Identification of miRNAs and Their Response to Cold Stress in Astragalus Membranaceus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Cold Stress Treatment
2.2. Small RNA Library Construction and Sequencing
2.3. Transcriptome Sequences Assembly
2.4. Identification of Conserved and Non-Conserved miRNAs
2.5. qRT-PCR Analysis of miRNAs
2.6. Target Gene Prediction of Identified miRNAs and Gene Ontology Analysis of the Targets
2.7. qRT-PCR Analysis of the miRNA Targets
3. Results
3.1. Summary of Small RNA Library Dataset by Deep Sequencing in A. Membranaceus
3.2. Identification of Conserved miRNAs and their Stem-Loop Precursors in A. Membranaceus
3.3. Identification of Non-Conserved miRNAs in A. Membranaceus
3.4. Validation of the Identified miRNAs and their Expression Level in Leaves and Roots by qRT-PCR
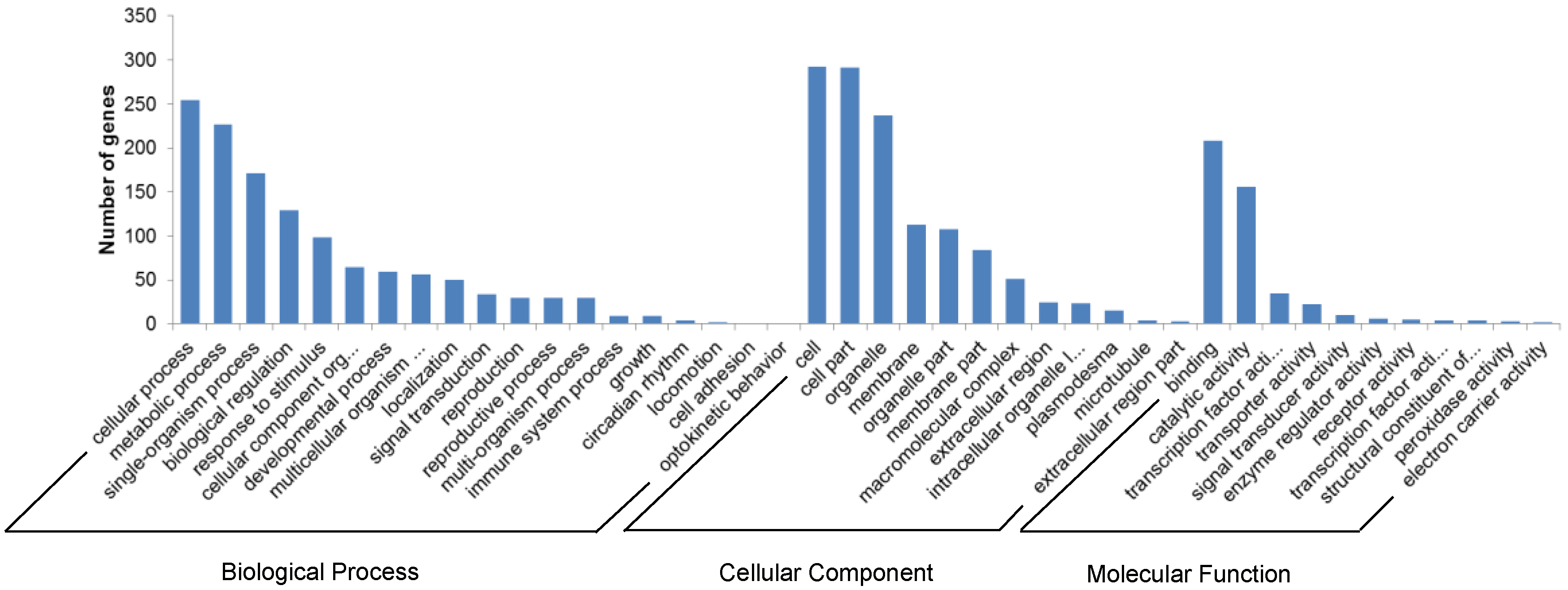
3.5. Target Prediction of A. Membranaceus miRNAs
3.6. Non-Coding RNAs Targeted by miRNAs in A. Membranaceus
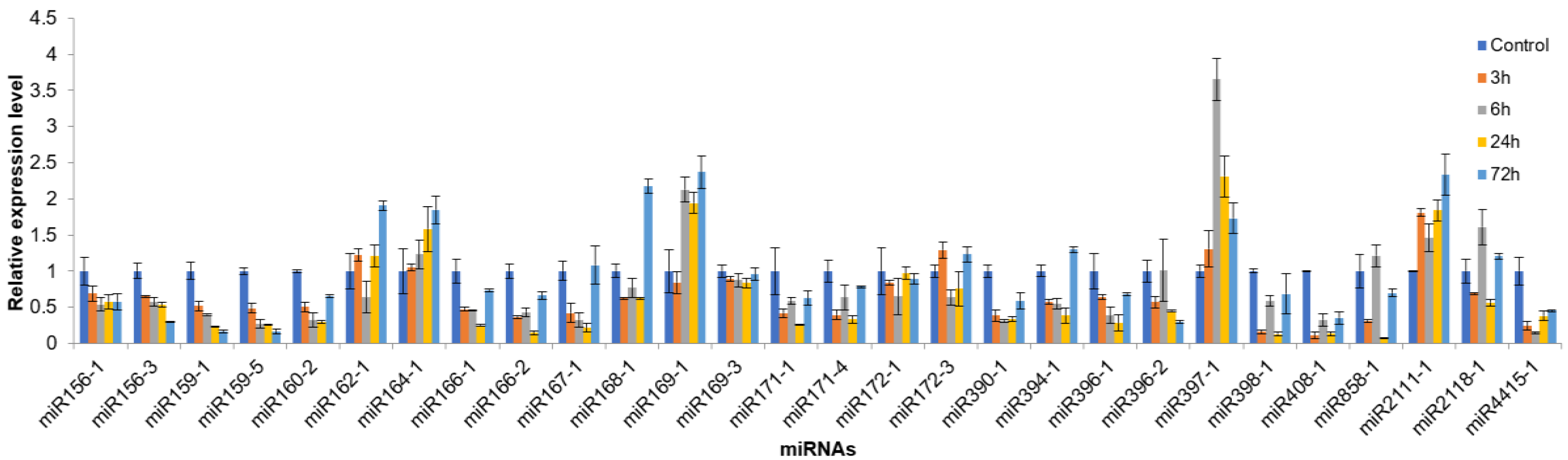
3.7. The Expression Pattern of miRNAs in A. Membranaceus Leaves under Cold Stress
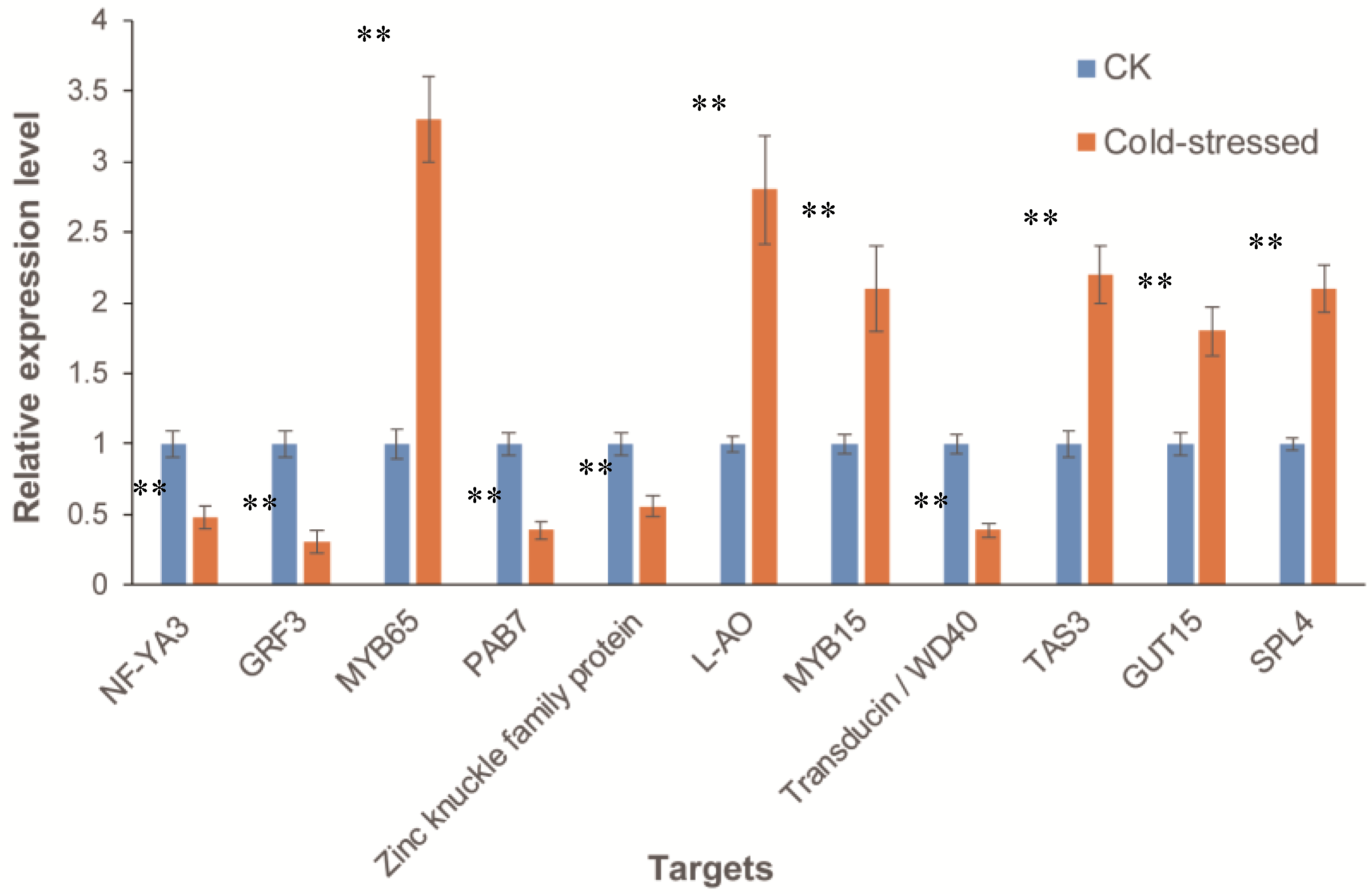
3.8. Expression of miRNA Targets in Response to Cold Stress in A. Membranaceus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nalbantsoy, A.; Nesil, T.; Yilmaz-Dilsiz, O.; Aksu, G.; Khan, S.; Bedir, E. Evaluation of the immunomodulatory properties in mice and in vitro anti-inflammatory activity of cycloartane type saponins from Astragalus species. J. Ethnopharmacol. 2012, 139, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.; Kim, E.H.; Chun, M.; Kang, S.; Shim, B.; Yu, Y.B.; Jeong, G.; Lee, J.S. Astragali Radix elicits anti-inflammation via activation of MKP-1, concomitant with attenuation of p38 and Erk. J. Ethnopharmacol. 2008, 115, 2008. [Google Scholar] [CrossRef] [PubMed]
- Leung, W.C.; Leung, K.N. In vitro and in vivo anti-tumor effects of Astragalus membranaceus. Cancer Lett. 2007, 252, 43–54. [Google Scholar]
- Chan, J.Y.; Lam, F.C.; Leung, P.C.; Che, C.T.; Feng, K.P. Antihyperglycemic and antioxidative effects of a herbal formulation of Radix Astragali, Radix Codonopsis and Cortex Lycii in a mouse model of type 2 diabetes mellitus. Phyther. Res. 2009, 23, 658–665. [Google Scholar] [CrossRef]
- Kim, C.; Ha, H.; Kim, J.S.; Kim, Y.T.; Kwon, S.C.; Park, S.W. Induction of growth hormone by the roots of Astragalus membranaceus in pituitary cell culture. Arch. Pharm. Res. 2003, 26, 34–39. [Google Scholar] [CrossRef]
- Liu, P.; Zhao, H.; Luo, Y. Anti-aging implications of Astragalus membranaceus (Huangqi): A well-known chinese tonic. Aging Dis. 2017, 8, 868–886. [Google Scholar] [CrossRef]
- Jiao, J.; Gai, Q.Y.; Wang, W.; Luo, M.; Gu, C.B.; Fu, Y.J.; Ma, W. Ultraviolet radiation-elicited enhancement of isoflavonoid accumulation, biosynthetic gene expression, and antioxidant activity in Astragalus membranaceus hairy root cultures. J. Agric. Food Chem. 2015, 63, 8216–8224. [Google Scholar] [CrossRef]
