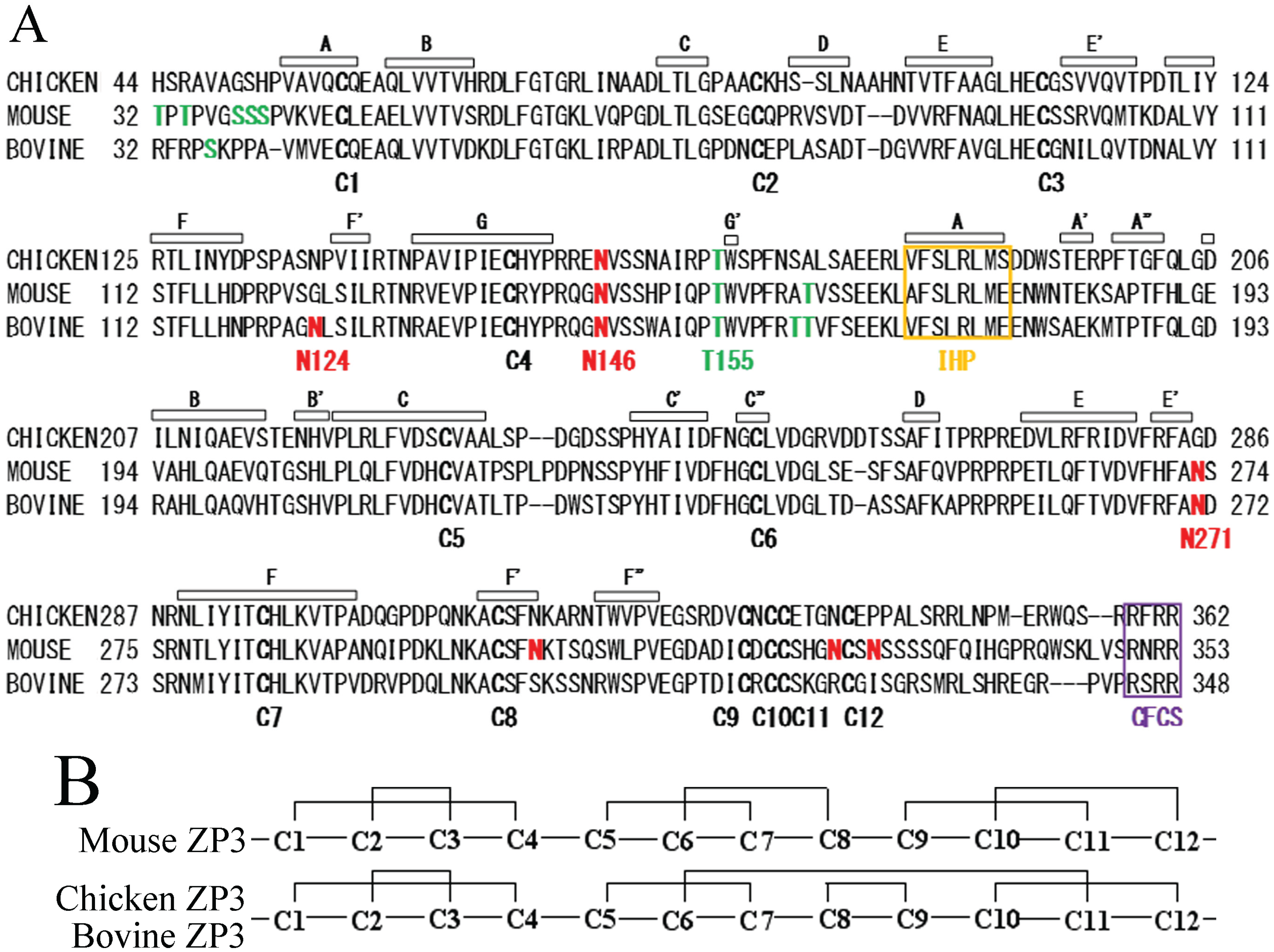
2.1. Identification of ZP4-Interaction Sites on ZP3
The disulfide bond pattern of bovine ZP3 has yet to be determined. However, sequence similarities between bovine ZP3 and porcine ZP3 suggest that the disulfide bond pattern of bovine ZP3 is the same as that of porcine ZP3 as well as that of chicken ZP3 (
Figure 1B) [
23]. The
N-terminal region Val-41 to Arg-143 of bovine ZP3 containing four Cys residues at 45, 77, 98, and 139 (translational initiation Met is numbered as (1) corresponds to ZP-N. As the interaction between IHP and EHP has been shown to inhibit premature assembly of ZP proteins [
19], IHP may be involved in the interaction of ZP3 with ZP4. The IHP is located in the region spanning Val-170 to Glu-177 (
Figure 1A). The region from IHP to the
C-terminus of bovine ZP3, containing eight Cys residues at 216, 238, 280, 299, 318, 320, 321, and 326 corresponds to ZP-C. Further fragmentation of ZP-N and ZP-C is not possible due to the disulfide bonds. The sequence similarities (57% identity) and the same disulfide bond pattern between bovine ZP3 and chicken ZP3 suggest that secondary and tertiary structures of bovine ZP3 are similar to those of chicken ZP3 (see
