The Effect of Anticoagulants, Temperature, and Time on the Human Plasma Metabolome and Lipidome from Healthy Donors as Determined by Liquid Chromatography-Mass Spectrometry

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Lipid Extraction and LC-MS
2.3. Metabolite Extraction and LC-MS
2.4. Data Preprocessing and Analysis
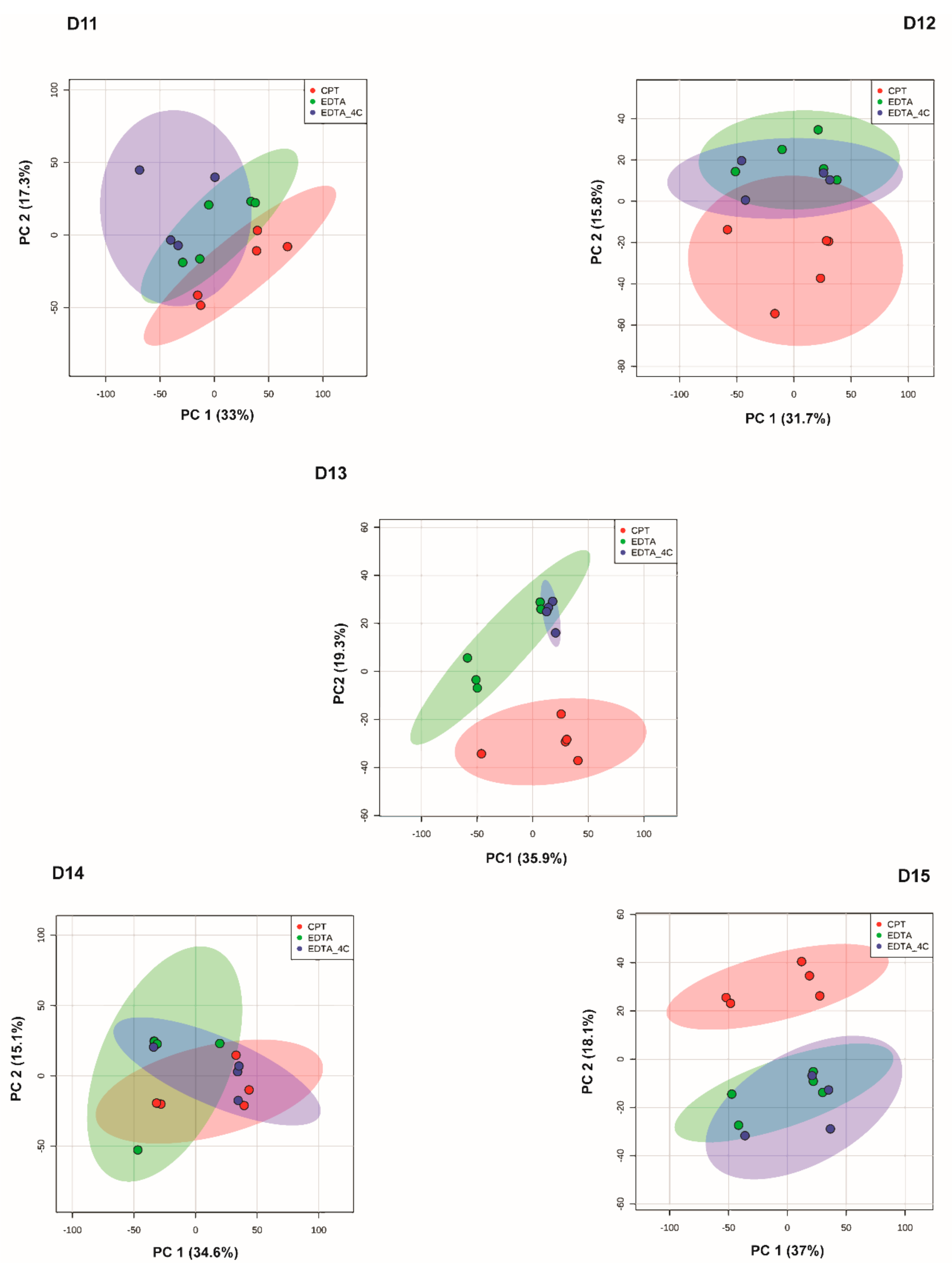
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lagarde, M.; Geloen, A.; Record, M.; Vance, D.; Spener, F. Lipidomics Is Emerging. Biochim. Et Biophys. Acta (Bba)—Bioenerg. 2003, 1634, 61. [Google Scholar] [CrossRef]
- Oliver, S.G. Functional Genomics: Lessons from Yeast. Philos Trans. R Soc. Lond B Biol. Sci. 2002, 357, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vazquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. Hmdb 4.0: The Human Metabolome Database for 2018. Nucleic Acids Res. 2017, 46, D608–D617. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A.; London, E. Functions of Lipid Rafts in Biological Membranes. Annu. Rev. Cell Dev. Biol. 1998, 14, 111–136. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Strum, J.C.; Bell, R.M. Lipid Biochemistry: Functions of Glycerolipids and Sphingolipids in Cellular Signaling. Faseb J. 1997, 11, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Gorgas, K.; Teigler, A.; Komljenovic, D.; Just, W.W. The Ether Lipid-Deficient Mouse: Tracking Down Plasmalogen Functions. Biochim. Et Biophys. Acta (Bba)-Bioenerg. 2006, 1763, 1511–1526. [Google Scholar] [CrossRef]
- Ikeda, M.; Kihara, A.; Igarashi, Y. Lipid Asymmetry of the Eukaryotic Plasma Membrane: Functions and Related Enzymes. Biol. Pharm. Bull. 2006, 29, 1542–1546. [Google Scholar] [CrossRef] [Green Version]
- Olzmann, J.A.; Carvalho, P. Dynamics and Functions of Lipid Droplets. Nat. Rev. Mol. Cell Biol. 2018. [Google Scholar] [CrossRef]
- Sillence, D.J. New Insights into Glycosphingolipid Functions--Storage, Lipid Rafts, and Translocators. Int. Rev. Cytol. 2007, 262, 151–189. [Google Scholar] [PubMed]
- Tazoe, F.; Yagyu, H.; Ishibashi, S. Functions of Enzymes Involved in Lipid Metabolism. Nihon Rinsho. Jpn. J. Clin. Med. 2007, 65, 26–30. [Google Scholar]
- Wanders, R.J.; van Grunsven, E.G.; Jansen, G.A. Lipid Metabolism in Peroxisomes: Enzymology, Functions and Dysfunctions of the Fatty Acid Alpha- and Beta-Oxidation Systems in Humans. Biochem. Soc. Trans. 2000, 28, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Han, X. Lipidomics for Studying Metabolism. Nat. Rev. Endocrinol. 2016, 12, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, J.; Heemels, M.T.; Shadan, S.; Weiss, U. Lipids in Health and Disease. Nature 2014, 510, 47. [Google Scholar] [CrossRef]
- Muro, E.; Atilla-Gokcumen, G.E.; Eggert, U.S. Lipids in Cell Biology: How Can We Understand Them Better? Mol. Biol. Cell 2014, 25, 1819–1823. [Google Scholar] [CrossRef] [PubMed]
- Mooradian, A.D. Dyslipidemia in Type 2 Diabetes Mellitus. Nat. Clin. Pract. : Endocrinol. Metab. 2009, 5, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Saghatelian, A.; Cravatt, B.F. Global Strategies to Integrate the Proteome and Metabolome. Curr. Opin. Chem. Biol. 2005, 9, 62–68. [Google Scholar] [CrossRef]
