-11. J. Biol. Chem. 2020, 295, 11292–11302. [Google Scholar] [CrossRef]
- Exconde, P.M.; Hernandez-Chavez, C.; Bourne, C.M.; Richards, R.M.; Bray, M.B.; Lopez, J.L.; Srivastava, T.; Egan, M.S.; Zhang, J.; Yoo, W.; et al. The tetrapeptide sequence of IL-18 and IL-1β regulates their recruitment and activation by inflammatory caspases. Cell Rep. 2023, 42, 113581. [Google Scholar] [CrossRef]
- Green, D.R. Inflammasomes and Other Caspase-Activation Platforms. Cold Spring Harb. Perspect. Biol. 2022, 14, a041061. [Google Scholar] [CrossRef]
- Green, D.R. The Death Receptor Pathway of Apoptosis. Cold Spring Harb. Perspect. Biol. 2022, 14, a041053. [Google Scholar] [CrossRef]
- Green, D.R. The Mitochondrial Pathway of Apoptosis: Part I: MOMP and Beyond. Cold Spring Harb. Perspect. Biol. 2022, 14, a041038. [Google Scholar] [CrossRef]
- Van de Craen, M.; Declercq, W.; Van den brande, I.; Fiers, W.; Vandenabeele, P. The proteolytic procaspase activation network: An in vitro analysis. Cell Death Differ. 1999, 6, 1117–1124. [Google Scholar] [CrossRef]
- Creagh, E.M.; Martin, S.J. Caspases: Cellular demolition experts. Biochem. Soc. Trans. 2001, 29 Pt 6, 696–702. [Google Scholar] [CrossRef]
- Mikolajczyk, J.; Scott, F.L.; Krajewski, S.; Sutherlin, D.P.; Salvesen, G.S. Activation and substrate specificity of caspase-14. Biochemistry 2004, 43, 10560–10569. [Google Scholar] [CrossRef]
- Fischer, H.; Stichenwirth, M.; Dockal, M.; Ghannadan, M.; Buchberger, M.; Bach, J.; Kapetanopoulos, A.; Declercq, W.; Tschachler, E.; Eckhart, L. Stratum corneum-derived caspase-14 is catalytically active. FEBS Lett. 2004, 577, 446–450. [Google Scholar] [CrossRef]
- Yuan, J.; Ofengeim, D. A guide to cell death pathways. Nat. Rev. Mol. Cell Biol. 2024, 25, 379–395. [Google Scholar] [CrossRef] [PubMed]
- Kayagaki, N.; Webster, J.D.; Newton, K. Control of Cell Death in Health and Disease. Annu. Rev. Pathol. 2024, 19, 157–180. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xu, P. Activation and pharmacological regulation of inflammasomes. Biomolecules 2022, 12, 1005. [Google Scholar] [CrossRef]
- Agnew, A.; Nulty, C.; Creagh, E.M. Regulation, activation and function of caspase-11 during health and disease. Int. J. Mol. Sci. 2021, 22, 1506. [Google Scholar] [CrossRef]
- Munday, N.A.; Vaillancourt, J.P.; Ali, A.; Casano, F.J.; Miller, D.K.; Molineaux, S.M.; Yamin, T.T.; Yu, V.L.; Nicholson, D.W. Molecular cloning and pro-apoptotic activity of ICErelII and ICErelIII, members of the ICE/CED-3 family of cysteine proteases. J. Biol. Chem. 1995, 270, 15870–15876. [Google Scholar] [CrossRef]
- Faucheu, C.; Blanchet, A.M.; Collard-Dutilleul, V.; Lalanne, J.L.; Diu-Hercend, A. Identification of a cysteine protease closely related to interleukin-1 beta-converting enzyme. Eur. J. Biochem. 1996, 236, 207–213. [Google Scholar] [CrossRef]
- Eckhart, L.; Kittel, C.; Gawlas, S.; Gruber, F.; Mildner, M.; Jilma, B.; Tschachler, E. Identification of a novel exon encoding the amino-terminus of the predominant caspase-5 variants. Biochem. Biophys. Res. Commun. 2006, 348, 682–688. [Google Scholar] [CrossRef]