Figure 1A for β-strands). Based on X-ray crystallographic models of the chicken ZP3 precursor, residues 143 to 169 of bovine ZP3 are thought to function as the flexible hinge region, although the three-dimensional structure of this region has yet to be determined [
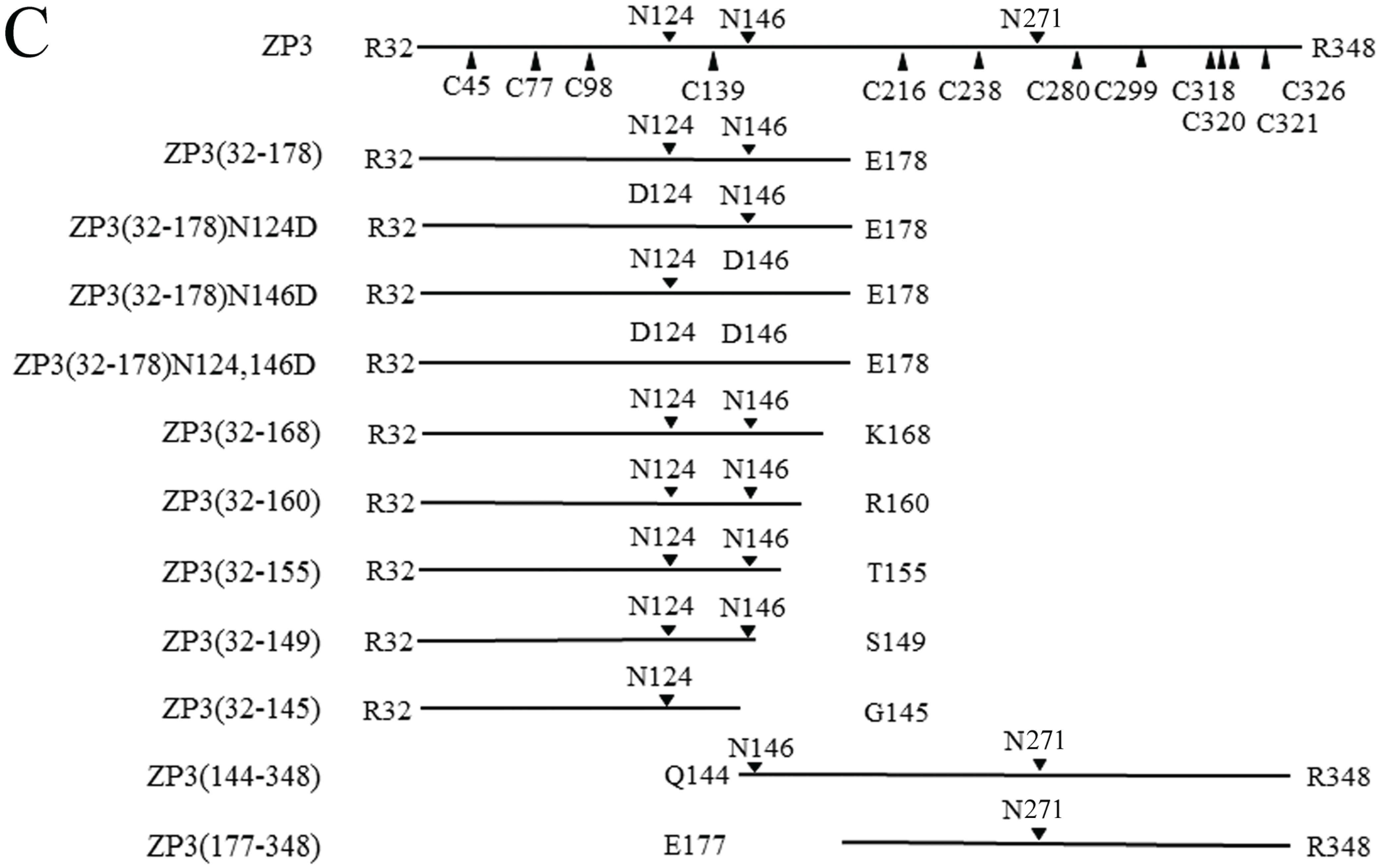
20]. Using these structural features as a starting point, fragments of the mature bovine ZP3 polypeptide spanning the
N-terminus of mature ZP3 to IHP (Arg-32 to Glu-178), the
N-terminus without the IHP (Arg-32 to Lys-168) and the
N-terminus truncated prior to the hinge region (Arg-32 to Gly-145), along with
C-terminal fragments Glu-177 to Arg-348 and Gln-144 to Arg-348 were expressed using a baculovirus-Sf9 cell system (
Figure 1C). Each ZP3 fragment was tagged with six histidine residues (His-tag) followed by S-tag on the
N-terminus of the protein and co-expressed with ZP4 containing an
N-terminal FLAG-tag and an S-tag by co-infecting Sf9 cells with the corresponding recombinant baculoviruses. ZP3 fragments were pulled down using TALON affinity resin specific for the His-tag, whereas co-precipitated ZP4 was detected with an antibody specific to FLAG-tag.