- Teahan, O.; Gamble, S.; Holmes, E.; Waxman, J.; Nicholson, J.K.; Bevan, C.; Keun, H.C. Impact of Analytical Bias in Metabonomic Studies of Human Blood Serum and Plasma. Anal. Chem. 2006, 78, 4307–4318. [Google Scholar] [CrossRef] [PubMed]
- Bando, K.; Kawahara, R.; Kunimatsu, T.; Sakai, J.; Kimura, J.; Funabashi, H.; Seki, T.; Bamba, T.; Fukusaki, E. Influences of Biofluid Sample Collection and Handling Procedures on Gc-Ms Based Metabolomic Studies. J. Biosci. Bioeng. 2010, 110, 491–499. [Google Scholar] [CrossRef]
- Barri, T.; Dragsted, L.O. Uplc-Esi-Qtof/Ms and Multivariate Data Analysis for Blood Plasma and Serum Metabolomics: Effect of Experimental Artefacts and Anticoagulant. Anal. Chim. Acta 2013, 768, 118–128. [Google Scholar] [CrossRef]
- Mei, H.; Hsieh, Y.; Nardo, C.; Xu, X.; Wang, S.; Ng, K.; Korfmacher, W.A. Investigation of Matrix Effects in Bioanalytical High-Performance Liquid Chromatography/Tandem Mass Spectrometric Assays: Application to Drug Discovery. Rapid Commun. Mass Spectrom 2003, 17, 97–103. [Google Scholar] [CrossRef]
- Yin, P.; Lehmann, R.; Xu, G. Effects of Pre-Analytical Processes on Blood Samples Used in Metabolomics Studies. Anal. Bioanal Chem. 2015, 407, 4879–4892. [Google Scholar] [CrossRef]
- Yin, P.; Peter, A.; Franken, H.; Zhao, X.; Neukamm, S.S.; Rosenbaum, L.; Lucio, M.; Zell, A.; Haring, H.U.; Xu, G.; et al. Preanalytical Aspects and Sample Quality Assessment in Metabolomics Studies of Human Blood. Clin. Chem. 2013, 59, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Wandro, S.; Carmody, L.; Gallagher, T.; LiPuma, J.J.; Whiteson, K. Making It Last: Storage Time and Temperature Have Differential Impacts on Metabolite Profiles of Airway Samples from Cystic Fibrosis Patients. Msystems 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Hernandes, V.V.; Barbas, C.; Dudzik, D. A Review of Blood Sample Handling and Pre-Processing for Metabolomics Studies. Electrophoresis 2017, 38, 2232–2241. [Google Scholar] [CrossRef] [PubMed]
- Haid, M.; Muschet, C.; Wahl, S.; Romisch-Margl, W.; Prehn, C.; Moller, G.; Adamski, J. Long-Term Stability of Human Plasma Metabolites During Storage at −80 Degrees C. J. Proteome Res. 2018, 17, 203–211. [Google Scholar] [CrossRef]
- Evans, M.J.; Livesey, J.H.; Ellis, M.J.; Yandle, T.G. Effect of Anticoagulants and Storage Temperatures on Stability of Plasma and Serum Hormones. Clin. Biochem. 2001, 34, 107–112. [Google Scholar] [CrossRef]
- Chuang, C.K.; Lin, S.P.; Lin, Y.T.; Huang, F.Y. Effects of Anticoagulants in Amino Acid Analysis: Comparisons of Heparin, Edta, and Sodium Citrate in Vacutainer Tubes for Plasma Preparation. Clin. Chem. 1998, 44, 1052–1056. [Google Scholar]