- Liu, R.; Xu, S.; Li, J.; Hu, Y.; Lin, Z. Expression profile of a PAL gene from Astragalus membranaceus var. Mongholicus and its crucial role in flux into flavonoid. Plant Cell Rep. 2006, 25, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Reis, R.S.; Eamens, A.L.; Waterhouse, P.M. Missing pieces in the puzzle of plant microRNAs. Trends plant sci. 2015, 20, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Zhang, L.; Xu, C.; Yuan, S.; Zhang, F.; Zheng, Y.; Zhao, C. Uncovering small RNA-mediated responses to cold stress in a wheat thermosensitive genic male-sterile. Plant Physiol. 2012, 159, 721–738. [Google Scholar] [CrossRef] [PubMed]
- Barrera-figueroa, B.E.; Gao, L.; Wu, Z.; Zhou, X.; Zhu, J.; Jin, H.; Liu, R.; Zhu, J.K. High throughput sequencing reveals novel and abiotic stress-regulated microRNAs in the inflorescences of rice. BMC Plant Biol. 2012, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, G.; Sutoh, K.; Zhu, J.; Zhang, W. Identification of cold-inducible microRNAs in plants by transcriptome analysis. Biochim. Biophys. Acta 2008, 1779, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wu, Z.; Jiang, F.; Zhou, R.; Yang, Z. Identification of chilling stress-responsive tomato microRNAs and their target genes by high-throughput sequencing and degradome analysis. BMC Genomics 2014, 15, 1130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, Y.; Huan, Q.; Chong, K. Deep sequencing of Brachypodium small RNAs at the global genome level identifies microRNAs involved in cold stress response. BMC Genomics 2009, 10, 449. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Wang, M.; Zhang, H.Y.; Liu, J.H. The miR396b of Poncirus trifoliata functions in cold tolerance by regulating ACC oxidase gene expression and modulating ethylene-polyamine. Plant Cell Physiol. 2016, 57, 1865–1878. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, D.; Mao, D.; Liu, X.; Ji, C.; Li, X.; Zhao, X.; Cheng, Z.; Chen, C.; Zhu, L. Overexpression of microRNA319 impacts leaf morphogenesis and leads to enhanced cold tolerance in rice (Oryza sativa L.). Plant Cell Environ. 2013, 36, 2207–2218. [Google Scholar] [CrossRef]
- Dong, C.H.; Pei, H. Over-expression of miR397 improves plant tolerance to cold stress in Arabidopsis thaliana. J. Plant Biol. 2014, 57, 209–217. [Google Scholar] [CrossRef]
- Fahlgren, N.; Howell, M.D.; Kasschau, K.D.; Chapman, E.J.; Sullivan, C.M.; Cumbie, J.S.; Givan, S.A.; Law, T.F.; Grant, S.R.; Dangl, J.L.; et al. High-throughput sequencing of Arabidopsis microRNAs: Evidence for frequent birth and death of MIRNA genes. PLoS ONE 2007, 2, e219. [Google Scholar] [CrossRef]
- Yin, Z.; Han, X.; Li, Y.; Wang, J.; Wang, D.; Wang, S.; Fu, X.; Ye, W. Comparative analysis of cotton small RNAs and their target genes in response to salt stress. Genes 2017, 8, 369. [Google Scholar] [CrossRef]