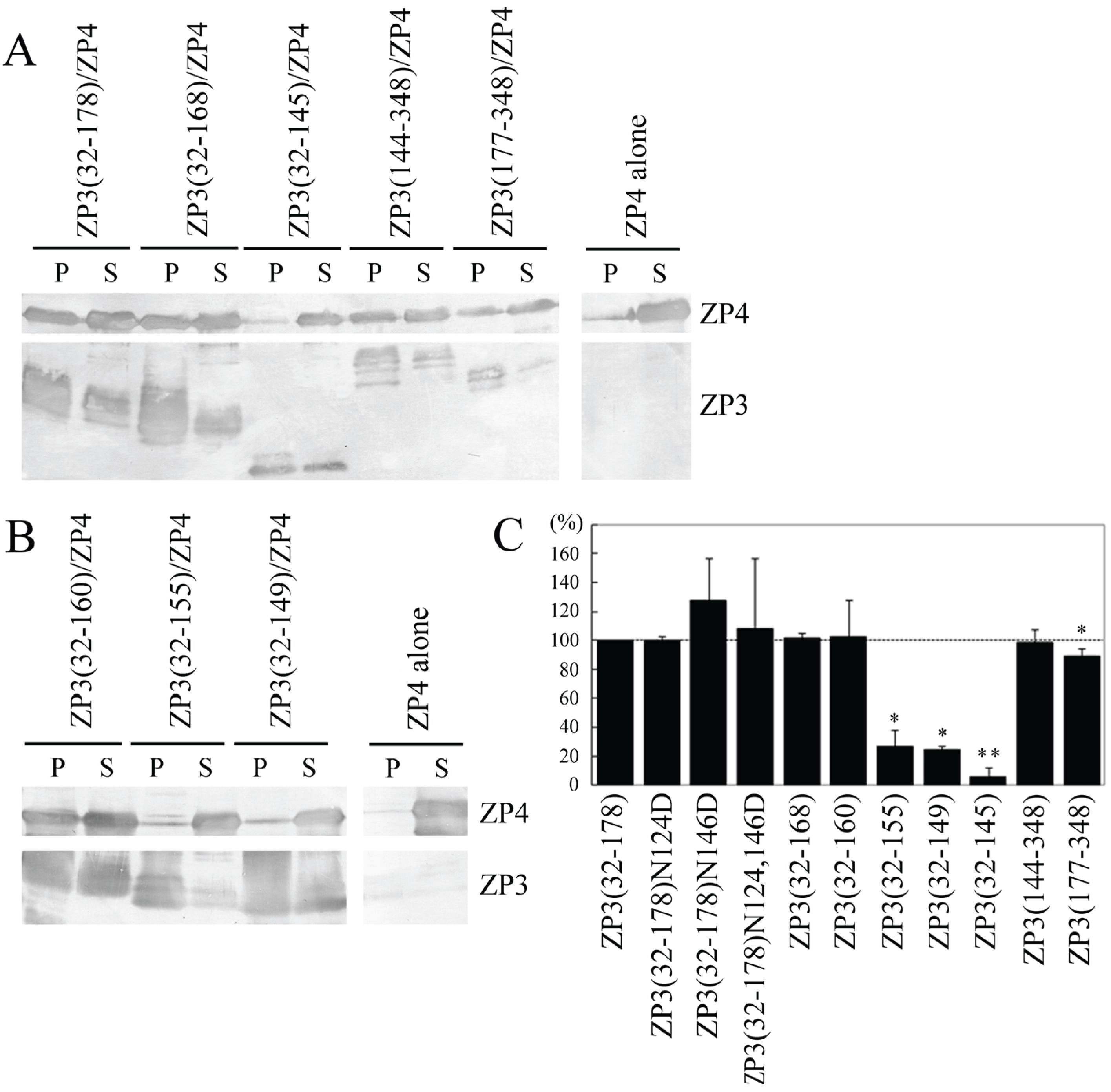
When ZP4 was expressed alone, only a small quantity of ZP4 was precipitated by TALON resin (
Figure 2A). This quantity was significantly increased when co-expressed with ZP3(32–178), ZP3(32–168), ZP3(144–348) or ZP3(177–348); however, when ZP4 was co-expressed with ZP3(32–145), the quantity of precipitated ZP4 was similar to that of ZP4 expressed alone (
Figure 2A). Relative quantities of precipitated ZP4 were calculated based on the quantities recovered when ZP4 was expressed alone and when it was co-expressed with ZP3(32–178), respectively, as determined using NIH Image J software. The percentages of co-precipitated ZP4 were not significantly different between ZP3(32–178) and ZP3(32–168), suggesting that the IHP of ZP3 is not essential for the interaction of ZP3 with ZP4. In contrast, the percentage of co-precipitated ZP4 was dramatically reduced to near basal levels when co-expressed with ZP3(32–145), suggesting that the region from 146 to 168 is necessary for the interaction of ZP3(32–178) with ZP4. The quantity of ZP4 recovered following co-expression with ZP3(177–348) was significantly lower than that of ZP4 co-expressed with ZP3(144–348); however, the difference in percentages was small, suggesting that one of the ZP4 binding sites is located in the region 177–348, making region 144–176 dispensable for the ZP4 binding of ZP3(144–348). Further efforts to identify the region within the 177–348 fragment responsible for ZP4 binding were not performed, as we instead chose to focus on the ZP4 interaction site in the region 146 to 168 of ZP3.