- Lopez-Bascon, M.A.; Calderon-Santiago, M.; Sanchez-Ceinos, J.; Fernandez-Vega, A.; Guzman-Ruiz, R.; Lopez-Miranda, J.; Malagon, M.M.; Priego-Capote, F. Influence of Sample Preparation on Lipidomics Analysis of Polar Lipids in Adipose Tissue. Talanta 2018, 177, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Bernini, P.; Bertini, I.; Luchinat, C.; Nincheri, P.; Staderini, S.; Turano, P. Standard Operating Procedures for Pre-Analytical Handling of Blood and Urine for Metabolomic Studies and Biobanks. J. Biomol. Nmr 2011, 49, 231–243. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Soltow, Q.A.; Strobel, F.H.; Mansfield, K.G.; Wachtman, L.; Park, Y.; Jones, D.P. High-Performance Metabolic Profiling with Dual Chromatography-Fourier-Transform Mass Spectrometry (Dc-Ftms) for Study of the Exposome. Metabolomics 2013, 9, S132–S143. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.M.; Walker, D.I.; Liang, Y.; Uppal, K.; Soltow, Q.A.; Tran, V.; Strobel, F.; Quyyumi, A.A.; Ziegler, T.R.; Pennell, K.D.; et al. Reference Standardization for Mass Spectrometry and High-Resolution Metabolomics Applications to Exposome Research. Toxicol Sci. 2015, 148, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Oresic, M. Mzmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef]
- Katajamaa, M.; Miettinen, J.; Oresic, M. Mzmine: Toolbox for Processing and Visualization of Mass Spectrometry Based Molecular Profile Data. Bioinformatics 2006, 22, 634–636. [Google Scholar] [CrossRef] [PubMed]
- Fahy, E.; Sud, M.; Cotter, D.; Subramaniam, S. Lipid Maps Online Tools for Lipid Research. Nucleic Acids Res. 2007, 35, W606–W612. [Google Scholar] [CrossRef]
- Koelmel, J.P.; Kroeger, N.M.; Ulmer, C.Z.; Bowden, J.A.; Patterson, R.E.; Cochran, J.A.; Beecher, C.W.W.; Garrett, T.J.; Yost, R.A. Lipidmatch: An Automated Workflow for Rule-Based Lipid Identification Using Untargeted High-Resolution Tandem Mass Spectrometry Data. BMC Bioinform. 2017, 18. [Google Scholar] [CrossRef]
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.; Shimizu, T.; Spener, F.; van Meer, G.; Wakelam, M.J.; Dennis, E.A. Update of the Lipid Maps Comprehensive Classification System for Lipids. J. Lipid Res. 2009, 50, S9–S14. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Park, Y.; Johnson, J.M.; Jones, D.P. Aplcms—Adaptive Processing of High-Resolution Lc/Ms Data. Bioinformatics 2009, 25, 1930–1936. [Google Scholar] [CrossRef]
- Uppal, K.; Soltow, Q.A.; Strobel, F.H.; Pittard, W.S.; Gernert, K.M.; Yu, T.; Jones, D.P. Xmsanalyzer: Automated Pipeline for Improved Feature Detection and Downstream Analysis of Large-Scale, Non-Targeted Metabolomics Data. BMC Bioinform. 2013, 14, 15. [Google Scholar] [CrossRef]
- Uppal, K.; Walker, D.I.; Jones, D.P. Xmsannotator: An R Package for Network-Based Annotation of High-Resolution Metabolomics Data. Anal. Chem. 2017, 89, 1063–1067. [Google Scholar] [CrossRef]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. Metaboanalyst: A Web Server for Metabolomic Data Analysis and Interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Using Metaboanalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2016, 55, 14101–141091. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. Metaboanalyst 4.0: Towards More Transparent and Integrative Metabolomics Analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed]
- Jorgenrud, B.; Jantti, S.; Mattila, I.; Poho, P.; Ronningen, K.S.; Yki-Jarvinen, H.; Oresic, M.; Hyotylainen, T. The Influence of Sample Collection Methodology and Sample Preprocessing on the Blood Metabolic Profile. Bioanalysis 2015, 7, 991–1006. [Google Scholar] [CrossRef] [PubMed]