- Monaghan, L.; Longman, D.; Cáceres, J.F. Translation-coupled mRNA quality control mechanisms. EMBO J. 2023, 42, e114378. [Google Scholar] [CrossRef]
- Schauvliege, R.; Vanrobaeys, J.; Schotte, P.; Beyaert, R. Caspase-11 gene expression in response to lipopolysaccharide and interferon-gamma requires nuclear factor-kappa B and signal transducer and activator of transcription (STAT) 1. J. Biol. Chem. 2002, 277, 41624–41630. [Google Scholar] [CrossRef]
- Lin, X.Y.; Choi, M.S.; Porter, A.G. Expression analysis of the human caspase-1 subfamily reveals specific regulation of the CASP5 gene by lipopolysaccharide and interferon-gamma. J. Biol. Chem. 2000, 275, 39920–39926. [Google Scholar] [CrossRef]
- Lagrange, B.; Benaoudia, S.; Wallet, P.; Magnotti, F.; Provost, A.; Michal, F.; Martin, A.; Di Lorenzo, F.; Py, B.F.; Molinaro, A.; et al. Human caspase-4 detects tetra-acylated LPS and cytosolic Francisella and functions differently from murine caspase-11. Nat. Commun. 2018, 9, 242. [Google Scholar] [CrossRef]
- Ghait, M.; Duduskar, S.N.; Rooney, M.; Häfner, N.; Reng, L.; Göhrig, B.; Reuken, P.A.; Bloos, F.; Bauer, M.; Sponholz, C.; et al. The non-canonical inflammasome activators Caspase-4 and Caspase-5 are differentially regulated during immunosuppression-associated organ damage. Front. Immunol. 2023, 14, 1239474. [Google Scholar] [CrossRef]
- Kayagaki, N.; Warming, S.; Lamkanfi, M.; Vande Walle, L.; Louie, S.; Dong, J.; Newton, K.; Qu, Y.; Liu, J.; Heldens, S.; et al. Non-canonical inflammasome activation targets caspase-11. Nature 2011, 479, 117–121. [Google Scholar] [CrossRef]
- Downs, K.P.; Nguyen, H.; Dorfleutner, A.; Stehlik, C. An overview of the non-canonical inflammasome. Mol. Asp. Med. 2020, 76, 100924. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.; Wang, Y.; Gao, W.; Ding, J.; Li, P.; Hu, L.; Shao, F. Inflammatory caspases are innate immune receptors for intracellular LPS. Nature 2014, 514, 187–192. [Google Scholar] [CrossRef]
- Viganò, E.; Diamond, C.E.; Spreafico, R.; Balachander, A.; Sobota, R.M.; Mortellaro, A. Human caspase-4 and caspase-5 regulate the one-step non-canonical inflammasome activation in monocytes. Nat. Commun. 2015, 6, 8761. [Google Scholar] [CrossRef]
- Wandel, M.P.; Kim, B.H.; Park, E.S.; Boyle, K.B.; Nayak, K.; Lagrange, B.; Herod, A.; Henry, T.; Zilbauer, M.; Rohde, J.; et al. Guanylate-binding proteins convert cytosolic bacteria into caspase-4 signaling platforms. Nat. Immunol. 2020, 21, 880–891. [Google Scholar] [CrossRef]
- Santos, J.C.; Boucher, D.; Schneider, L.K.; Demarco, B.; Dilucca, M.; Shkarina, K.; Heilig, R.; Chen, K.W.; Lim, R.Y.H.; Broz, P. Human GBP1 binds LPS to initiate assembly of a caspase-4 activating platform on cytosolic bacteria. Nat. Commun. 2020, 11, 3276. [Google Scholar] [CrossRef]
- Zamyatina, A.; Heine, H. Lipopolysaccharide Recognition in the Crossroads of TLR4 and Caspase-4/11 Mediated Inflammatory Pathways. Front. Immunol. 2020, 11, 585146. [Google Scholar] [CrossRef]
- Barnett, K.C.; Li, S.; Liang, K.; Ting, J.P. A 360° view of the inflammasome: Mechanisms of activation, cell death, and diseases. Cell 2023, 186, 2288–2312. [Google Scholar] [CrossRef]