- Li, Y.; Wan, L.; Bi, S.; Wan, X.; Li, Z.; Cao, J.; Tong, Z.; Xu, H.; He, F.; Li, X. Identification of drought-responsive microRNAs from roots and leaves of alfalfa by high-throughput sequencing. Genes (Basel) 2017, 8, 119. [Google Scholar] [CrossRef]
- Article, L.B.; Coruh, C.; Cho, S.H.; Shahid, S.; Liu, Q.; Wierzbicki, A.; Axtell, M.J. Comprehensive annotation of Physcomitrella patens small RNA loci reveals that the heterochromatic short interfering RNA pathway is largely conserved in land plants. Plant Cell 2015, 27, 2148–2162. [Google Scholar]
- Chen, M.; Cao, Z. Genome-wide expression profiling of microRNAs in poplar upon infection with the foliar rust fungus Melampsora larici-populina. BMC Genomics 2015, 16, 696. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinformatics 2010, 11, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Meyers, B.C.; Axtell, M.J.; Bartel, B.; Bartel, D.P.; Baulcombe, D.; Bowman, J.L.; Cao, X.; Carrington, J.C.; Chen, X.; Green, P.J.; et al. Criteria for annotation of plant microRNAs. Plant Cell 2008, 20, 3186–3190. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C (T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Zhou, X.; Ling, Y.; Zhang, Z.; Su, Z. agriGO: A GO analysis toolkit for the agricultural community. Nucleic Acids Res. 2010, 38, 64–70. [Google Scholar] [CrossRef]
- Gao, F.; Wang, J.; Wei, S.; Li, Z.; Wang, N.; Li, H.; Feng, J.; Li, H.; Zhou, Y.; Zhang, F. Transcriptomic analysis of drought stress responses in Ammopiptanthus mongolicus leaves using the RNA-seq technique. PLoS ONE 2015, 10, e0124382. [Google Scholar] [CrossRef]
- Goettel, W.; Liu, Z.; Xia, J.; Zhang, W.; Zhao, P.X.; An, Y.C. Systems and evolutionary characterization of microRNAs and their underlying regulatory networks in soybean cotyledons. PLoS ONE 2014, 9, e86153. [Google Scholar] [CrossRef]
- Arenas-Huertero, C.; Pérez, B.; Rabanal, F.; Blanco-Melo, D.; De la Rosa, C.; Estrada-Navarrete, G.; Sanchez, F.; Covarrubias, A.A.; Reyes, J.L. Conserved and novel miRNAs in the legume Phaseolus vulgaris in response to stress. Plant Mol. Biol. 2009, 70, 385–401. [Google Scholar] [CrossRef] [PubMed]
- De Sousa Cardoso, T.C.; Portilho, L.G.; de Oliveira, C.L.; McKeown, P.C.; Maluf, W.R.; Gomes, L.A.; Teixeira, T.A.; do Amaral, L.R.; Spillane, C.; de Souza Gomes, M. Genome-wide identification and in silico characterisation of microRNAs, their targets and processing pathway genes in Phaseolus vulgaris L. Plant Biol. 2016, 18, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, A.W.; Shi, B.; Huang, C.; Langridge, P.; Baumann, U. Discovery of barley miRNAs through deep sequencing of short reads. BMC Genomics 2011, 12, 129. [Google Scholar] [CrossRef]