- Zivkovic, A.M.; Wiest, M.M.; Nguyen, U.; Nording, M.L.; Watkins, S.M.; German, J.B. Assessing Individual Metabolic Responsiveness to a Lipid Challenge Using a Targeted Metabolomic Approach. Metabolomics 2009, 5, 209–218. [Google Scholar] [CrossRef]
- Yu, Z.; Kastenmuller, G.; He, Y.; Belcredi, P.; Moller, G.; Prehn, C.; Mendes, J.; Wahl, S.; Roemisch-Margl, W.; et al. Differences between Human Plasma and Serum Metabolite Profiles. PLoS ONE 2011, 6, e21230. [Google Scholar] [CrossRef] [PubMed]
- Hebels, D.G.; Georgiadis, P.; Keun, H.C.; Athersuch, T.J.; Vineis, P.; Vermeulen, R.; Portengen, L.; Bergdahl, I.A.; Hallmans, G.; et al. Kyrtopoulos, and Consortium EnviroGenomarkers Project. Performance in Omics Analyses of Blood Samples in Long-Term Storage: Opportunities for the Exploitation of Existing Biobanks in Environmental Health Research. Env. Health Perspect 2013, 121, 480–487. [Google Scholar] [CrossRef]
- Barton, R.H.; Nicholson, J.K.; Elliott, P.; Holmes, E. High-Throughput 1h Nmr-Based Metabolic Analysis of Human Serum and Urine for Large-Scale Epidemiological Studies: Validation Study. Int. J. Epidemiol. 2008, 37, 31–40. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Ellis, D.I.; Brown, M.; Halsall, A.; O’Hagan, S.; Spasic, I.; Tseng, A.; Kell, D.B. A Gc-Tof-Ms Study of the Stability of Serum and Urine Metabolomes During the Uk Biobank Sample Collection and Preparation Protocols. Int. J. Epidemiol. 2008, 37, 23–30. [Google Scholar] [CrossRef]
- Heiskanen, L.A.; Suoniemi, M.; Ta, H.X.; Tarasov, K.; Ekroos, K. Long-Term Performance and Stability of Molecular Shotgun Lipidomic Analysis of Human Plasma Samples. Anal. Chem. 2013, 85, 8757–8763. [Google Scholar] [CrossRef]
- Gika, H.G.; Theodoridis, G.A.; Wingate, J.E.; Wilson, I.D. Within-Day Reproducibility of an Hplc-Ms-Based Method for Metabonomic Analysis: Application to Human Urine. J. Proteome Res. 2007, 6, 3291–3303. [Google Scholar] [CrossRef]
- Deprez, S.; Sweatman, B.C.; Connor, S.C.; Haselden, J.N.; Waterfield, C.J. Optimisation of Collection, Storage and Preparation of Rat Plasma for 1h Nmr Spectroscopic Analysis in Toxicology Studies to Determine Inherent Variation in Biochemical Profiles. J. Pharm. Biomed. Anal. 2002, 30, 1297–1310. [Google Scholar] [CrossRef]
- Gonzalez-Covarrubias, V.; Dane, A.; Hankemeier, T.; Vreeken, R.J. The Influence of Citrate, Edta, and Heparin Anticoagulants to Human Plasma Lc-Ms Lipidomic Profiling. Metabolomics 2013, 9, 337–348. [Google Scholar] [CrossRef]
- Goossens, W.; Van Duppen, V.; Verwilghen, R.L. K2- or K3-Edta: The Anticoagulant of Choice in Routine Haematology? Clin. Lab. Haematol. 1991, 13, 291–295. [Google Scholar] [CrossRef]
- Lippi, G.; Giampaolo, L.; Guidi, G. Effects of Anticoagulants on Lipoprotein(a) Measurements with Four Commercial Assays. Eur. J. Clin. Chem. Clin. Biochem. 1996, 34, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Boyd, E.M.; Murray, R.B. The Lipid Content of Leucocytes from Heparinized Blood. Cmaj: Can. Med. Assoc. J. 1937, 37, 229–231. [Google Scholar]