Figure 2.
Co-precipitation of ZP4 with ZP3 fragments. (A,B) Detection of N-terminally FLAG- and S-tagged ZP4 co-precipitated with N-terminally His- and S-tagged ZP3 fragments by Western blot. ZP4 was expressed alone (ZP4 alone) or in combination with ZP3 fragments (indicated above each panel). The recombinant ZP3 fragments were pulled down using TALON resin. FLAG-tagged ZP4 in the pellet (P) and in the supernatant (S) was detected with an antibody specific to FLAG-tag (upper panels, ZP4); His-tagged ZP3 fragments in the P and the S were detected with an antibody specific to His-tag (lower panels, ZP3); (C) Percentage of ZP4 co-precipitated with each ZP3 fragment among the total ZP4 recovered in the pellet and supernatant. Percentages were quantified as described in the Experimental Section. The percentage of ZP4 co-precipitated with ZP3(32–178) is represented as 100%. Experiments were performed at least three times. Data are shown as mean ± S.D. Statistically significant differences, relative to 100% expression, are indicated as p > 0.05 (*) and p > 0.01 (**).
Figure 2.
Co-precipitation of ZP4 with ZP3 fragments. (A,B) Detection of N-terminally FLAG- and S-tagged ZP4 co-precipitated with N-terminally His- and S-tagged ZP3 fragments by Western blot. ZP4 was expressed alone (ZP4 alone) or in combination with ZP3 fragments (indicated above each panel). The recombinant ZP3 fragments were pulled down using TALON resin. FLAG-tagged ZP4 in the pellet (P) and in the supernatant (S) was detected with an antibody specific to FLAG-tag (upper panels, ZP4); His-tagged ZP3 fragments in the P and the S were detected with an antibody specific to His-tag (lower panels, ZP3); (C) Percentage of ZP4 co-precipitated with each ZP3 fragment among the total ZP4 recovered in the pellet and supernatant. Percentages were quantified as described in the Experimental Section. The percentage of ZP4 co-precipitated with ZP3(32–178) is represented as 100%. Experiments were performed at least three times. Data are shown as mean ± S.D. Statistically significant differences, relative to 100% expression, are indicated as p > 0.05 (*) and p > 0.01 (**).
![Biomolecules 05 03339 g002]()
Evidence of a ZP4 binding site in the hinge region of ZP3 (residues 146–168) was further investigated by co-precipitation of ZP4 with each of the following ZP3 fragments: ZP3(32–160), ZP3(32–155), and ZP3(32–149) (
Figure 1C). The co-precipitation of ZP4 with ZP3(32–160) was similar to that of ZP3(32–168) (
Figure 2B). Further deletion from 156 to 160 dramatically reduced the co-precipitation of ZP4; additional deletions between residues 150 and 155 failed to restore the level of co-precipitated ZP4. These results suggest that the region spanning 161 to 178 is not necessary for the interaction of ZP3(32–178) with ZP4, whereas residues 156 to 160 are critically important for the binding of ZP4 to ZP3(32–178). Further deletion from 146 to 149 reduced the quantity of precipitated ZP4 by a small but significant extent, suggesting that the region from 146 to 149 is also involved in the ZP4 binding of ZP3.
Among residues 156–160, Arg-160 is the least conserved, with Ser, Trp, and His residues observed at this site in other mammals. Val-157 is almost completely conserved, although Leu is also found in some mammals. Trp-156, Pro-158, and Phe-159 are completely conserved across all mammals described to date; this strong conservation suggests that these residues are likely important for the binding of ZP4 to ZP3(32–178).
Our results indicating an interaction between the C-terminal half of ZP3 and ZP4 are consistent with previous reports describing avian homologs of ZP3. In addition to this region, these data suggest that there is also an additional ZP4-interaction site in the flexible hinge region of bovine ZP3.
