
Differential Degradation and Detoxification of an Aromatic Pollutant by Two Different Peroxidases



Abstract
:1. Introduction
2. Results and Discussion
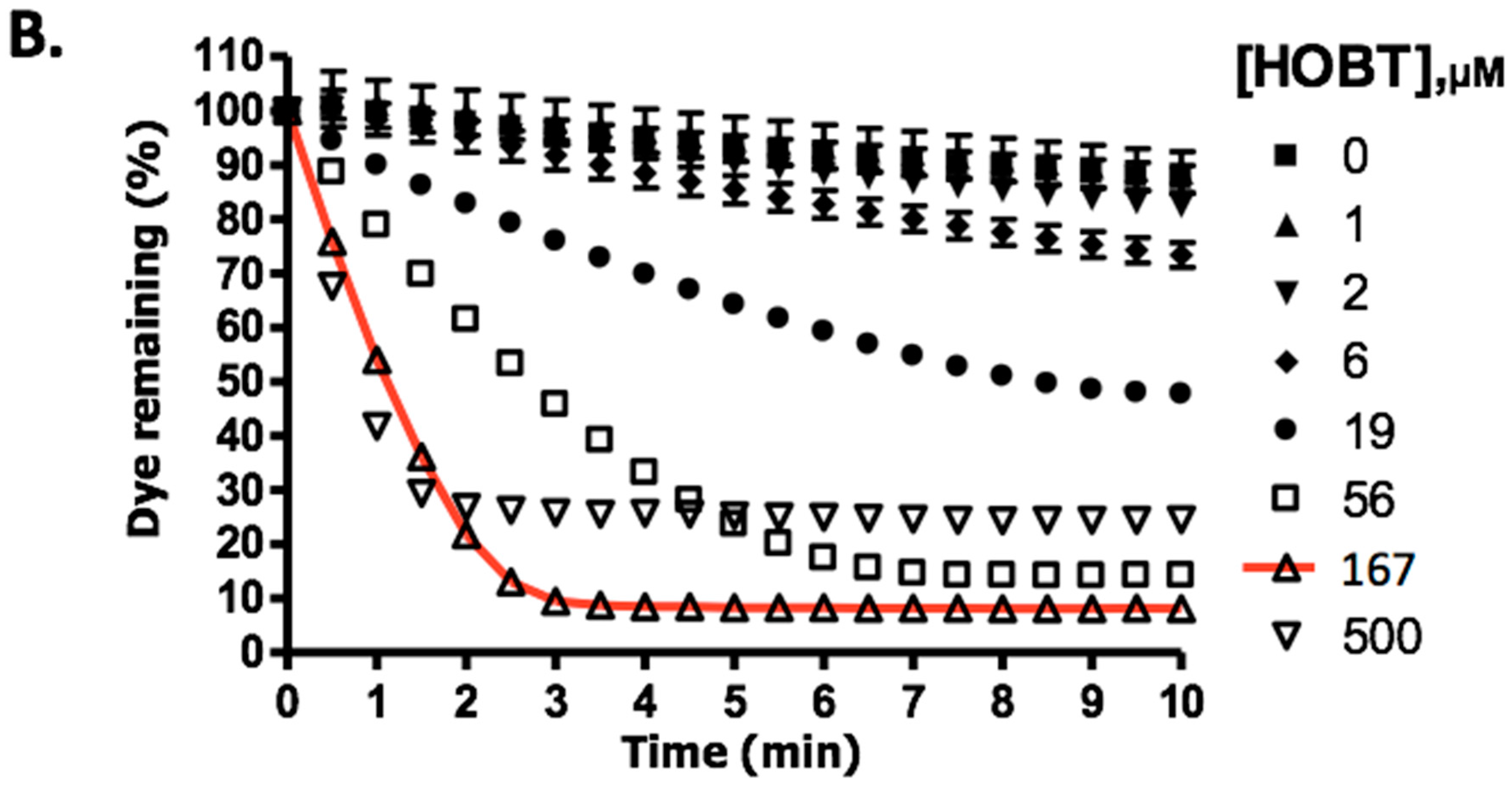
2.1. The Requirement for Redox Mediator
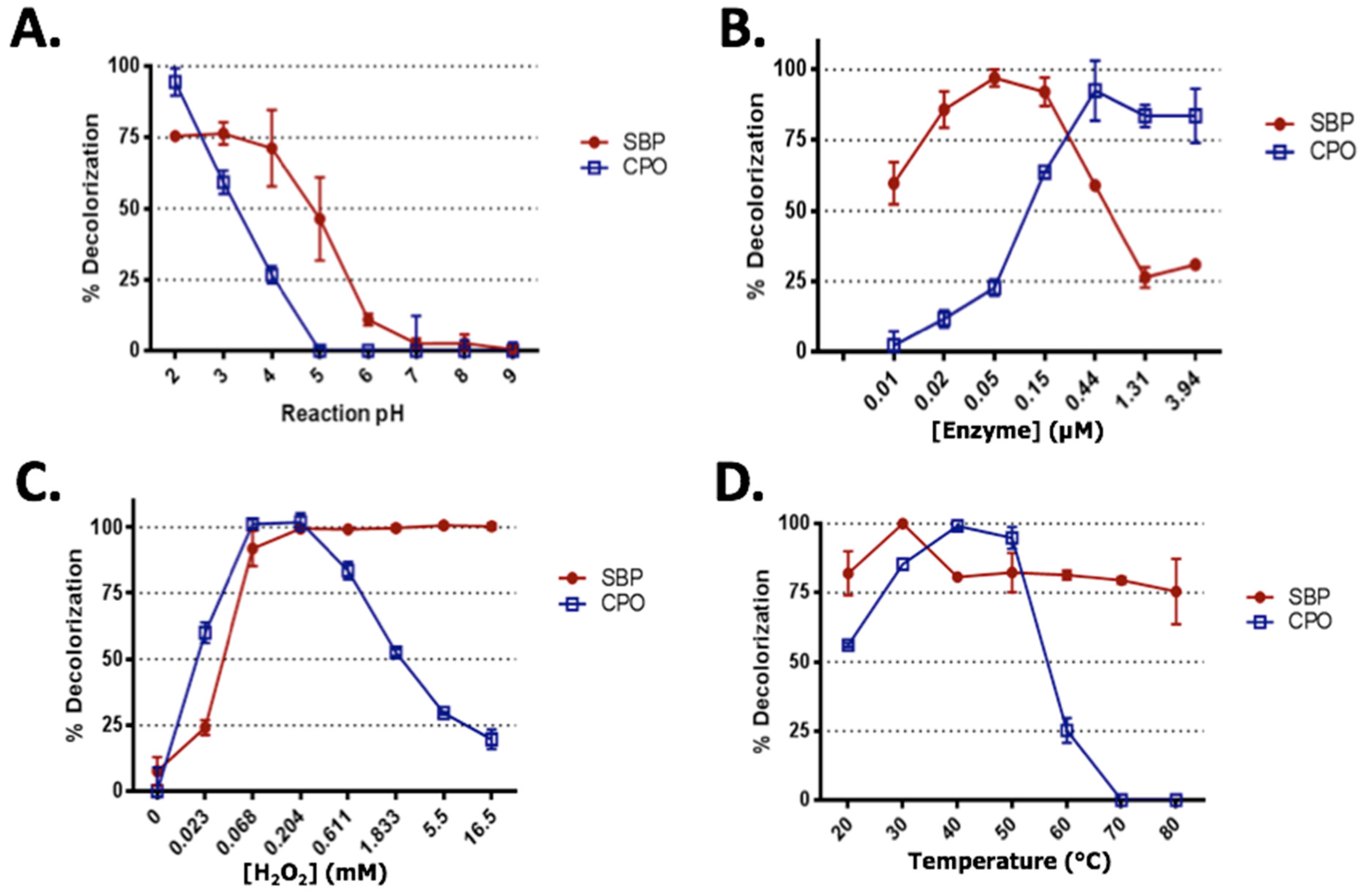
2.2. Effect of Reaction pH
2.3. Enzyme Concentrations
2.4. Hydrogen Peroxide Optimization
2.5. Effect of Temperature on SRB Dye Degradation
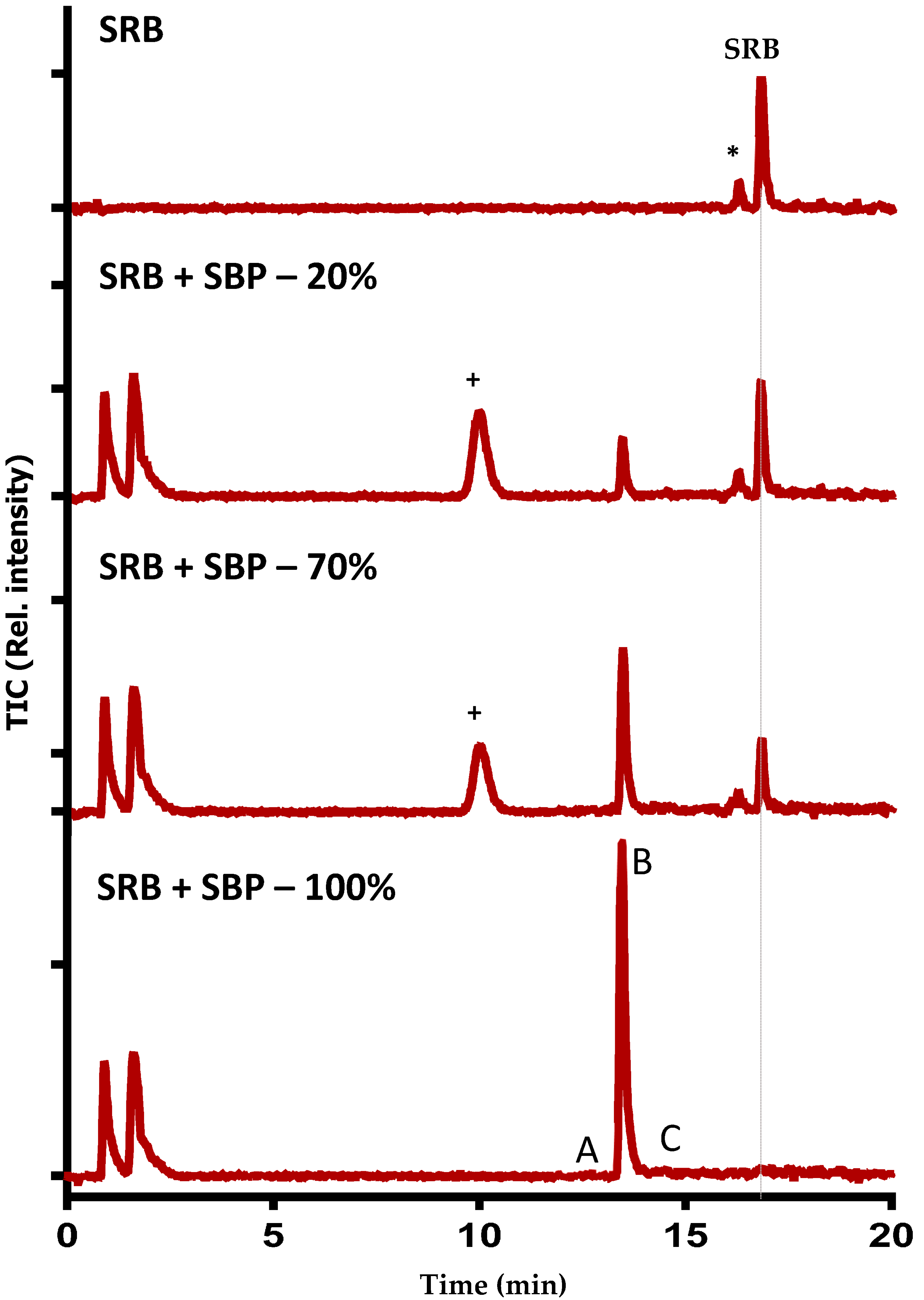
2.6. LC–MS Analyses
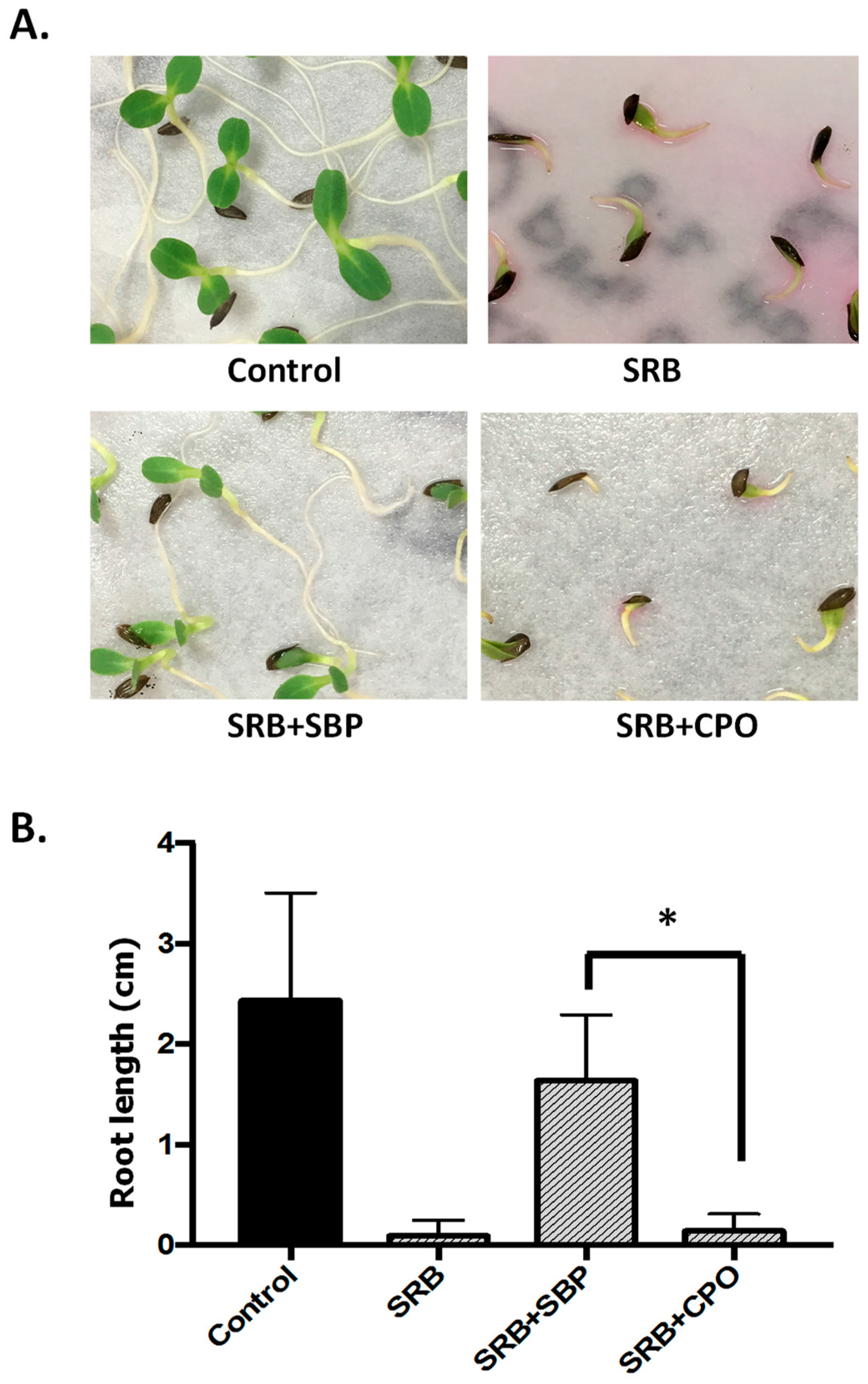
2.7. Toxicity Tests
2.8. The Effect of Halides on Enzymatic Degradation
3. Materials and Methods
3.1. Enzymes and Chemicals
3.2. Dye Degradation and Optimization Studies
3.3. HPLC and LC–MS Analyses
3.4. Phytotoxicity Assay
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, D.; Gersberg, R.M.; Ng, W.J.; Tan, S.K. Removal of pharmaceuticals and personal care products in aquatic plant-based systems: A review. Environ. Pollut. 2014, 184, 620–639. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Santos, J.M.; Diniz, M.S.; Ferreira, A.M.; Costa, M.H.; Costa, P.M. Effects of carcinogenic versus non-carcinogenic AHR-active PAHs and their mixtures: Lessons from ecological relevance. Environ. Res. 2015, 138, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.D.; Plewa, M.J.; Wagner, E.D.; Schoeny, R.; DeMarini, D.M. Occurrence, genotoxicity, and carcinogenicity of regulated and emerging disinfection by-products in drinking water: A review and roadmap for research. Mutat. Res./Rev. Mutat. Res. 2007, 636, 178–242. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.Z.; Ahmad, S.; Ahmad, M. Mutagenicity and genotoxicity of tannery effluents used for irrigation at Kanpur, India. Ecotoxicol. Environm. Saf. 2010, 73, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- De Lima, R.O.A.; Bazo, A.P.; Salvadori, D.M.F.; Rech, C.M.; de Palma Oliveira, D.; de Aragão Umbuzeiro, G. Mutagenic and carcinogenic potential of a textile azo dye processing plant effluent that impacts a drinking water source. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2007, 626, 53–60. [Google Scholar] [CrossRef] [PubMed]