- Eldon, M.B.; Ronald, B.M. The Effect of Anticoagulants on Blood Lipids. J. Biol. Chem. 1937, 117, 629–638. [Google Scholar]
- Denery, J.R.; Nunes, A.A.; Dickerson, T.J. Characterization of Differences between Blood Sample Matrices in Untargeted Metabolomics. Anal. Chem. 2011, 83, 1040–1047. [Google Scholar] [CrossRef] [PubMed]
- Kirwan, J.A.; Brennan, L.; Broadhurst, D.; Fiehn, O.; Cascante, M.; Dunn, W.B.; Schmidt, M.A.; Velagapudi, V. Preanalytical Processing and Biobanking Procedures of Biological Samples for Metabolomics Research: A White Paper, Community Perspective (for Precision Medicine and Pharmacometabolomics Task Group-the Metabolomics Society Initiative). Clin. Chem. 2018, 64, 1158–1182. [Google Scholar] [CrossRef]
- Devanapalli, B.; Bermingham, M.A.; Mahajan, D. Effect of Long-Term Storage at-80 Degrees C on the Various Lipid Parameters in Stored Plasma Samples. Clin. Chim. Acta 2002, 322, 179–181. [Google Scholar] [CrossRef]
- Reckless, J.P.D.; Betteridge, D.J.; Wu, P.; Payne, B.; Galton, D.J. Effect of Storage on Plasma-Lipoproteins. Lancet 1977, 2, 350. [Google Scholar] [CrossRef]
- Shih, W.J.; Bachorik, P.S.; Haga, J.A.; Myers, G.L.; Stein, E.A. Estimating the Long-Term Effects of Storage at-70 Degrees C on Cholesterol, Triglyceride, and Hdl-Cholesterol Measurements in Stored Sera. Clin. Chem. 2000, 46, 351–364. [Google Scholar]
- Tiedink, H.G.M.; Katan, M.B. Variability in Lipoprotein Concentrations in Serum after Prolonged Storage at -20-Degrees-C. Clin. Chim. Acta 1989, 180, 147–155. [Google Scholar] [CrossRef]
- Bachorik, P.S.; Walker, R.; Brownell, K.D.; Stunkard, A.J.; Kwiterovich, P.O. Determination of High Density Lipoprotein-Cholesterol in Stored Human Plasma. J. Lipid Res. 1980, 21, 608–616. [Google Scholar]
- Kamlage, B.; Neuber, S.; Bethan, B.; Gonzalez Maldonado, S.; Wagner-Golbs, A.; Peter, E.; Schmitz, O.; Schatz, P. Impact of Prolonged Blood Incubation and Extended Serum Storage at Room Temperature on the Human Serum Metabolome. Metabolites 2018, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Yuille, M.; Illig, T.; Hveem, K.; Schmitz, G.; Hansen, J.; Neumaier, M.; Tybring, G.; Wichmann, E.; Ollier, B. Laboratory Management of Samples in Biobanks: European Consensus Expert Group Report. Biopreserv Biobank 2010, 8, 65–69. [Google Scholar] [CrossRef]
- Cuhadar, S.; Koseoglu, M.; Atay, A.; Dirican, A. The Effect of Storage Time and Freeze-Thaw Cycles on the Stability of Serum Samples. Biochem. Med. Cas. Hrvat. Drus. Med. Biokem. 2013, 23, 70–77. [Google Scholar] [CrossRef]
- Pinto, J.; Domingues, M.R.M.; Galhano, E.; Pita, C.; Almeida, M.D.; Carreira, I.M.; Gil, A.M. Human Plasma Stability During Handling and Storage: Impact on Nmr Metabolomics. Analyst 2014, 139, 1168–1177. [Google Scholar] [CrossRef]
- Lee, J.E.; Kim, S.Y.; Shin, S.Y. Effect of Repeated Freezing and Thawing on Biomarker Stability in Plasma and Serum Samples. Osong Public Health Res. Perspect 2015, 6, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Yue, B.E.; Pattison, E.; Roberts, W.L.; Rockwood, A.L.; Danne, O.; Danne, O.; Lueders, C.; Mockel, M. Choline in Whole Blood and Plasma: Sample Preparation and Stability. Clin. Chem 2008, 54, 590–593. [Google Scholar] [CrossRef] [Green Version]