- Akuma, D.C.; Wodzanowski, K.A.; Schwartz Wertman, R.; Exconde, P.M.; Vázquez Marrero, V.R.; Odunze, C.E.; Grubaugh, D.; Shin, S.; Taabazuing, C.; Brodsky, I.E. Catalytic activity and autoprocessing of murine caspase-11 mediate noncanonical inflammasome assembly in response to cytosolic LPS. Elife 2024, 13, e83725. [Google Scholar] [CrossRef]
- Kayagaki, N.; Stowe, I.B.; Lee, B.L.; O’Rourke, K.; Anderson, K.; Warming, S.; Cuellar, T.; Haley, B.; Roose-Girma, M.; Phung, Q.T.; et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 2015, 526, 666–671. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef]
- Lee, B.L.; Stowe, I.B.; Gupta, A.; Kornfeld, O.S.; Roose-Girma, M.; Anderson, K.; Warming, S.; Zhang, J.; Lee, W.P.; Kayagaki, N. Caspase-11 auto-proteolysis is crucial for noncanonical inflammasome activation. J. Exp. Med. 2018, 215, 2279–2288. [Google Scholar] [CrossRef]
- Fu, J.; Schroder, K.; Wu, H. Mechanistic insights from inflammasome structures. Nat. Rev. Immunol. 2024. [Google Scholar] [CrossRef]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Calabrese, L.; Fiocco, Z.; Mellett, M.; Aoki, R.; Rubegni, P.; French, L.E.; Satoh, T.K. Role of the NLRP1 inflammasome in skin cancer and inflammatory skin diseases. Br. J. Dermatol. 2024, 190, 305–315. [Google Scholar] [CrossRef]
- Rojas-Lopez, M.; Gil-Marqués, M.L.; Kharbanda, V.; Zajac, A.S.; Miller, K.A.; Wood, T.E.; Hachey, A.C.; Egger, K.T.; Goldberg, M.B. NLRP11 is a pattern recognition receptor for bacterial lipopolysaccharide in the cytosol of human macrophages. Sci. Immunol. 2023, 8, eabo4767. [Google Scholar] [CrossRef]
- Salova, M.; Sipos, W.; Tschachler, E.; Eckhart, L. NOD2 and reproduction-associated NOD-like receptors have been lost during the evolution of pangolins. Immunogenetics 2022, 74, 261–268. [Google Scholar] [CrossRef]
- Moretti, J.; Jia, B.; Hutchins, Z.; Roy, S.; Yip, H.; Wu, J.; Shan, M.; Jaffrey, S.R.; Coers, J.; Blander, J.M. Caspase-11 interaction with NLRP3 potentiates the noncanonical activation of the NLRP3 inflammasome. Nat. Immunol. 2022, 23, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Ma, J.; Li, W.; Liu, Q.; Qin, X.; Qian, Y.; Wang, C.; Zhang, Y.; Li, Y.; Jiang, D.; et al. The orphan receptor Nur77 binds cytoplasmic LPS to activate the non-canonical NLRP3 inflammasome. Immunity 2023, 56, 753–767.e8. [Google Scholar] [CrossRef] [PubMed]
- Colaço-Gaspar, M.; Hofer, P.; Oberer, M.; Zechner, R. PNPLA-mediated lipid hydrolysis and transacylation—At the intersection of catabolism and anabolism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2024, 1869, 159410. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, Q.; Qian, Y.; Wang, C.; Kong, C.; Sun, L.; Sun, L.; Liu, H.; Zhang, Y.; Jiang, D.; et al. Adipose triglyceride lipase suppresses noncanonical inflammasome by hydrolyzing LPS. Nat. Chem. Biol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, I.; Tan, Y.; Di Gioia, M.; Broggi, A.; Ruan, J.; Shi, J.; Donado, C.A.; Shao, F.; Wu, H.; Springstead, J.R.; et al. An endogenous caspase-11 ligand elicits interleukin-1 release from living dendritic cells. Science 2016, 352, 1232–1236. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.H.; Indramohan, M.; Ratsimandresy, R.A.; Gangopadhyay, A.; Morris, E.P.; Monack, D.M.; Dorfleutner, A.; Stehlik, C. The oxidized phospholipid oxPAPC protects from septic shock by targeting the non-canonical inflammasome in macrophages. Nat. Commun. 2018, 9, 996. [Google Scholar] [CrossRef]