- Huang, Y.; Ren, H.T.; Xiong, J.L.; Gao, X.C.; Sun, X.H. Identification and characterization of known and novel microRNAs in three tissues of Chinese giant salamander base on deep sequencing approach. Genomics 2017, 109, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Sung, G.; Spatafora, J.W.; Carrington, J.C. Evolution of microRNA genes by inverted duplication of target gene sequences in Arabidopsis thaliana. Nat. Genet. 2005, 36, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, Y.; Wu, J.; Ding, B.; Fei, Z. A dynamic evolutionary and functional landscape of plant phased small interfering RNAs. BMC Biol. 2015, 13, 32. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Qu, X.; Jin, W. PhaseTank: Genome-wide computational identification of phasiRNAs and their regulatory cascades. Bioinformatics 2015, 31, 284–286. [Google Scholar] [CrossRef]
- Mb, S.; Moxon, S.; Mapleson, D.; Hc, W.; Mohorianu, I.; Folkes, L.; Schwach, F.; Dalmay, T.; Moulton, V. The UEA sRNA workbench: A suite of tools for analysing and visualizing next generation sequencing microRNA and small RNA datasets. Bioinformatics 2012, 28, 2059–2061. [Google Scholar]
- Talmor-Neiman, M.; Stav, R.; Klipcan, L.; Buxdorf, K.; Baulcombe, D.C.; Arazi, T. Identification of trans -acting siRNAs in moss and an RNA- dependent RNA polymerase required for their biogenesis. Plant J. 2006, 48, 511–521. [Google Scholar] [CrossRef]
- Axtell, M.J.; Jan, C.; Rajagopalan, R.; Bartel, D.P. A Two-hit trigger for siRNA biogenesis in plants. Cell 2006, 127, 565–577. [Google Scholar] [CrossRef]
- Plewka, P.; Thompson, A.; Szymanski, M.; Nuc, P.; Knop, K.; Rasinska, A.; Bialkowska, A.; Szweykowska-Kulinska, Z.; Karlowski, W.M.; Jarmolowski, A. A stable tRNA-like molecule is generated from the long noncoding RNA GUT15 in Arabidopsis. RNA Biol. 2018, 15, 726–738. [Google Scholar] [CrossRef]
- Morin, R.D.; Aksay, G.; Dolgosheina, E.; Ebhardt, H.A.; Magrini, V.; Mardis, E.R.; Sahinalp, S.C.; Unrau, P.J. Comparative analysis of the small RNA transcriptomes of Pinus contorta and Oryza sativa. Genome Res. 2008, 18, 571–584. [Google Scholar] [CrossRef]
- Szittya, G.; Moxon, S.; Santos, D.M.; Jing, R.; Fevereiro, M.P.S.; Moulton, V.; Dalmay, T. High-throughput sequencing of Medicago truncatula short RNAs identifies eight new miRNA families. BMC Genomics 2008, 9, 593. [Google Scholar] [CrossRef]
- Song, C.; Wang, C.; Zhang, C.; Korir, N.K.; Yu, H.; Ma, Z.; Fang, J. Deep sequencing discovery of novel and conserved microRNAs in trifoliate orange (Citrus trifoliata). BMC Genomics 2010, 11, 1–12. [Google Scholar] [CrossRef]
- Henderson, I.R.; Zhang, X.; Lu, C.; Johnson, L.; Meyers, B.C.; Green, P.J.; Jacobsen, S.E. Dissecting Arabidopsis thaliana DICER function in small RNA processing, gene silencing and DNA methylation patterning. Nat. Genet. 2006, 38, 721–725. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Zhang, B. MicroRNA: A new target for improving plant tolerance to abiotic stress. J. Exp. Bot. 2015, 66, 1749–1761. [Google Scholar] [CrossRef]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.; Weigel, D.; Poethig, R.S. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef]