- De Roos, A.J.; Ray, R.M.; Gao, D.L.; Wernli, K.J.; Fitzgibbons, E.D.; Ziding, F.; Astrakianakis, G.; Thomas, D.B.; Checkoway, H. Colorectal cancer incidence among female textile workers in Shanghai, China: A case-cohort analysis of occupational exposures. Cancer Causes Control 2005, 16, 1177–1188. [Google Scholar] [CrossRef] [PubMed]
- Amini, M.; Arami, M.; Mahmoodi, N.M.; Akbari, A. Dye removal from colored textile wastewater using acrylic grafted nanomembrane. Desalination 2011, 267, 107–113. [Google Scholar] [CrossRef]
- Capar, G.; Yilmaz, L.; Yetis, U. Reclamation of acid dye bath wastewater: Effect of pH on nanofiltration performance. J. Membr. Sci. 2006, 281, 560–569. [Google Scholar] [CrossRef]
- Lin, D.; Zhao, Q.; Hu, L.; Xing, B. Synthesis and characterization of cubic mesoporous bridged polysilsesquioxane for removing organic pollutants from water. Chemosphere 2014, 103, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Ali, I. New generation adsorbents for water treatment. Chem. Rev. 2012, 112, 5073–5091. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.B.; Vakili, M.; Horri, B.A.; Poh, P.E.; Abdullah, A.Z.; Salamatinia, B. Adsorption of dyes by nanomaterials: Recent developments and adsorption mechanisms. Sep. Purif. Technol. 2015, 150, 229–242. [Google Scholar] [CrossRef]
- Wang, J.; Chen, B. Adsorption and coadsorption of organic pollutants and a heavy metal by graphene oxide and reduced graphene materials. Chem. Eng. J. 2015, 281, 379–388. [Google Scholar] [CrossRef]