- Bochkov, V.N.; Kadl, A.; Huber, J.; Gruber, F.; Binder, B.R.; Leitinger, N. Protective role of phospholipid oxidation products in endotoxin-induced tissue damage. Nature 2002, 419, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Di Gioia, M.; Zanoni, I. Dooming Phagocyte Responses: Inflammatory Effects of Endogenous Oxidized Phospholipids. Front. Endocrinol. 2021, 12, 626842. [Google Scholar] [CrossRef]
- Casson, C.N.; Yu, J.; Reyes, V.M.; Taschuk, F.O.; Yadav, A.; Copenhaver, A.M.; Nguyen, H.T.; Collman, R.G.; Shin, S. Human caspase-4 mediates noncanonical inflammasome activation against gram-negative bacterial pathogens. Proc. Natl. Acad. Sci. USA 2015, 112, 6688–6693. [Google Scholar] [CrossRef]
- Baker, P.J.; Boucher, D.; Bierschenk, D.; Tebartz, C.; Whitney, P.G.; D’Silva, D.B.; Tanzer, M.C.; Monteleone, M.; Robertson, A.A.; Cooper, M.A.; et al. NLRP3 inflammasome activation downstream of cytoplasmic LPS recognition by both caspase-4 and caspase-5. Eur. J. Immunol. 2015, 45, 2918–2926. [Google Scholar] [CrossRef]
- Wiggins, K.A.; Parry, A.J.; Cassidy, L.D.; Humphry, M.; Webster, S.J.; Goodall, J.C.; Narita, M.; Clarke, M.C.H. IL-1α cleavage by inflammatory caspases of the noncanonical inflammasome controls the senescence-associated secretory phenotype. Aging Cell 2019, 18, e12946. [Google Scholar] [CrossRef] [PubMed]
- Davies, C.W.; Stowe, I.; Phung, Q.T.; Ho, H.; Bakalarski, C.E.; Gupta, A.; Zhang, Y.; Lill, J.R.; Payandeh, J.; Kayagaki, N.; et al. Discovery of a caspase cleavage motif antibody reveals insights into noncanonical inflammasome function. Proc. Natl. Acad. Sci. USA 2021, 118, e2018024118. [Google Scholar] [CrossRef] [PubMed]
- Krippner-Heidenreich, A.; Talanian, R.V.; Sekul, R.; Kraft, R.; Thole, H.; Ottleben, H.; Lüscher, B. Targeting of the transcription factor Max during apoptosis: Phosphorylation-regulated cleavage by caspase-5 at an unusual glutamic acid residue in position P1. Biochem. J. 2001, 358 Pt 3, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Miura, M.; Jung, Y.K.; Zhu, H.; Li, E.; Yuan, J. Murine caspase-11, an ICE-interacting protease, is essential for the activation of ICE. Cell 1998, 92, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Koenig, U.; Eckhart, L.; Tschachler, E. Evidence that caspase-13 is not a human but a bovine gene. Biochem. Biophys. Res. Commun. 2001, 285, 1150–1154. [Google Scholar] [CrossRef] [PubMed]
- Devant, P.; Cao, A.; Kagan, J.C. Evolution-inspired redesign of the LPS receptor caspase-4 into an interleukin-1β converting enzyme. Sci. Immunol. 2021, 6, eabh3567. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Suleski, M.; Craig, J.M.; Kasprowicz, A.E.; Sanderford, M.; Li, M.; Stecher, G.; Hedges, S.B. TimeTree 5: An expanded resource for species divergence times. Mol. Biol. Evol. 2022, 39, msac174. [Google Scholar] [CrossRef] [PubMed]