- Stokes, Y.M.; Salmond, C.E.; Carpenter, L.M.; Welby, T.J. Stability of Total Cholesterol, High-Density-Lipoprotein Cholesterol, and Triglycerides in Frozen Sera. Clin. Chem. 1986, 32, 995–999. [Google Scholar] [PubMed]
- Pottala, J.V.; Espeland, M.A.; Polreis, J.; Robinson, J.; Harris, W.S. Correcting the Effects of -20 Degrees C Storage and Aliquot Size on Erythrocyte Fatty Acid Content in the Women’s Health Initiative. Lipids 2012, 47, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Metherel, A.H.; Aristizabal Henao, J.J.; Stark, K.D. Epa and Dha Levels in Whole Blood Decrease More Rapidly When Stored at -20 Degrees C as Compared with Room Temperature, 4 and −75 Degrees C. Lipids 2013, 48, 1079–1091. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Year | Material | Method | Time | Temperature | Anticoagulant | Freeze/Thaw | Other | Conclusion |
|---|---|---|---|---|---|---|---|---|---|
| Teahan et al. [17] | 2006 | Serum and plasma | NMR | 2 h clot time | Room temperature and ice | Heparin | Yes | Variation due to individual. Freeze-thaw cycle should be minimal, no number of cycle recommended. Addition of heparin reduced the effect compared to non-heparinated serum or plasma samples. Serum clot contact time is the significant factor in introducing variation. Clotting on ice delayed the effect of temperature. | |
| Bando et al. [18] | 2010 | Plasma and urine | GC-MS | NA | Room temperature and ice | Plasma: K2EDTA, Sodium heparin | Citrate, 2-oxoglutarate, hippurate, threitol, threonate elevated in 4 h pooled sample. Heparinated plasma overlapped with other endogenous metabolites like sugars potentially causing inter-sample variation. EDTA did not overlap with other endogenous metabolites. Recommended the use of EDTA plasma for GC-MS analysis. | ||
| Barri et al. [19] | 2013 | Serum and plasma | LC-MS | NA | NA | K2EDTA, Li-Heparin, Na-citrate | Coagulation effect on serum led to release of peptides, hypoxanthine, and xanthine and can be nullified with robust data processing. Plasma anticoagulant can lead to ion suppression or enhancement on metabolites. Anticoagulant cation can make metabolites dominant in positive ESI/MS mode. Heparin preferred over others due to no observable matrix effects. EDTA and citrate plasma elicit sodium and potassium formate cluster causing ion suppression or enhancement of the coeluting metabolites. Blood serum is an alternative to avoid the anticoagulant matrix effect in the plasma sample. | ||
| Yin et al. [22] | 2013 | Serum and plasma | LC-MS | 2, 4, 8, and 24 h | Room temperature and ice | Heparin, EDTA | Yes | Hemolysis | L-carnitine significantly decreased after two to four cycle. No significant effect of up to two freeze-thaw cycles. Hypoxanthine, sphingosine-1-phosphate, and linolenyl carnitine significantly altered in the range of 0 h to 24 h. No change in metabolome up to 4 h when stored in ice. Sphingosine-1-P and hypoxanthine showed significant change after 4 h at room temperature. Significant hemolysis effect was observed in two species of lysophosphatidylcholines. Chemical noise pattern was observed in lithium heparinate and serum blood collection tubes. Polyethylene glycol ion cluster potentially leached out from the plastic bead in the collection tube. |