- Yagub, M.T.; Sen, T.K.; Afroze, S.; Ang, H.M. Dye and its removal from aqueous solution by adsorption: A review. Adv. Colloid Interface Sci. 2014, 209, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Chaalal, O.; Zekri, A.Y.; Soliman, A.M. A novel technique for the removal of strontium from water using thermophilic bacteria in a membrane reactor. J. Ind. Eng. Chem. 2015, 21, 822–827. [Google Scholar] [CrossRef]
- García-Delgado, C.; Yunta, F.; Eymar, E. Bioremediation of multi-polluted soil by spent mushroom (Agaricus bisporus) substrate: Polycyclic aromatic hydrocarbons degradation and Pb availability. J. Hazard. Mater. 2015, 300, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Helbling, D.E. Bioremediation of pesticide-contaminated water resources: The challenge of low concentrations. Curr. Opin. Biotechnol. 2015, 33, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Guo, S.; Jiao, K.; Hou, J.; Xie, H.; Xu, H. Bioremediation of soils co-contaminated with heavy metals and 2,4,5-trichlorophenol by fruiting body of Clitocybe maxima. J. Hazard. Mater. 2015, 294, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Paniagua-Michel, J. Chapter 31—Bioremediation with microalgae: Toward sustainable production of biofuels. In Handbook of Marine Microalgae; Kim, S.-K., Ed.; Academic Press: Boston, MA, USA, 2015; pp. 471–481. [Google Scholar]
- Ahemad, M. Enhancing phytoremediation of chromium-stressed soils through plant-growth-promoting bacteria. J. Genet. Eng. Biotechnol. 2015, 13, 51–58. [Google Scholar] [CrossRef]
- Li, W.-W.; Yu, H.-Q. From wastewater to bioenergy and biochemicals via two-stage bioconversion processes: A future paradigm. Biotechnol. Adv. 2011, 29, 972–982. [Google Scholar] [CrossRef] [PubMed]
- Malik, B.; Pirzadah, T.B.; Tahir, I.; Dar, T.u.H.; Rehman, R.U. In Soil Remediation and Plants; Hakeem, M., Sabir, M., Ozturk, M., Mermut, A., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 131–146. [Google Scholar]
- Ullah, A.; Heng, S.; Munis, M.F.H.; Fahad, S.; Yang, X. Phytoremediation of heavy metals assisted by plant growth promoting (PGP) bacteria: A review. Environ. Exp. Bot. 2015, 117, 28–40. [Google Scholar] [CrossRef]