- Cui, N.; Sun, X.; Sun, M.; Jia, B.; Duanmu, H.; Lv, D.; Duan, X.; Zhu, Y. Overexpression of OsmiR156k leads to reduced tolerance to cold stress in rice (Oryza Sativa). Mol. Breed. 2015, 35, 1–11. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, F.; Cao, H.; Peng, H.; Ni, Z.; Sun, Q.; Yao, Y. TamiR159 directed wheat TaGAMYB cleavage and its involvement in anther development and heat response. PLoS One 2012, 7, e48445. [Google Scholar] [CrossRef] [PubMed]
- Boualem, A.; Laporte, P.; Jovanovic, M.; Laffont, C.; Plet, J.; Combier, J.; Niebel, A.; Crespi, M.; Frugier, F. MicroRNA166 controls root and nodule development in Medicago truncatula. Plant J. 2008, 54, 876–887. [Google Scholar] [CrossRef] [PubMed]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef]
- Marin, E.; Jouannet, V.; Nussaume, L.; Crespi, M.D.; Maizel, A. miR390, Arabidopsis TAS3 tasiRNAs, and their AUXIN RESPONSE FACTOR targets define an autoregulatory network quantitatively regulating lateral root growth. Plant Cell 2010, 22, 1104–1117. [Google Scholar] [CrossRef]
- Zeng, Q.; Yang, C.; Ma, Q.; Li, X.; Dong, W.; Nian, H. Identification of wild soybean miRNAs and their target genes responsive to aluminum stress. BMC Plant Biol. 2012, 12, 1–16. [Google Scholar] [CrossRef]
- Kulcheski, F.R.; de Oliveira, L.F.; Molina, L.G.; Almerão, M.P.; Rodrigues, F.A.; Marcolino, J.; Barbosa, J.F.; Stolf-Moreira, R.; Nepomuceno, A.L.; Marcelino-Guimarães, F.C.; et al. Identification of novel soybean microRNAs involved in abiotic and biotic stresses. BMC Genomics 2011, 12, 1–17. [Google Scholar] [CrossRef]
- Fotopoulos, V.; Kanellis, A.K. Altered apoplastic ascorbate redox state in tobacco plants via ascorbate oxidase overexpression results in delayed dark-induced senescence in detached leaves. Plant Physiol. Biochem. 2013, 73, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Chezem, W.R.; Memon, A.; Li, F.; Weng, J.; Clay, N.K. SG2-Type R2R3-MYB transcription factor MYB15 controls defense-induced lignification and basal immunity in Arabidopsis. Plant Cell 2017, 29, 1907–1926. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, H.S.; Bahk, S.; An, J.; Yoo, Y.; Kim, J.; Chung, W.S. Phosphorylation of the transcriptional repressor MYB15 by mitogen-activated protein kinase 6 is required for freezing tolerance in Arabidopsis. Nucleic Acids Res. 2017, 45, 6613–6627. [Google Scholar] [CrossRef]
- Pastore, J.J.; Limpuangthip, A.; Yamaguchi, N.; Wu, M.; Sang, Y.; Han, S.; Malaspina, L.; Chavdaroff, N.; Yamaguchi, A.; Wagner, D. LATE MERISTEM IDENTITY2 acts together with LEAFY to activate APETALA1. Development 2011, 138, 3189–3198. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.F.; Li, W.F.; Xu, H.Y.; Qi, L.W.; Han, S.Y. Role of cin-miR2118 in drought stress responses in Caragana intermedia and Tobacco. Gene 2015, 574, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; He, H.; Yu, D. Identification of nitrogen starvation-responsive microRNAs in Arabidopsis thaliana. PLoS ONE 2012, 7, e48951. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Total | Percentage of Total (%) | Distinct | Percentage of Distinct (%) |
