


Herpesvirus Entry Mediator as an Immune Checkpoint Target and a Potential Prognostic Biomarker in Myeloid and Lymphoid Leukemia

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. In Vitro Tumor and Healthy Control Cell Lines
2.3. CFSE CD4+ T Cell Proliferation in Response to HVEM Blocker-Treated and Untreated Tumor Cells In Vitro
2.4. Flow Cytometry
2.5. Study Subjects
2.6. Reverse Transcription-Quantitative PCR (RT-qPCR)
2.7. Statistical Analysis
3. Results
3.1. HVEM Gene Expression and HVEM Surface Protein Are Upregulated in Different Cancer Cell Lines In Vitro
3.2. CD4+ T Cell Proliferation in Response to HVEM Blocker-Treated and Untreated Tumor Cells In Vitro
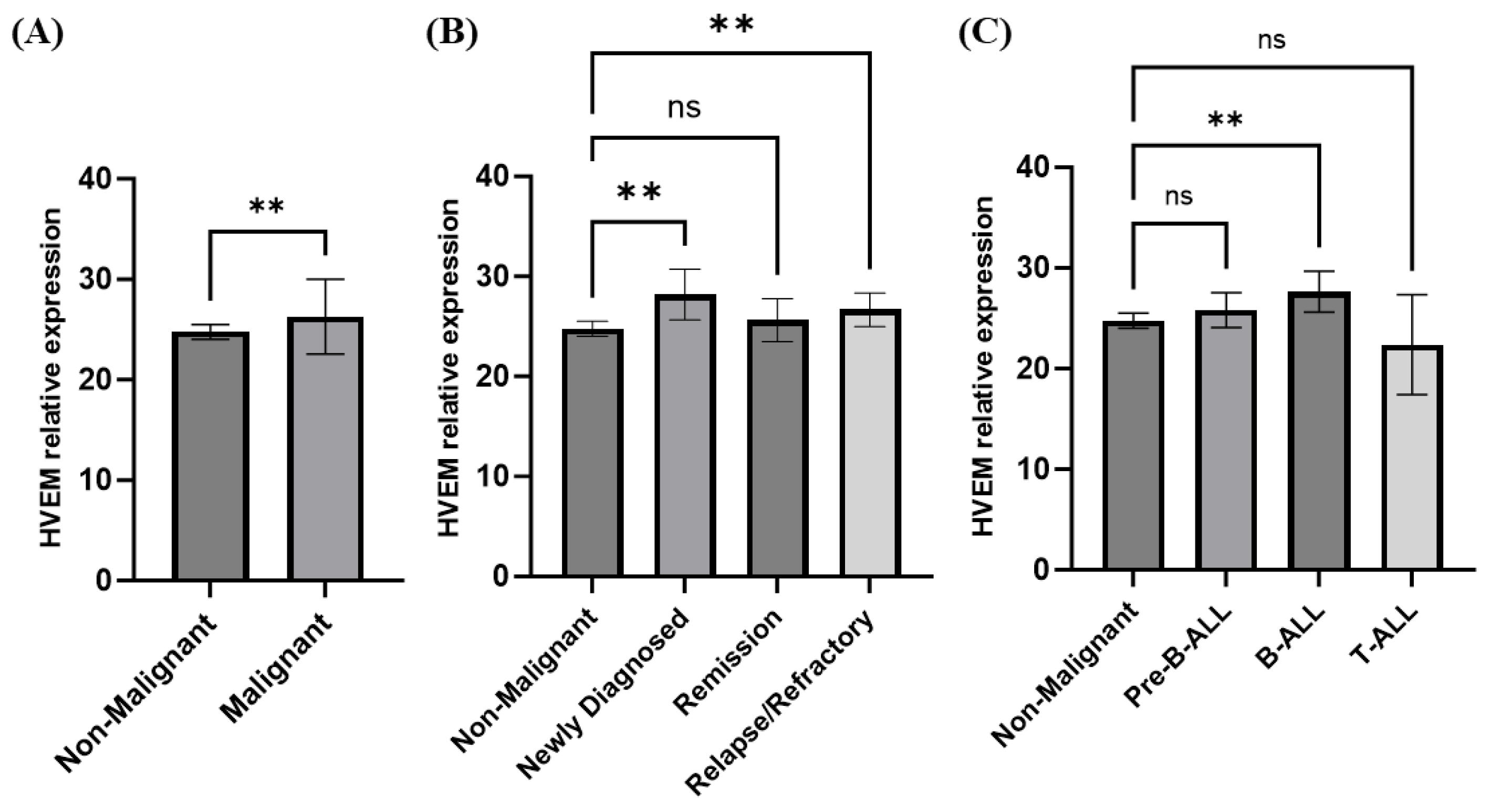
3.3. Elevated HVEM Gene Expression in Acute Lymphocytic Leukemia (ALL)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sordo-Bahamonde, C.; Lorenzo-Herrero, S.; Granda-Díaz, R.; Martínez-Pérez, A.; Aguilar-García, C.; Rodrigo, J.P.; García-Pedrero, J.M.; Gonzalez, S. Beyond the anti-PD-1/PD-L1 era: Promising role of the BTLA/HVEM axis as a future target for cancer immunotherapy. Mol. Cancer 2023, 22, 142. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Siddiqui, B.A.; Anandhan, S.; Yadav, S.S.; Subudhi, S.K.; Gao, J.; Goswami, S.; Allison, J.P. The Next Decade of Immune Checkpoint Therapy. Cancer Discov. 2021, 11, 838–857. [Google Scholar] [CrossRef]
- Wolchok, J.D. Checkpoint blockade: The end of the beginning. Nat. Rev. Immunol. 2021, 21, 621. [Google Scholar] [CrossRef]
- Shui, J.W.; Kronenberg, M. HVEM is a TNF Receptor with Multiple Regulatory Roles in the Mucosal Immune System. Immune Netw. 2014, 14, 67–72. [Google Scholar] [CrossRef]
- Morel, Y.; Truneh, A.; Sweet, R.W.; Olive, D.; Costello, R.T. The TNF superfamily members LIGHT and CD154 (CD40 ligand) costimulate induction of dendritic cell maturation and elicit specific CTL activity. J. Immunol. 2001, 167, 2479–2486. [Google Scholar] [CrossRef]
- Duhen, T.; Pasero, C.; Mallet, F.; Barbarat, B.; Olive, D.; Costello, R.T. LIGHT costimulates CD40 triggering and induces immunoglobulin secretion; a novel key partner in T cell-dependent B cell terminal differentiation. Eur. J. Immunol. 2004, 34, 3534–3541. [Google Scholar] [CrossRef] [PubMed]
- Pasero, C.; Speiser, D.E.; Derré, L.; Olive, D. The HVEM network: New directions in targeting novel costimulatory/co-inhibitory molecules for cancer therapy. Curr. Opin. Pharmacol. 2012, 12, 478–485. [Google Scholar] [CrossRef]
- Murphy, T.L.; Murphy, K.M. Slow down and survive: Enigmatic immunoregulation by BTLA and HVEM. Annu. Rev. Immunol. 2010, 28, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Freeman, G.J. The CD160, BTLA, LIGHT/HVEM pathway: A bidirectional switch regulating T-cell activation. Immunol. Rev. 2009, 229, 244–258. [Google Scholar] [CrossRef]
- Šedý, J.R.; Ramezani-Rad, P. HVEM network signaling in cancer. Adv. Cancer Res. 2019, 142, 145–186. [Google Scholar]