- Bilal, M.; Asgher, M.; Iqbal, H.M.; Hu, H.; Zhang, X. Bio-based degradation of emerging endocrine-disrupting and dye-based pollutants using cross-linked enzyme aggregates. Environ. Sci. Pollut. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Coconi-Linares, N.; Ortiz-Vázquez, E.; Fernández, F.; Loske, A.M.; Gómez-Lim, M.A. Recombinant expression of four oxidoreductases in Phanerochaete chrysosporium improves degradation of phenolic and non-phenolic substrates. J. Biotechnol. 2015, 209, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Husain, Q. Peroxidase mediated decolorization and remediation of wastewater containing industrial dyes: A review. Rev. Environ. Sci. Biotechnol. 2010, 9, 117–140. [Google Scholar] [CrossRef]
- Li, J.; Peng, J.; Zhang, Y.; Ji, Y.; Shi, H.; Mao, L.; Gao, S. Removal of triclosan via peroxidases-mediated reactions in water: Reaction kinetics, products and detoxification. J. Hazard. Mater. 2016, 310, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Alneyadi, A.H.; Ashraf, S.S. Differential enzymatic degradation of thiazole pollutants by two different peroxidases—A comparative study. Chem. Eng. J. 2016, 303, 529–538. [Google Scholar] [CrossRef]
- Duran, N.; Esposito, E. Potential applications of oxidative enzymes and phenoloxidase-like compounds in wastewater and soil treatment: A review. Appl. Catal. B Environ. 2000, 28, 83–99. [Google Scholar] [CrossRef]
- Jain, R.; Mathur, M.; Sikarwar, S.; Mittal, A. Removal of the hazardous dye Rhodamine B through photocatalytic and adsorption treatments. J. Environ. Manag. 2007, 85, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.-F.; Huang, Y.-P.; Liu, D.-F.; Huang, Y.; Guo, W.; David, J. Photocatalytic degradation of the dye Sulforhodamine-B: A comparative study of different light sources. J. Environ. Sci. 2007, 19, 97–102. [Google Scholar] [CrossRef]