2.2. Involvement of the Hinge Region of ZP3 in the Sperm-Binding Activity of the ZP3/ZP4 Complex
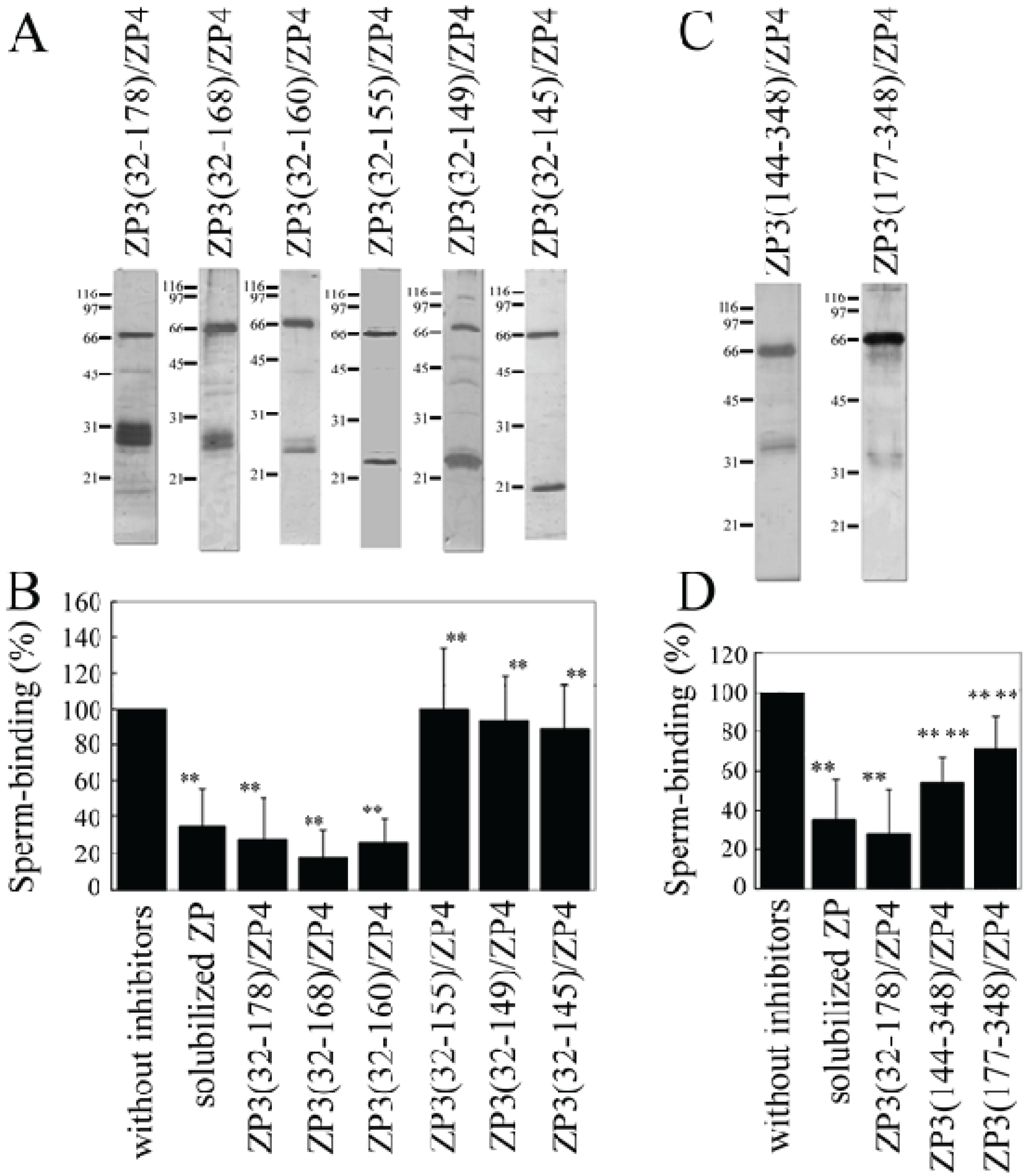
A mixture of each of the ZP3 fragments and ZP4 was partially purified using TALON resin (
Figure 3A,C) and used to test the inhibitory activity of each of the mixtures for the binding of bovine sperm to plastic wells coated with solubilized bovine ZP. The number of bovine sperm binding to ZP-coated wells was reduced to ~35% in the presence of solubilized bovine ZP. Neither the ZP3 fragment nor the ZP4 alone significantly inhibited sperm-ZP binding. In contrast, the ZP3(32–178)/ZP4 mixture significantly inhibited binding (
Figure 3B). We next tried to identify a region on ZP3(32–178) involved in the formation of a sperm-binding active complex with ZP4. The co-expressed mixtures of ZP3(32–168)/ZP4 and ZP3(32–160)/ZP4 showed inhibitory activities for sperm-ZP binding that were similar to those of ZP3(32–178)/ZP4, whereas ZP3(32–155)/ZP4, ZP3(32–149)/ZP4, and ZP3(32–145)/ZP4 failed to significantly inhibit sperm-ZP binding. Thus, the inhibitory activity seen here is consistent with that of the co-precipitation result, confirming our previous result showing that the formation of the ZP3/ZP4 complex is necessary for its sperm-binding activity [
12] and further suggesting that residues 161 to 178 are not necessary for the formation of the sperm-binding active ZP3/ZP4 complex.