- Malissen, N.; Macagno, N.; Granjeaud, S.; Granier, C.; Moutardier, V.; Gaudy-Marqueste, C.; Habel, N.; Mandavit, M.; Guillot, B.; Pasero, C.; et al. HVEM has a broader expression than PD-L1 and constitutes a negative prognostic marker and potential treatment target for melanoma. OncoImmunology 2019, 8, e1665976. [Google Scholar] [CrossRef] [PubMed]
- Aldahlawi, A.; Basingab, F.; Alrahimi, J.; Zaher, K.; Pushparaj, P.N.; Hassan, M.A.; Al-Sakkaf, K. Herpesvirus entry mediator as a potential biomarker in breast cancer compared with conventional cytotoxic T-lymphocyte-associated antigen 4. Biomed. Rep. 2023, 19, 56. [Google Scholar] [CrossRef]
- M’hidi, H.; Thibult, M.-L.; Chetaille, B.; Rey, F.; Bouadallah, R.; Nicollas, R.; Olive, D.; Xerri, L. High expression of the inhibitory receptor BTLA in T-follicular helper cells and in B-cell small lymphocytic lymphoma/chronic lymphocytic leukemia. Am. J. Clin. Pathol. 2009, 132, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Guo, R.; Hsu, T.L.; Yu, G.L.; Ni, J.; Kwon, B.S.; Jiang, G.W.; Lu, J.; Tan, J.; Ugustus, M.; et al. LIGHT, a novel ligand for lymphotoxin beta receptor and TR2/HVEM induces apoptosis and suppresses in vivo tumor formation via gene transfer. J. Clin. Invest. 1998, 102, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Demerlé, C.; Gorvel, L.; Olive, D. BTLA-HVEM Couple in Health and Diseases: Insights for Immunotherapy in Lung Cancer. Front. Oncol. 2021, 11, 682007. [Google Scholar] [CrossRef]
- Hobo, W.; Norde, W.J.; Schaap, N.; Fredrix, H.; Maas, F.; Schellens, K.; Falkenburg, J.H.F.; Korman, A.J.; Olive, D.; van der Voort, R.; et al. B and T lymphocyte attenuator mediates inhibition of tumor-reactive CD8+ T cells in patients after allogeneic stem cell transplantation. J. Immunol. 2012, 189, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Pasero, C.; Barbarat, B.; Just-Landi, S.; Bernard, A.; Aurran-Schleinitz, T.; Rey, J.; Eldering, E.; Truneh, A.; Costello, R.T.; Olive, D. A role for HVEM, but not lymphotoxin-beta receptor, in LIGHT-induced tumor cell death and chemokine production. Eur. J. Immunol. 2009, 39, 2502–2514. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, S.; Sharma, P.; Jobanputra, Y.; Zhang, Y. Transformation of Chronic Myeloid Leukemia to Acute Biphenotypic Leukemia. J. Med Cases 2020, 11, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Gertner-Dardenne, J.; Fauriat, C.; Orlanducci, F.; Thibult, M.-L.; Pastor, S.; Fitzgibbon, J.; Bouabdallah, R.; Xerri, L.; Olive, D. The co-receptor BTLA negatively regulates human Vγ9Vδ2 T-cell proliferation: A potential way of immune escape for lymphoma cells. Blood 2013, 122, 922–931. [Google Scholar] [CrossRef]
- Park, J.-J.; Anand, S.; Zhao, Y.; Matsumura, Y.; Sakoda, Y.; Kuramasu, A.; Strome, S.E.; Chen, L.; Tamada, K. Expression of anti-HVEM single-chain antibody on tumor cells induces tumor-specific immunity with long-term memory. Cancer Immunol. Immunother. 2012, 61, 203–214. [Google Scholar] [CrossRef]