- Gosetti, F.; Bolfi, B.; Marengo, E. Identification of Sulforhodamine B photodegradation products present in nonpermanent tattoos by micro liquid chromatography coupled with tandem high-resolution mass spectrometry. Anal. Bioanal. Chem. 2015, 407, 4649–4659. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Tang, X.; Sun, Y.; Wang, Y.; Long, Y.; Jiang, J.; Xu, H. Removal of Rhodamine B from wastewater by modified Volvariella volvacea: Batch and column study. RSC Adv. 2015, 5, 25337–25347. [Google Scholar] [CrossRef]
- Husain, M.; Husain, Q. Applications of redox mediators in the treatment of organic pollutants by using oxidoreductive enzymes: A review. Crit. Rev. Environ. Sci. Technol. 2007, 38, 1–42. [Google Scholar] [CrossRef]
- Van der Zee, F.P.; Cervantes, F.J. Impact and application of electron shuttles on the redox (bio)transformation of contaminants: A review. Biotechnol. Adv. 2009, 27, 256–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalsoom, U.; Ashraf, S.S.; Meetani, M.A.; Rauf, M.A.; Bhatti, H.N. Mechanistic study of a diazo dye degradation by soybean peroxidase. Chem. Cent. J. 2013, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ali, L.; Algaithi, R.; Habib, H.M.; Souka, U.; Rauf, M.A.; Ashraf, S.S. Soybean peroxidase-mediated degradation of an azo dye—A detailed mechanistic study. BMC Biochem. 2013, 14, 35. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.S. Raising environmental awareness through applied biochemistry laboratory experiments. Biochem. Mol. Biol. Educ. 2013, 41, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Marchis, T.; Avetta, P.; Bianco-Prevot, A.; Fabbri, D.; Viscardi, G.; Laurenti, E. Oxidative degradation of Remazol Turquoise Blue G 133 by soybean peroxidase. J. Inorg. Biochem. 2011, 105, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, J.; Tan, Y.; Jiang, Y.; Hu, M.; Li, S.; Zhai, Q. Rapid decolorization of anthraquinone and triphenylmethane dye using chloroperoxidase: Catalytic mechanism, analysis of products and degradation route. Chem. Eng. J. 2014, 244, 9–18. [Google Scholar] [CrossRef]