- Digby, Z.; Tourlomousis, P.; Rooney, J.; Boyle, J.P.; Bibo-Verdugo, B.; Pickering, R.J.; Webster, S.J.; Monie, T.P.; Hopkins, L.J.; Kayagaki, N.; et al. Evolutionary loss of inflammasomes in the Carnivora and implications for the carriage of zoonotic infections. Cell Rep. 2021, 36, 109614. [Google Scholar] [CrossRef]
- Yang, D.; Zheng, X.; Chen, S.; Wang, Z.; Xu, W.; Tan, J.; Hu, T.; Hou, M.; Wang, W.; Gu, Z.; et al. Sensing of cytosolic LPS through caspy2 pyrin domain mediates noncanonical inflammasome activation in zebrafish. Nat. Commun. 2018, 9, 3052. [Google Scholar] [CrossRef]
- Eckhart, L.; Ballaun, C.; Uthman, A.; Kittel, C.; Stichenwirth, M.; Buchberger, M.; Fischer, H.; Sipos, W.; Tschachler, E. Identification and characterization of a novel mammalian caspase with proapoptotic activity. J. Biol. Chem. 2005, 280, 35077–35080. [Google Scholar] [CrossRef]
- Kalai, M.; Lamkanfi, M.; Denecker, G.; Boogmans, M.; Lippens, S.; Meeus, A.; Declercq, W.; Vandenabeele, P. Regulation of the expression and processing of caspase-12. J. Cell Biol. 2003, 162, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Eckhart, L.; Ballaun, C.; Uthman, A.; Gawlas, S.; Buchberger, M.; Fischer, H.; Tschachler, E. Duplication of the caspase-12 prodomain and inactivation of NLRC4/IPAF in the dog. Biochem. Biophys. Res. Commun. 2009, 384, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.; Koenig, U.; Eckhart, L.; Tschachler, E. Human caspase 12 has acquired deleterious mutations. Biochem. Biophys. Res. Commun. 2002, 293, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, V.A.K.; Zhao, Y.; Shao, F. Innate immunity to intracellular LPS. Nat. Immunol. 2019, 20, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.P.; Creagh, E.M. Caspase-4 and -5 biology in the pathogenesis of inflammatory bowel disease. Front. Pharmacol. 2022, 13, 919567. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Russo, A.J.; Wright, S.S.; Muthupalani, S.; Rathinam, V.A. Hierarchical cell-type-specific functions of caspase-11 in LPS shock and antibacterial host defense. Cell Rep. 2021, 35, 109012. [Google Scholar] [CrossRef] [PubMed]
- Kajiwara, Y.; Schiff, T.; Voloudakis, G.; Gama Sosa, M.A.; Elder, G.; Bozdagi, O.; Buxbaum, J.D. A critical role for human caspase-4 in endotoxin sensitivity. J. Immunol. 2014, 193, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Flood, B.; Oficjalska, K.; Laukens, D.; Fay, J.; O’Grady, A.; Caiazza, F.; Heetun, Z.; Mills, K.H.; Sheahan, K.; Ryan, E.J.; et al. Altered expression of caspases-4 and -5 during inflammatory bowel disease and colorectal cancer: Diagnostic and therapeutic potential. Clin. Exp. Immunol. 2015, 181, 39–50. [Google Scholar] [CrossRef]
- Matikainen, S.; Nyman, T.A.; Cypryk, W. Function and Regulation of Noncanonical Caspase-4/5/11 Inflammasome. J. Immunol. 2020, 204, 3063–3069. [Google Scholar] [CrossRef]
- Akhter, A.; Caution, K.; Abu Khweek, A.; Tazi, M.; Abdulrahman, B.A.; Abdelaziz, D.H.; Voss, O.H.; Doseff, A.I.; Hassan, H.; Azad, A.K.; et al. Caspase-11 promotes the fusion of phagosomes harboring pathogenic bacteria with lysosomes by modulating actin polymerization. Immunity 2012, 37, 35–47. [Google Scholar] [CrossRef]