- Aubert, N.; Brunel, S.; Olive, D.; Marodon, G. Blockade of HVEM for Prostate Cancer Immunotherapy in Humanized Mice. Cancers 2021, 13, 3009. [Google Scholar] [CrossRef] [PubMed]
- Sordo-Bahamonde, C.; Lorenzo-Herrero, S.; Gonzalez-Rodriguez, A.P.; Payer, R.; González-García, E.; López-Soto, A.; Gonzalez, S. BTLA/HVEM Axis Induces NK Cell Immunosuppression and Poor Outcome in Chronic Lymphocytic Leukemia. Cancers 2021, 13, 1766. [Google Scholar] [CrossRef] [PubMed]
- Sideras, K.; Biermann, K.; Yap, K.; Mancham, S.; Boor, P.P.; Hansen, B.E.; Stoop, H.J.; Peppelenbosch, M.P.; van Eijck, C.H.; Sleijfer, S.; et al. Tumor cell expression of immune inhibitory molecules and tumor-infiltrating lymphocyte count predict cancer-specific survival in pancreatic and ampullary cancer. Int. J. Cancer 2017, 141, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Lu, M. Increased expression of TNFRSF14 indicates good prognosis and inhibits bladder cancer proliferation by promoting apoptosis. Mol. Med. Rep. 2018, 18, 3403–3410. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-H.; Kim, S.-H.; Lee, Y.; Lee, B.B.; Kwon, B.; Song, H.; Kwon, B.S.; Park, J.-E. Tumor necrosis factor receptor superfamily 14 is involved in atherogenesis by inducing proinflammatory cytokines and matrix metalloproteinases. Arter. Thromb. Vasc. Biol. 2001, 21, 2004–2010. [Google Scholar] [CrossRef] [PubMed]
- Haymaker, C.L.; Wu, R.C.; Ritthipichai, K.; Bernatchez, C.; Forget, M.-A.; Chen, J.Q.; Liu, H.; Wang, E.; Marincola, F.; Hwu, P.; et al. BTLA marks a less-differentiated tumor-infiltrating lymphocyte subset in melanoma with enhanced survival properties. Oncoimmunology 2015, 4, e1014246. [Google Scholar] [CrossRef]
- Malm, M.; Saghaleyni, R.; Lundqvist, M.; Giudici, M.; Chotteau, V.; Field, R.; Varley, P.G.; Hatton, D.; Grassi, L.; Svensson, T.; et al. Evolution from adherent to suspension: Systems biology of HEK293 cell line development. Sci. Rep. 2020, 10, 18996. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ye, Y.; Yu, H.; Lin, S.-H.; Tu, H.; Liang, D.; Chang, D.W.; Huang, M.; Wu, X. Immune checkpoint-related serum proteins and genetic variants predict outcomes of localized prostate cancer, a cohort study. Cancer Immunol. Immunother. 2021, 70, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Aldahlawi, A.; Alqadiri, A.; Alahdal, H.; Al-Sakkaf, K.; Alrahimi, J.; Basingab, F. Tumor necrosis factor alpha and lipopolysaccharides synergistic effects on T-cell immunoglobulin and mucin domain 3 regulation in dendritic cells. J. King Saud Univ. Sci. 2022, 34, 102213. [Google Scholar] [CrossRef]
- Basingab, F.S.; Ahmadi, M.; Morgan, D.J. IFNγ-Dependent Interactions between ICAM-1 and LFA-1 Counteract Prostaglandin E2-Mediated Inhibition of Antitumor CTL Responses. Cancer Immunol. Res. 2016, 4, 400–411. [Google Scholar] [CrossRef]