- Zhang, J.; Feng, M.; Jiang, Y.; Hu, M.; Li, S.; Zhai, Q. Efficient decolorization/degradation of aqueous azo dyes using buffered H2O2 oxidation catalyzed by a dosage below ppm level of chloroperoxidase. Chem. Eng. J. 2012, 191, 236–242. [Google Scholar] [CrossRef]
- Valderrama, B.; Ayala, M.; Vazquez-Duhalt, R. Suicide inactivation of peroxidases and the challenge of engineering more robust enzymes. Chem. Biol. 2002, 9, 555–565. [Google Scholar] [CrossRef]
- Silva, M.C.; Torres, J.A.; de Sá, L.R.V.; Chagas, P.M.B.; Ferreira-Leitão, V.S.; Corrêa, A.D. The use of soybean peroxidase in the decolourization of Remazol Brilliant Blue R and toxicological evaluation of its degradation products. J. Mol. Catal. B Enzym. 2013, 89, 122–129. [Google Scholar] [CrossRef]
- Shuler, M.L.; Kargi, F. Bioprocess Engineering Basic Concepts; Prentice-Hall: Englewood Cliffs, NJ, USA, 1992. [Google Scholar]
- McEldoon, J.P.; Dordick, J.S. Unusual thermal stability of soybean peroxidase. Biotechnol. Prog. 1996, 12, 555–558. [Google Scholar] [CrossRef]
- Wright, H.; Nicell, J.A. Characterization of soybean peroxidase for the treatment of aqueous phenols. Bioresour. Technol. 1999, 70, 69–79. [Google Scholar] [CrossRef]
- Ravi Dhas, C.; Venkatesh, R.; Jothivenkatachalam, K.; Nithya, A.; Benjamin, B.S.; Raj, A.M.E.; Jeyadheepan, K.; Sanjeeviraja, C. Visible light driven photocatalytic degradation of Rhodamine B and Direct Red using cobalt oxide nanoparticles. Ceram. Int. 2015, 41, 9301–9313. [Google Scholar] [CrossRef]
- Natarajan, T.S.; Thomas, M.; Natarajan, K.; Bajaj, H.C.; Tayade, R.J. Study on UV-led/TiO2 process for degradation of rhodamine B dye. Chem. Eng. J. 2011, 169, 126–134. [Google Scholar] [CrossRef]