|---|---|---|---|---|
| Raw reads | 9,685,427 | 100.00 | 3,697,922 | 100 |
| 3ADT & length filter | 3,020,110 | 31.18 | 949,768 | 25.68 |
| Junk reads | 58,185 | 0.60 | 48,416 | 1.31 |
| Rfam | 840,690 | 8.68 | 100,245 | 2.71 |
| Repeats | 5584 | 0.06 | 1321 | 0.04 |
| Clean reads | 5,761,893 | 59.49 | 2,598,545 | 70.27 |
| rRNA | 625,393 | 6.46 | 59,659 | 0.62 |
| tRNA | 160,729 | 1.66 | 29,496 | 0.3 |
| snoRNA | 5296 | 0.05 | 1977 | 0.02 |
| snRNA | 7931 | 0.08 | 3860 | 0.04 |
| Other Rfam RNA | 41,341 | 0.43 | 5253 | 0.05 |
| miRNA Family | miRNA Name | miRNA Sequence (5′-3′) | Length (nt) | Representative Homologous miRNA | Total Reads | Conservation in Other Plant Specie | |||
|---|---|---|---|---|---|---|---|---|---|
| gma | ath | ptc | osa | ||||||
| MIR156 | ame-miR156-1 | ugacagaagagagugagcac | 20 | gma-miR156u | 13,342 | ++ | + | + | + |
| ame-miR156-2 | ugacagaagagagagagcac | 20 | gma-miR156b | 10 | ++ | + | + | + | |
| ame-miR156-3 | uugacagaagauagagagcac | 21 | gma-miR156d | 45,036 | ++ | ++ | ++ | + | |
| ame-miR156-4 | ugacagaagauagagagcac | 20 | gma-miR156d | 603 | + | + | + | + | |
| MIR159 | ame-miR159-1 | uuuggauugaagggagcucua | 21 | gma-miR159a-3p | 40,206 | ++ | ++ | ++ | + |
| ame-miR159-2 | uuuggauugaagggagcu | 18 | gma-miR159a-3p | 135 | ++ | ++ | ++ | ++ | |
| ame-miR159-3 | uuuggacugaagggagcuccu | 21 | csi-miR159d | 88 | + | + | + | + | |
| ame-miR159-4 | uuggacugaaggggccucuu | 20 | gma-miR319f | 48 | ++ | + | + | + | |
| ame-miR159-5 | uuggacugaagggagcuccc | 20 | ath-miR319a | 1459 | ++ | ++ | ++ | + | |
| ame-miR159-6 | uggacugaagggagcuccuuc | 21 | gma-miR319q | 437 | ++ | + | + | + | |
| MIR160 | ame-miR160-1 | ugccuggcucccugaaugcca | 21 | mtr-miR160c | 261 | + | + | ++ | ++ |
| ame-miR160-2 | ugccuggcucccuguaugcca | 21 | gma-miR160a-5p | 2374 | ++ | ++ | ++ | ++ | |
| ame-miR160-3 | gcguaugaggagccaagcaua | 21 | gma-miR160a-3p | 127 | ++ | + | + | + | |
| MIR162 | ame-miR162-1 | ucgauaaaccucugcauccag | 21 | ath-miR162a-3p | 961 | ++ | ++ | ++ | ++ |
| ame-miR162-2 | ggaggcagcgguucaucgauc | 21 | csi-miR162-5p | 60 | + | + | + | + | |
| MIR164 | ame-miR164-1 | uggagaagcagggcacgugca | 21 | ath-miR164a | 1584 | ++ | ++ | ++ | ++ |
| ame-miR164-2 | caugugccccucuuccccauc | 21 | zma-miR164c-3p | 41 | + | + | |||
| MIR166 | ame-miR166-1 | ucggaccaggcuucauucccc | 21 | gma-miR166d | 71,955 | ++ | + | + | + |
| ame-miR166-2 | ucggaccaggcuucauucccg | 21 | gma-miR166j-3p | 8165 | ++ | ||||
| ame-miR166-3 | ucucggaccaggcuucauucc | 21 | gma-miR166k | 7665 | ++ | + | + | + | |
| ame-miR166-4 | ggaauguugucuggcucgagg | 21 | gma-miR166a-5p | 276 | ++ | + | + | ||
| ame-miR166-5 | ggaauguuguuuggcucgagg | 21 | gma-miR166h-5p | 162 | ++ | + | + | ||