- Lewis, M.D.; de Leenheer, E.; Fishman, S.; Siew, L.K.; Gross, G.; Wong, F.S. A reproducible method for the expansion of mouse CD8+ T lymphocytes. J. Immunol. Methods 2015, 417, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Pierzchalski, A.; Mittag, A.; Bocsi, J.; Tarnok, A. An innovative cascade system for simultaneous separation of multiple cell types. PLoS ONE 2013, 8, e74745. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔC(T) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Choudhry, H.; Hassan, M.A.; Al-Malki, A.L.; Al-Sakkaf, K.A. Suppression of circulating AP001429.1 long non-coding RNA in obese patients with breast cancer. Oncol. Lett. 2021, 22, 508. [Google Scholar] [CrossRef] [PubMed]
- Rotte, A. Combination of CTLA-4 and PD-1 blockers for treatment of cancer. J. Exp. Clin. Cancer Res. 2019, 38, 255. [Google Scholar] [CrossRef] [PubMed]
- De Velasco, G.; Je, Y.; Bossé, D.; Awad, M.M.; Ott, P.A.; Moreira, R.B.; Schutz, F.; Bellmunt, J.; Sonpavde, G.P.; Hodi, F.S.; et al. Comprehensive Meta-analysis of Key Immune-Related Adverse Events from CTLA-4 and PD-1/PD-L1 Inhibitors in Cancer Patients. Cancer Immunol. Res. 2017, 5, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Hokuto, D.; Sho, M.; Yamato, I.; Yasuda, S.; Obara, S.; Nomi, T.; Nakajima, Y. Clinical impact of herpesvirus entry mediator expression in human hepatocellular carcinoma. Eur. J. Cancer 2015, 51, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Tsang, J.Y.S.; Chan, K.-W.; Ni, Y.-B.; Hlaing, T.; Hu, J.; Chan, S.-K.; Cheung, S.-Y.; Tse, G.M. Expression and Clinical Significance of Herpes Virus Entry Mediator (HVEM) in Breast Cancer. Ann. Surg. Oncol. 2017, 24, 4042–4050. [Google Scholar] [CrossRef]
- del Rio, M.-L.; de Juan, C.Y.-D.; Roncador, G.; Caleiras, E.; Álvarez-Esteban, R.; Pérez-Simón, J.A.; Rodriguez-Barbosa, J.-I. Genetic deletion of HVEM in a leukemia B cell line promotes a preferential increase of PD-1- stem cell-like T cells over PD-1+ T cells curbing tumor progression. Front. Immunol. 2023, 14, 1113858. [Google Scholar] [CrossRef]
- I Montgomery, R.; Warner, M.S.; Lum, B.J.; Spear, P.G. Herpes simplex virus-1 entry into cells mediated by a novel member of the TNF/NGF receptor family. Cell 1996, 87, 36–427. [Google Scholar] [CrossRef] [PubMed]
- Yegin, Z.; Duran, T.; Yildirim, I.H. Thymoquinone Down-regulates VEGFA and Up-regulates FLT1 Transcriptional Levels in Human Breast Cancer Cells. Int. J. Hum. Genet. 2020, 20, 19–24. [Google Scholar] [CrossRef]
- Mintz, M.A.; Felce, J.H.; Chou, M.Y.; Mayya, V.; Xu, Y.; Shui, J.W.; An, J.; Li, Z.; Marson, A.; Okada, T.; et al. The HVEM-BTLA Axis Restrains T Cell Help to Germinal Center B Cells and Functions as a Cell-Extrinsic Suppressor in Lymphomagenesis. Immunity 2019, 51, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Pal, D.; Blair, H.J.; Elder, A.; Dormon, K.; Rennie, K.J.; Coleman, D.J.L.; Weiland, J.; Rankin, K.S.; Filby, A.; Heidenreich, O.; et al. Long-term in vitro maintenance of clonal abundance and leukaemia-initiating potential in acute lymphoblastic leukaemia. Leukemia 2016, 30, 700–1691. [Google Scholar] [CrossRef] [PubMed]