- Luan, J.; Li, M.; Ma, K.; Li, Y.; Zou, Z. Photocatalytic activity of novel Y2InSbO7 and Y2GdSbO7 nanocatalysts for degradation of environmental pollutant Rhodamine B under visible light irradiation. Chem. Eng. J. 2011, 167, 162–171. [Google Scholar] [CrossRef]
- Bibi, I.; Bhatti, H.N.; Asgher, M. Comparative study of natural and synthetic phenolic compounds as efficient laccase mediators for the transformation of cationic dye. Biochem. Eng. J. 2011, 56, 225–231. [Google Scholar] [CrossRef]
- Nouren, S.; Bhatti, H.N. Mechanistic study of degradation of Basic Violet 3 by Citrus limon peroxidase and phytotoxicity assessment of its degradation products. Biochem. Eng. J. 2015, 95, 9–19. [Google Scholar] [CrossRef]
- Anastasi, A.; Coppola, T.; Prigione, V.; Varese, G.C. Pyrene degradation and detoxification in soil by a consortium of basidiomycetes isolated from compost: Role of laccases and peroxidases. J. Hazard. Mater. 2009, 165, 1229–1233. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Asgher, M.; Iqbal, M.; Hu, H.; Zhang, X. Chitosan beads immobilized manganese peroxidase catalytic potential for detoxification and decolorization of textile effluent. Int. J. Biol. Macromol. 2016, 89, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.C.; Corrêa, A.D.; Amorim, M.T.S.P.; Parpot, P.; Torres, J.A.; Chagas, P.M.B. Decolorization of the phthalocyanine dye Reactive Blue 21 by turnip peroxidase and assessment of its oxidation products. J. Mol. Catal. B Enzym. 2012, 77, 9–14. [Google Scholar] [CrossRef]
- Kunz, A.; Mansilla, H.; Durán, N. A degradation and toxicity study of three textile reactive dyes by ozone. Environ. Technol. 2002, 23, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Duhalt, R.; Ayala, M.; Márquez-Rocha, F.J. Biocatalytic chlorination of aromatic hydrocarbons by chloroperoxidase of Caldariomyces fumago. Phytochemistry 2001, 58, 929–933. [Google Scholar] [CrossRef]