The combinations of ZP3(144–348)/ZP4 and ZP3(177–348)/ZP4 significantly inhibited sperm-ZP binding, although the inhibitory activity of ZP3(177–348)/ZP4 was significantly lower than that of ZP3(144–348)/ZP4 (
Figure 3D). These results suggest that the
C-terminal region of ZP3 can form sperm-binding active complexes with ZP4 and that the region spanning residues 144 to 176 is involved in the formation of complexes possessing higher sperm-binding activity. Similarly, the inhibitory activity of ZP3(144–348)/ZP4 for sperm-ZP binding was significantly lower than that of the ZP3(32–178)/ZP4 mixture, suggesting that residues 144 to 176 contribute more to the sperm-binding activity of the ZP3(32–178)/ZP4 mixture than to the activity of ZP3(144–348)/ZP4.
2.3. Involvement of N-Linked Glycans on ZP3(32–178) in the Formation of Sperm-Binding Complexes
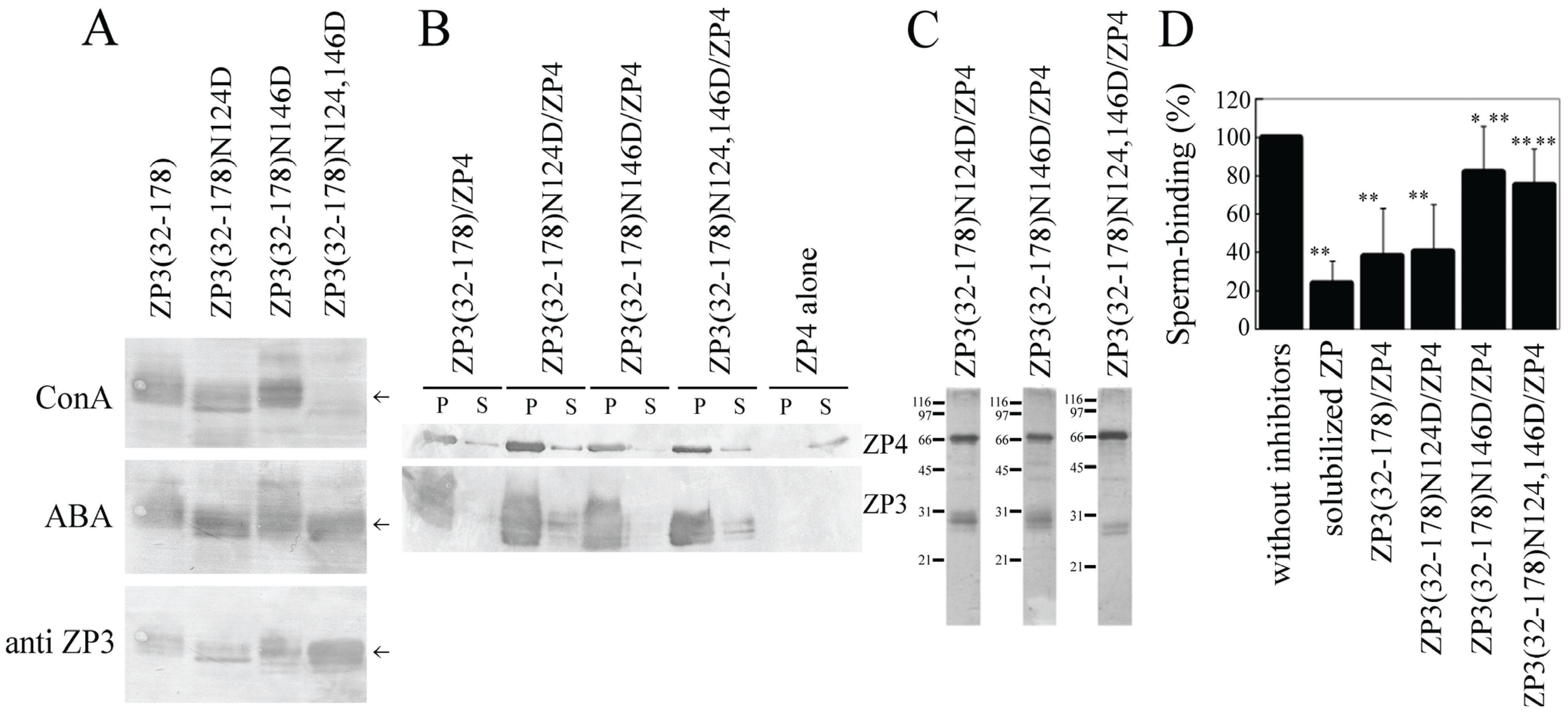
ZP3(32–178) has two
N-glycosylation sites at Asn-124 and Asn-146. Here, we mutated each of these sites, either alone or in combination, to Asp. Concanavalin A recognized both ZP3(32–178) and ZP3(32–178) with Asn-124 mutated to Asp (ZP3(32–178)N124D), as well as ZP3(32–178) with Asn-146 mutated to Asp (ZP3(32–178)N146D) (
Figure 4A), indicating that both Asn residues are
N-glycosylated. Concanavalin A did not recognize ZP3(32–178) with both Asn-124 and Asn-146 mutated to Asp residues (ZP3(32–178)N124,146D) (
Figure 4A), confirming the mutation of the two
N-glycosylated Asn residues. Mutation of either Asn-124 or Asn-146 to Asp failed to reduce the quantity of ZP4 co-precipitated with ZP3(32–178) (
Figure 4B and
Figure 2C), indicating that
N-glycosylation is not necessary for complex formation between ZP3(32–178) and ZP4. This result further indicates that these mutations did not destroy the steric structure of ZP3(32–178), since ZP3(32–178) retained the activity for the formation of a complex with ZP4 after the
N-glycosylation sites were mutated.