- Valle-Reyes, S.; Dobrovinskaya, O.; Pelayo, R.; Schnoor, M. Acute Lymphoblastic Leukemia Cell Lines in Immunology Research. Trends Immunol. 2021, 42, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Demerlé, C.; Gorvel, L.; Mello, M.; Pastor, S.; Degos, C.; Zarubica, A.; Angelis, F.; Fiore, F.; Nunes, J.A.; Malissen, B.; et al. Anti-HVEM mAb therapy improves antitumoral immunity both in vitro and in vivo, in a novel transgenic mouse model expressing human HVEM and BTLA molecules challenged with HVEM expressing tumors. J. Immunother Cancer 2023, 11, e006348. [Google Scholar] [CrossRef] [PubMed]
- Ning, Z.; Liu, K.; Xiong, H. Roles of BTLA in Immunity and Immune Disorders. Front. Immunol. 2021, 12, 654960. [Google Scholar] [CrossRef] [PubMed]
- Compaan, D.M.; Gonzalez, L.C.; Tom, I.; Loyet, K.M.; Eaton, D.; Hymowitz, S.G. Attenuating lymphocyte activity: The crystal structure of the BTLA-HVEM complex. J. Biol. Chem. 2005, 280, 39553–39561. [Google Scholar] [CrossRef] [PubMed]
- Le Mercier, I.; Lines, J.L.; Noelle, R.J. Beyond CTLA-4 and PD-1, the Generation Z of Negative Checkpoint Regulators. Front. Immunol. 2015, 6, 159365. [Google Scholar] [CrossRef]
- del Rio, M.-L.; Kaye, J.; Rodriguez-Barbosa, J.-I. Detection of protein on BTLAlow cells and in vivo antibody-mediated down-modulation of BTLA on lymphoid and myeloid cells of C57BL/6 and BALB/c BTLA allelic variants. Immunobiology 2010, 215, 570–578. [Google Scholar] [CrossRef]
- Liu, J.; Li, J.; He, M.; Zhang, G.-L.; Zhao, Q. Distinct Changes of BTLA and HVEM Expressions in Circulating CD4+ and CD8+ T Cells in Hepatocellular Carcinoma Patients. J. Immunol. Res. 2018, 2018, 4561571. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Gamaleldin, M.A.; Imbaby, S.A.E. The role of tumor necrosis factor receptor superfamily member 4 (TNFRSF4) gene expression in diagnosis and prognosis of acute myeloid leukemia. Mol. Biol. Rep. 2021, 48, 6831–6843. [Google Scholar] [CrossRef] [PubMed]
- Lichtenegger, F.S.; Kondla, I.; Krempasky, M.; Weber, A.L.; Herold, T.; Krupka, C.; Spiekermann, K.; Schneider, S.; Büchner, T.; Berdel, W.E.; et al. RNA and protein expression of herpesvirus entry mediator (HVEM) is associated with molecular markers, immunity-related pathways and relapse-free survival of patients with AML. Cancer Immunol. Immunother. 2015, 64, 1505–1515. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Meng, H.; Tian, Y.; Wang, Y.; Song, T.; Zhang, T.; Wu, Q.; Cui, Y.; Li, H.; Zhang, W.; et al. High expression of HVEM is associated with improved prognosis in intrahepatic cholangiocarcinoma. Oncol. Lett. 2021, 21, 69. [Google Scholar] [CrossRef]