- Garcia, J.C.; Simionato, J.I.; Almeida, V.D.C.; Palácio, S.M.; Rossi, F.L.; Schneider, M.V.; Souza, N.E.D. Evolutive follow-up of the photocatalytic degradation of real textile effluents in TiO2 and TiO2/H2O2 systems and their toxic effects on Lactuca sativa seedlings. J. Braz. Chem. Soc. 2009, 20, 1589–1597. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Optimum Conditions | SBP-Based Degradation | CPO-Based Degradation |
|---|---|---|
| [HOBT] (μM) | 50 | 167 |
| pH | 4 | 2 |
| [Enzyme] (pM) | 50 | 440 |
| [Dye] (ppm) | 10 | 6.25 |
| [H2O2] (mM) | 0.2 | 0.2 |
| Operational temperature range (°C) | 20–80 | 20–50 |
| Compound ID | m/z | RT (min) | SRB DYE + SBP | SRB DYE + CPO | ||
|---|---|---|---|---|---|---|
| Intensity | Intensity | |||||
| A | 102 | 12.56 | + | 0.5 × 104 | + | 4.0 × 104 |
| B | 116 | 14.56 | + | 1.0 × 104 | + | 6.0 × 104 |
| C | 120 | 13.46 | + | 7.5 × 104 | + | 9.0 × 104 |
| D | 168 | 2.84 | − | − | + | 1.4 × 104 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alneyadi, A.H.; Shah, I.; AbuQamar, S.F.; Ashraf, S.S. Differential Degradation and Detoxification of an Aromatic Pollutant by Two Different Peroxidases. Biomolecules 2017, 7, 31. https://doi.org/10.3390/biom7010031
Alneyadi AH, Shah I, AbuQamar SF, Ashraf SS. Differential Degradation and Detoxification of an Aromatic Pollutant by Two Different Peroxidases. Biomolecules. 2017; 7(1):31. https://doi.org/10.3390/biom7010031
Chicago/Turabian StyleAlneyadi, Aysha Hamad, Iltaf Shah, Synan F. AbuQamar, and Syed Salman Ashraf. 2017. "Differential Degradation and Detoxification of an Aromatic Pollutant by Two Different Peroxidases" Biomolecules 7, no. 1: 31. https://doi.org/10.3390/biom7010031
APA StyleAlneyadi, A. H., Shah, I., AbuQamar, S. F., & Ashraf, S. S. (2017). Differential Degradation and Detoxification of an Aromatic Pollutant by Two Different Peroxidases. Biomolecules, 7(1), 31. https://doi.org/10.3390/biom7010031