| MIR167_1 | ame-miR167-1 | ugaagcugccagcaugaucuga | 22 | gma-miR167g | 6608 | ++ | + | + | + |
| ame-miR167-2 | ugaagcugccagcaugaucug | 21 | gma-miR167c | 3720 | ++ | ++ | ++ | ++ | |
| ame-miR167-3 | ugaagcugccagcaugaucua | 21 | gma-miR167a | 2600 | ++ | ++ | ++ | ++ | |
| ame-miR167-4 | gucaugcugugacagccucacu | 22 | cas-miR167b | 210 | + | ||||
| ame-miR167-5 | agaucauguggcaguuucacc | 21 | ahy-miR167-3p | 68 | ++ | + | |||
| MIR168 | ame-miR168-1 | cccgccuugcaucaacugaau | 21 | aly-miR168a-3p | 478 | ++ | ++ | + | |
| ame-miR168-2 | ucgcuuggugcaggucggga | 20 | gma-miR168a | 30 | ++ | ++ | ++ | + | |
| MIR169_2 | ame-miR169-1 | cagccaaggaugacuugccgg | 21 | gma-miR169a | 105 | ++ | ++ | ++ | ++ |
| ame-miR169-2 | ugagccagggaugacuugccgg | 22 | gma-miR169d | 12 | + | + | + | + | |
| ame-miR169-3 | ugagccaaggaugacuugccgg | 22 | gma-miR169d | 47 | ++ | + | + | + | |
| ame-miR169-4 | ugcagccaaggaugacuugcc | 21 | gma-miR169b | 39 | + | + | + | + | |
| ame-miR169-5 | ggcaaguuggccuuggcuau | 20 | zma-miR169r-3p | 4 | + | + | + | ||
| MIR171 | ame-miR171-1 | ugauugagccgugccaauauc | 21 | gma-miR171e | 1419 | ++ | + | ++ | ++ |
| ame-miR171-2 | uugagccgcgccaauaucacu | 21 | gma-miR171k-3p | 439 | ++ | + | + | + | |
| ame-miR171-3 | uugagccgugccaauaucac | 20 | gma-miR171i-3p | 87 | + | + | + | + | |
| ame-miR171-4 | uugagccgcgucaauaucuca | 21 | gma-miR171m | 1713 | ++ | + | + | + | |
| ame-miR171-5 | agguauuggcgcgccucaauu | 21 | osa-miR171i-5p | 5 | + | + | + | + | |
| ame-miR171-6 | cgauguuggugagguucaauc | 21 | gma-miR171k-5p | 40 | ++ | + | + | + | |
| MIR172 | ame-miR172-1 | agaaucuugaugaugcugcau | 21 | gma-miR172a | 1122 | ++ | ++ | ++ | ++ |
| ame-miR172-2 | ggagcaucaucaagauucaca | 21 | aly-miR172c-5p | 5 | + | + | ++ | + | |
| ame-miR172-3 | agaaucuugaugaugcugcag | 21 | ath-miR172c | 133 | + | ++ | + | + | |
| ame-miR172-4 | gcagcaucaucaagauucaca | 21 | csi-miR172b-5p | 6 | ++ | + | + | + | |
| MIR390 | ame-miR390-1 | aagcucaggagggauagcgcc | 21 | gma-miR390a-5p | 350 | ++ | ++ | ++ | ++ |
| ame-miR390-2 | cgcuauccauccugaguuuca | 21 | gma-miR390a-3p | 11 | ++ | ++ | + | + | |
| MIR394 | ame-miR394-1 | uuggcauucuguccaccucc | 20 | gma-miR394c-5p | 546 | ++ | ++ | ++ | ++ |
| MIR396 | ame-miR396-1 | uuccacagcuuucuugaacuu | 21 | gma-miR396b-5p | 6105 | ++ | ++ | ++ | ++ |
| ame-miR396-2 | guucaauaaagcugugggaag | 21 | gma-miR396i-3p | 1717 | ++ | ++ | + | ++ | |
| ame-miR396-3 | cucaagaaagcugugggaga | 20 | gma-miR396b-3p | 1313 | + | + | + | + | |
| ame-miR396-4 | cuuccacagcuuucuugaacug | 22 | gma-miR396a-5p | 523 | + | + | + | + | |
| MIR397 | ame-miR397-1 | ucauugagugcagcguugaug | 21 | gma-miR397a | 579 | ++ | ++ | ++ | ++ |
| MIR398 | ame-miR398-1 | uguguucucaggucaccccuu | 21 | gma-miR398a | 3826 | ++ | ++ | ++ | ++ |
| ame-miR398-2 | gggucguccugagaccacaug | 21 | bra-miR398-5p | 13 | + | + | + | ||
| ame-miR398-3 | uguguucucaggucgccccug | 21 | gma-miR398c | 666 | ++ | + | ++ | ++ | |