Figure 3.
Inhibitory activity of a mixture of ZP3 fragment and ZP4 for the binding of bovine sperm to the zona pellucida (ZP). (A,C) SDS-polyacrylamide gel electrophoresis of a mixture of each ZP3 fragment and ZP4. N-terminally His-tagged ZP4 and an N-terminally His-tagged ZP3 fragment (indicated above each panel) were co-expressed in Sf9 cells, and the mixture was partially purified using TALON resin specific to His-tag. Gels were then silver-stained, with molecular mass standards (kDa) indicated on the left side of each panel. Bands at ~66 kDa correspond to ZP4, and the bands at 21 to 31 kDa in (A) and the bands at ~35 kDa in (C) correspond to ZP3 fragments; (B,D) Inhibitory activity of the mixture of ZP3 fragment and ZP4 for sperm-ZP binding. The number of sperm binding to ZP-coated wells in the absence of inhibitors was designated as 100%. Assays were repeated at least three times. Data are presented as means ± S.D. with statistical significance relative to “without inhibitors” and “ZP3(32–178)/ZP4” (asterisks on the left side and the right side of S.D. bars, respectively) indicated as p > 0.05 (*) and p > 0.01 (**).
Figure 3.
Inhibitory activity of a mixture of ZP3 fragment and ZP4 for the binding of bovine sperm to the zona pellucida (ZP). (A,C) SDS-polyacrylamide gel electrophoresis of a mixture of each ZP3 fragment and ZP4. N-terminally His-tagged ZP4 and an N-terminally His-tagged ZP3 fragment (indicated above each panel) were co-expressed in Sf9 cells, and the mixture was partially purified using TALON resin specific to His-tag. Gels were then silver-stained, with molecular mass standards (kDa) indicated on the left side of each panel. Bands at ~66 kDa correspond to ZP4, and the bands at 21 to 31 kDa in (A) and the bands at ~35 kDa in (C) correspond to ZP3 fragments; (B,D) Inhibitory activity of the mixture of ZP3 fragment and ZP4 for sperm-ZP binding. The number of sperm binding to ZP-coated wells in the absence of inhibitors was designated as 100%. Assays were repeated at least three times. Data are presented as means ± S.D. with statistical significance relative to “without inhibitors” and “ZP3(32–178)/ZP4” (asterisks on the left side and the right side of S.D. bars, respectively) indicated as p > 0.05 (*) and p > 0.01 (**).
![Biomolecules 05 03339 g003]()
Figure 4.
Effect of
N-glycosylation site mutation of the ZP3 fragment on active sperm-binding complex formation between ZP3 fragment and ZP4. (
A) Lectin blots of
N-glycosylation site mutants of the ZP3(32–178) fragment. ZP3(32–178) fragment with Asn-124 mutated to Asp (ZP3(32–178)N124D), Asn-146 mutated to Asp (ZP3(32–178)N146D), and both Asn residues mutated to Asp (ZP3(32–178)N124,146D) as well as ZP3(32–178) were subjected to lectin blots. The same membrane was probed three times with Concanavalin A (ConA),
Agaricus bisporus agglutinin (ABA), and, finally, anti-porcine ZP3 antiserum (anti ZP3), as described in the Experimental Section. ZP3 fragments are indicated by arrows;
(B) Effect of
N-glycosylation site mutation on ZP4 co-precipitation.