- Wang, X.; Ni, J.; You, Y.; Feng, G.; Zhang, S.; Bao, W.; Hou, H.; Li, H.; Liu, L.; Zheng, M.; et al. SNX10-mediated LPS sensing causes intestinal barrier dysfunction via a caspase-5-dependent signaling cascade. EMBO J. 2021, 40, e108080. [Google Scholar] [CrossRef]
- Dickinson, M.S.; Coers, J. SNX10 and caspase-5 sort out endosomal LPS for a gut-wrenching Slug-fest. EMBO J. 2021, 40, e110128. [Google Scholar] [CrossRef]
- Bitto, N.J.; Baker, P.J.; Dowling, J.K.; Wray-McCann, G.; De Paoli, A.; Tran, L.S.; Leung, P.L.; Stacey, K.J.; Mansell, A.; Masters, S.L.; et al. Membrane vesicles from Pseudomonas aeruginosa activate the noncanonical inflammasome through caspase-5 in human monocytes. Immunol. Cell Biol. 2018, 96, 1120–1130. [Google Scholar] [CrossRef]
- Sharma, A.K.; Ismail, N. Non-canonical inflammasome pathway: The role of cell death and inflammation in Ehrlichiosis. Cells 2023, 12, 2597. [Google Scholar] [CrossRef]
- Salskov-Iversen, M.L.; Johansen, C.; Kragballe, K.; Iversen, L. Caspase-5 expression is upregulated in lesional psoriatic skin. J. Investig. Dermatol. 2011, 131, 670–676. [Google Scholar] [CrossRef]
- Zwicker, S.; Hattinger, E.; Bureik, D.; Batycka-Baran, A.; Schmidt, A.; Gerber, P.A.; Rothenfusser, S.; Gilliet, M.; Ruzicka, T.; Wolf, R. Th17 micro-milieu regulates NLRP1-dependent caspase-5 activity in skin autoinflammation. PLoS ONE 2017, 12, e0175153. [Google Scholar] [CrossRef]
- Zasłona, Z.; Flis, E.; Wilk, M.M.; Carroll, R.G.; Palsson-McDermott, E.M.; Hughes, M.M.; Diskin, C.; Banahan, K.; Ryan, D.G.; Hooftman, A.; et al. Caspase-11 promotes allergic airway inflammation. Nat. Commun. 2020, 11, 1055. [Google Scholar] [CrossRef]
- Bolívar, B.E.; Brown-Suedel, A.N.; Rohrman, B.A.; Charendoff, C.I.; Yazdani, V.; Belcher, J.D.; Vercellotti, G.M.; Flanagan, J.M.; Bouchier-Hayes, L. Noncanonical Roles of Caspase-4 and Caspase-5 in Heme-Driven IL-1β Release and Cell Death. J. Immunol. 2021, 206, 1878–1889. [Google Scholar] [CrossRef]
- Dutra, F.F.; Alves, L.S.; Rodrigues, D.; Fernandez, P.L.; de Oliveira, R.B.; Golenbock, D.T.; Zamboni, D.S.; Bozza, M.T. Hemolysis-induced lethality involves inflammasome activation by heme. Proc. Natl. Acad. Sci. USA 2014, 111, E4110–E4118. [Google Scholar] [CrossRef]
- Saleh, M.; Vaillancourt, J.P.; Graham, R.K.; Huyck, M.; Srinivasula, S.M.; Alnemri, E.S.; Steinberg, M.H.; Nolan, V.; Baldwin, C.T.; Hotchkiss, R.S.; et al. Differential modulation of endotoxin responsiveness by human caspase-12 polymorphisms. Nature 2004, 429, 75–79. [Google Scholar] [CrossRef]
- Rui, H.; Yan, T.; Hu, Z.; Liu, R.; Wang, L. The association between caspase-5 gene polymorphisms and rheumatoid arthritis in a Chinese population. Gene 2018, 642, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S., Jr.; Yamamoto, H.; Navarro, M.; Maestro, M.; Reventós, J.; Perucho, M. Frameshift mutations at mononucleotide repeats in caspase-5 and other target genes in endometrial and gastrointestinal cancer of the microsatellite mutator phenotype. Cancer Res. 1999, 59, 2995–3002. [Google Scholar] [PubMed]