- Kang, S.H.; Hwang, H.J.; Yoo, J.W.; Kim, H.; Choi, E.S.; Hwang, S.-H.; Cho, Y.-U.; Jang, S.; Park, C.-J.; Im, H.J.; et al. Expression of Immune Checkpoint Receptors on T-Cells and Their Ligands on Leukemia Blasts in Childhood Acute Leukemia. Anticancer. Res. 2019, 39, 5531–5539. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | HVEM Ct Mean | GAPDH Ct Mean | ∆Ct | ∆∆Ct | Fold Change (FC) |
|---|---|---|---|---|---|
| MCF-7 | 32.486 | 21.885 | 10.602 | −0.854 | 1.8 |
| HepG2 | 33.60 | 23.223 | 10.230 | −1.225 | 2.3 |
| K562 | 25.268 | 22.368 | 2.900 | −8.556 | 376.3 * |
| Hek293 | 33.230 | 21.774 | 11.456 | 0 | 1 |
| Total | Non-Malignant | Malignant | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Parameters | Mean ± SEM | Median | IQR | Mean ± SEM | Median | IQR | Mean ± SEM | Median | IQR | |
| Number of participants, n (%) | 33 (100) | 10 (30) | 23 (70) | |||||||
| Gender | - | 6 Female (60%), 4 Male (40%) | 13 Female (57%), 10 Male (43%) | |||||||
| Age (years) | 17 ± 3 | 12 | 21.25 | 35 ± 3 | 38 | 12.5 | 8 ± 1.3 | 7 | 4 | |
| Initial WBC count (unit) | - | - | - | - | - | - | 74.13 ± 39 | 13.3 | 54.65 | |
| Disease status at study enrollment | ||||||||||
| Newly diagnosed | Remission | Relapse | Refractory | |||||||
| 3 (13%) | 8 (35%) | 10% (45%) | 5 (22%) | |||||||
| Male | Female | Male | Female | Male | Female | Male | Female | |||
| 2 | 1 | 3 | 5 | 4 | 6 | 2 | 3 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basingab, F.S.; Alzahrani, R.A.; Alrofaidi, A.A.; Barefah, A.S.; Hammad, R.M.; Alahdal, H.M.; Alrahimi, J.S.; Zaher, K.A.; Algiraigri, A.H.; El-Daly, M.M.; et al. Herpesvirus Entry Mediator as an Immune Checkpoint Target and a Potential Prognostic Biomarker in Myeloid and Lymphoid Leukemia. Biomolecules 2024, 14, 523. https://doi.org/10.3390/biom14050523
Basingab FS, Alzahrani RA, Alrofaidi AA, Barefah AS, Hammad RM, Alahdal HM, Alrahimi JS, Zaher KA, Algiraigri AH, El-Daly MM, et al. Herpesvirus Entry Mediator as an Immune Checkpoint Target and a Potential Prognostic Biomarker in Myeloid and Lymphoid Leukemia. Biomolecules. 2024; 14(5):523. https://doi.org/10.3390/biom14050523
Chicago/Turabian StyleBasingab, Fatemah S., Reem A. Alzahrani, Aisha A. Alrofaidi, Ahmed S. Barefah, Rawan M. Hammad, Hadil M. Alahdal, Jehan S. Alrahimi, Kawther A. Zaher, Ali H. Algiraigri, Mai M. El-Daly, and et al. 2024. "Herpesvirus Entry Mediator as an Immune Checkpoint Target and a Potential Prognostic Biomarker in Myeloid and Lymphoid Leukemia" Biomolecules 14, no. 5: 523. https://doi.org/10.3390/biom14050523
APA StyleBasingab, F. S., Alzahrani, R. A., Alrofaidi, A. A., Barefah, A. S., Hammad, R. M., Alahdal, H. M., Alrahimi, J. S., Zaher, K. A., Algiraigri, A. H., El-Daly, M. M., Alkarim, S. A., & Aldahlawi, A. M. (2024). Herpesvirus Entry Mediator as an Immune Checkpoint Target and a Potential Prognostic Biomarker in Myeloid and Lymphoid Leukemia. Biomolecules, 14(5), 523. https://doi.org/10.3390/biom14050523