N-terminally FLAG- and S-tagged ZP4 were expressed alone (ZP4 alone) or in combination with
N-terminally His- and S-tagged ZP3 fragments (indicated above each panel). The recombinant ZP3 fragments were pulled down by TALON resin specific to His-tag. FLAG-tagged ZP4 in the pellets (P) and in supernatants (S) were detected with an antibody specific to FLAG-tag (upper panel, ZP4); His-tagged ZP3 fragments in the P and the S were detected with an antibody specific to His-tag (lower panel, ZP3). Percentages of co-precipitated ZP4 are shown in
Figure 2C; (
C) SDS-polyacrylamide gel electrophoresis of mixtures of
N-glycosylation site mutants of ZP3(32–178) fragment and ZP4. Gels were then silver-stained, with molecular mass standards (kDa) indicated on the left side of each panel. Bands at ~66kDa correspond to ZP4; bands at 26 to 31kDa correspond to ZP3 fragments; (
D) Inhibitory activity of the mixture of ZP3 fragment and ZP4 for sperm-ZP binding. The number of sperm binding to the ZP-coated well in the absence of inhibitors was designated as 100%. Assays were repeated at least three times. Data are presented as means ± S.D. with statistical significance relative to “without inhibitors” and “ZP3(32–178)/ZP4” (asterisks on the left side and the right side of S.D. bars, respectively) indicated as
p > 0.05 (*) and
p > 0.01 (**).
Figure 4.
Effect of
N-glycosylation site mutation of the ZP3 fragment on active sperm-binding complex formation between ZP3 fragment and ZP4. (
A) Lectin blots of
N-glycosylation site mutants of the ZP3(32–178) fragment. ZP3(32–178) fragment with Asn-124 mutated to Asp (ZP3(32–178)N124D), Asn-146 mutated to Asp (ZP3(32–178)N146D), and both Asn residues mutated to Asp (ZP3(32–178)N124,146D) as well as ZP3(32–178) were subjected to lectin blots. The same membrane was probed three times with Concanavalin A (ConA),
Agaricus bisporus agglutinin (ABA), and, finally, anti-porcine ZP3 antiserum (anti ZP3), as described in the Experimental Section. ZP3 fragments are indicated by arrows;
(B) Effect of
N-glycosylation site mutation on ZP4 co-precipitation.
N-terminally FLAG- and S-tagged ZP4 were expressed alone (ZP4 alone) or in combination with
N-terminally His- and S-tagged ZP3 fragments (indicated above each panel). The recombinant ZP3 fragments were pulled down by TALON resin specific to His-tag. FLAG-tagged ZP4 in the pellets (P) and in supernatants (S) were detected with an antibody specific to FLAG-tag (upper panel, ZP4); His-tagged ZP3 fragments in the P and the S were detected with an antibody specific to His-tag (lower panel, ZP3). Percentages of co-precipitated ZP4 are shown in
Figure 2C; (
C) SDS-polyacrylamide gel electrophoresis of mixtures of
N-glycosylation site mutants of ZP3(32–178) fragment and ZP4. Gels were then silver-stained, with molecular mass standards (kDa) indicated on the left side of each panel. Bands at ~66kDa correspond to ZP4; bands at 26 to 31kDa correspond to ZP3 fragments; (
D) Inhibitory activity of the mixture of ZP3 fragment and ZP4 for sperm-ZP binding. The number of sperm binding to the ZP-coated well in the absence of inhibitors was designated as 100%. Assays were repeated at least three times. Data are presented as means ± S.D. with statistical significance relative to “without inhibitors” and “ZP3(32–178)/ZP4” (asterisks on the left side and the right side of S.D. bars, respectively) indicated as
p > 0.05 (*) and
p > 0.01 (**).
![Biomolecules 05 03339 g004]()
Co-expressed mixtures of ZP3(32–178)N124D and ZP4 inhibited sperm-ZP binding at levels similar to ZP3(32–178)/ZP4, whereas the mixture of ZP3(32–178)N146D and ZP4 exhibited significantly lower levels of inhibitory activity than ZP3(32–178)/ZP4 (
Figure 4C,D). Mutation of both Asn residues to Asp also significantly reduced the inhibitory activity of the ZP3(32–178)/ZP4 mixture (
Figure 4D). Thus, the mutation of Asn-146 to Asp did not reduce the co-precipitation of ZP4, but it did reduce the sperm-binding activity of the complex, suggesting that
N-glycan linked to Asn-146 is involved in the sperm-binding activity of ZP3(32–178)/ZP4.
When chicken ZP3 was analyzed by X-ray crystallography, the single
N-glycosylation site corresponding to Asn-146 of bovine ZP3 was mutated to Gln; however, the recombinant chicken ZP3 without
N-glycans still retained significant sperm-binding activity [
20]. On the other hand, mutation of a single
O-glycosylation site corresponding to Thr-155 in bovine ZP3 to Ala reduced the sperm-binding activity. As bovine ZP3 does not, by itself, inhibit sperm-ZP binding [
12], the mechanisms regulating the binding of sperm to ZP might differ in bovines and chickens.
In combination with previous reports detailing the structure of chicken ZP3, the data presented herein suggest that glycans linked to the flexible hinge region are involved in sperm binding in vitro. Further studies will be necessary to determine whether the N-glycan at Asn-146 of bovine ZP3 binds to sperm directly or if the absence of an N-glycan at Asn-146 has an allosteric effect on the structure of the ZP3(32–178)/ZP4 complex.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}