- Soung, Y.H.; Jeong, E.G.; Ahn, C.H.; Kim, S.S.; Song, S.Y.; Yoo, N.J.; Lee, S.H. Mutational analysis of caspase 1, 4, and 5 genes in common human cancers. Hum. Pathol. 2008, 39, 895–900. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.L.; Liu, W.; Gao, C.X.; Shang, Z.; Ning, L.J.; Liu, X. CASP-1, -2 and -5 gene polymorphisms and cancer risk: A review and meta-analysis. Biomed. Rep. 2013, 1, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Hisama, F.M.; Pillai, R.K.; Sidorova, J.; Patterson, K.; Gokingco, C.; Yacobi-Bach, M.; Oshima, J. Caspase 5 depletion is linked to hyper-inflammatory response and progeroid syndrome. Geroscience 2024, 46, 2771–2775. [Google Scholar] [CrossRef] [PubMed]
- Pfalzgraff, A.; Weindl, G. Intracellular Lipopolysaccharide Sensing as a Potential Therapeutic Target for Sepsis. Trends Pharmacol. Sci. 2019, 40, 187–197. [Google Scholar] [CrossRef]
- Cornelis, S.; Kersse, K.; Festjens, N.; Lamkanfi, M.; Vandenabeele, P. Inflammatory caspases: Targets for novel therapies. Curr. Pharm. Des. 2007, 13, 367–385. [Google Scholar] [CrossRef] [PubMed]
- Dhani, S.; Zhao, Y.; Zhivotovsky, B. A long way to go: Caspase inhibitors in clinical use. Cell Death Dis. 2021, 12, 949. [Google Scholar] [CrossRef]
- Dai, Z.; Liu, W.C.; Chen, X.Y.; Wang, X.; Li, J.L.; Zhang, X. Gasdermin D-mediated pyroptosis: Mechanisms, diseases, and inhibitors. Front. Immunol. 2023, 14, 1178662. [Google Scholar] [CrossRef]
- Kopp, A.; Hagelueken, G.; Jamitzky, I.; Moecking, J.; Schiffelers, L.D.J.; Schmidt, F.I.; Geyer, M. Pyroptosis inhibiting nanobodies block Gasdermin D pore formation. Nat. Commun. 2023, 14, 7923. [Google Scholar] [CrossRef]
- Wei, C.; Jiang, W.; Wang, R.; Zhong, H.; He, H.; Gao, X.; Zhong, S.; Yu, F.; Guo, Q.; Zhang, L.; et al. Brain endothelial GSDMD activation mediates inflammatory BBB breakdown. Nature 2024. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Huang, M.; Yao, Y.M. Recent advances in the biology of IL-1 family cytokines and their potential roles in development of sepsis. Cytokine Growth Factor. Rev. 2019, 45, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Liu, J.; Zeng, L.; Zhu, S.; Wang, H.; Billiar, T.R.; Kroemer, G.; Klionsky, D.J.; Zeh, H.J.; Jiang, J.; et al. Extracellular SQSTM1 mediates bacterial septic death in mice through insulin receptor signalling. Nat. Microbiol. 2020, 5, 1576–1587. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eckhart, L.; Fischer, H. Caspase-5: Structure, Pro-Inflammatory Activity and Evolution. Biomolecules 2024, 14, 520. https://doi.org/10.3390/biom14050520
Eckhart L, Fischer H. Caspase-5: Structure, Pro-Inflammatory Activity and Evolution. Biomolecules. 2024; 14(5):520. https://doi.org/10.3390/biom14050520
Chicago/Turabian StyleEckhart, Leopold, and Heinz Fischer. 2024. "Caspase-5: Structure, Pro-Inflammatory Activity and Evolution" Biomolecules 14, no. 5: 520. https://doi.org/10.3390/biom14050520
APA StyleEckhart, L., & Fischer, H. (2024). Caspase-5: Structure, Pro-Inflammatory Activity and Evolution. Biomolecules, 14(5), 520. https://doi.org/10.3390/biom14050520