| ame-miR398-4 | ggagugaaucugagaacacaag | 22 | gma-miR398b-5p | 440 | + | + | |||
| MIR408 | ame-miR408-1 | augcacugccucuucccuggc | 21 | gma-miR408a-3p | 3496 | ++ | ++ | ++ | + |
| MIR477 | ame-miR477-1 | ucccucaaaggcuuccaguau | 21 | ppt-miR477c | 31 | + | + | ||
| MIR530 | ame-miR530-1 | ucugcauuugcaccugcacuu | 21 | stu-miR530 | 49 | + | + | + | |
| MIR858 | ame-miR858-1 | cucguugucuguucgaccuug | 21 | csi-miR858-3p | 15 | + | |||
| MIR1514 | ame-miR1514-1 | uuuucauuuuuaaaauaggca | 21 | gma-miR408a-3p | 10 | + | |||
| MIR2111 | ame-miR2111-1 | uaaucugcauccugagguuu | 20 | gma-miR2111b | 197 | ++ | + | + | |
| ame-miR2111-2 | uaaucugcauccugagguu | 19 | gma-miR2111b | 14 | + | + | + | ||
| ame-miR2111-3 | uaaucugcauccugaggugu | 20 | gma-miR2111b | 95 | + | + | + | ||
| ame-miR2111-4 | guccuugggaugcagauuacc | 21 | gma-miR2111a | 82 | + | + | + | ||
| MIR2118 | ame-miR2118-1 | uuaccgauuccacccaugccuc | 21 | mtr-miR2118 | 850 | + | + | + | |
| MIR4415 | ame-miR4415-1 | uugauucucaucacaacuugg | 21 | gma-miR4415a-3p | 1879 | + | |||
| ame-miR4415-2 | auguugugaugggaaucaaug | 21 | gma-miR4415b-5p | 50 | + | ||||
| MIR5083 | ame-miR5083-1 | agacuacaauuaucugaucaau | 22 | osa-miR5083 | 15 | + | |||
| miRNA Name | miRNA Sequence | Length (nt) | Total Reads |
|---|---|---|---|
| ame-miRN-1 | guucaagucccuccuuccgc | 20 | 31 |
| ame-miRN-2 | uccguuguagucuaguugguc | 21 | 2043 |
| ame-miRN-3 | gucgauauguccgagugguuaag | 23 | 1399 |
| ame-miRN-4 | caauuuagucccugaaauugu | 21 | 26 |
| ame-miRN-5 | gggcauuuggucuagugguaug | 22 | 696 |
| ame-miRN-6 | gaagcggaugggggccggcg | 20 | 402 |
| ame-miRN-7 | guauuguaaguggcagagug | 20 | 307 |
| ame-miRN-8 | acaaaucuuugaaaacaggaa | 21 | 8 |
| ame-miRN-9 | ccuguuuucaaauauuuguac | 21 | 2 |
| ame-miRN-10 | uauaguuuguuugaugguag | 20 | 78 |
| ame-miRN-11 | ugaaucucaguggaucguggc | 21 | 69 |
| ame-miRN-12 | gcuuccauagcauagugguag | 21 | 21 |
| ame-miRN-13 | uucuaauuucuccucccuuuc | 21 | 7 |
| ame-miRN-14 | uuuuugguucauuaaauaacu | 21 | 6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abla, M.; Sun, H.; Li, Z.; Wei, C.; Gao, F.; Zhou, Y.; Feng, J. Identification of miRNAs and Their Response to Cold Stress in Astragalus Membranaceus. Biomolecules 2019, 9, 182. https://doi.org/10.3390/biom9050182
Abla M, Sun H, Li Z, Wei C, Gao F, Zhou Y, Feng J. Identification of miRNAs and Their Response to Cold Stress in Astragalus Membranaceus. Biomolecules. 2019; 9(5):182. https://doi.org/10.3390/biom9050182
Chicago/Turabian StyleAbla, Merhaba, Huigai Sun, Zhuyun Li, Chunxiang Wei, Fei Gao, Yijun Zhou, and Jinchao Feng. 2019. "Identification of miRNAs and Their Response to Cold Stress in Astragalus Membranaceus" Biomolecules 9, no. 5: 182. https://doi.org/10.3390/biom9050182
APA StyleAbla, M., Sun, H., Li, Z., Wei, C., Gao, F., Zhou, Y., & Feng, J. (2019). Identification of miRNAs and Their Response to Cold Stress in Astragalus Membranaceus. Biomolecules, 9(5), 182. https://doi.org/10.3390/biom9050182