| Wandro et al. [23] | 2017 | Sputum | GC-MS | 4 °C and −20 °C | NA | Yes | Aspartic acid, glycine, isoleucine, serine, and uracil abundance increased after a day when stored at 4 °C. No effect at −20 °C. No effect of one to two freeze-thaw cycle. | ||
| Haid et al. [25] | 2018 | Plasma | LC-MS | Long term storage at −80 °C | NA | Increase in concentration for amino acids, hexoses, butyrylcarnitine, phospholipids containing more than 40 carbon. The decrease in concentration of acylcarnitines, lysophosphatidylcholines, diacyl-phosphatidylcholines, acyl-alkyl phosphatidylcholines, and sphingomyelin. | |||
| Jorgenrud et al. [44] | 2015 | Plasma and serum | GC-MS | Room temperature and 4 °C | EDTA, citrate | Amino acids higher in EDTA plasma, Amines abundance higher in serum and lowest in citrate plasma. Phenolic compounds abundance highest in EDTA and lowest in citrate plasma. Total carboxylic acid higher in citrate plasma compared to EDTA, Sterols, lactic acid, and serine abundances were lower in citrate compared to EDTA. No effect of temperature on lipids. | |||
| Mei et al. [20] | 2003 | Serum and plasma | LC-MS | Li-Heparin, Na-Heparin, Na2EDTA | Li-Heparin and polymers from the container showed matrix effect. | ||||
| Zivkovic et al. [45] | 2009 | Serum | GC-FID | 4 °C, −20 °C, −80 °C | 0–4% of metabolites affected in most lipid classes when stored for a week at 4 °C, −20 °C and −80 °C | ||||
| Yu et al. [46] | 2011 | Serum and plasma | FIA-MS | Reproducibility comparatively better in plasma. Arginine, PC (38:1), LPC (16:0, 17:0, 18:0, 18:1), serine, phenylalanine, glycine were 20%–26% higher in serum compared to plasma. | |||||
| Hebels et al. [47] | 2013 | Plasma | LC-MS | 0 h to 24 h | Room temperature and −80 °C | Heparin, EDTA, citrate | No effect of storage time on metabolites. <1% metabolites significantly different at FDR<0.05. | ||
| Barton et al. [48] | 2008 | Serum and urine | NMR | 0 h to 36 h | Plasma and urine metabolic profiles are not affected when stored at 4 °C up to 24 h. | ||||
| Dunn et al. [49] | 2008 | Serum and urine | GC-MS | 0 h to 24 h | No significant changes in metabolome was observed at two different storage time at 4 °C. | ||||
| Heiskanen et al. [50] | 2013 | Plasma | Shotgun MS | Plasma at −80 °C monitored for 42 months | Plasma sample volume (5 and 10 µL) | The higher plasma volume provided more stability to lipid concentration. The storage time did not have an effect on lipid stability. | |||
| Gika et al. [51] | 2007 | Urine | LC-MS | 1 month | Two temperatures −20 °C and -80 °C | Yes | Urine extract in autosampler for 20 h at 4 °C | No detectable effect on metabolites at two different temperatures for a month. Sample stable for at least 20 h at 4 °C in the autosampler. The sample can withstand one to nine freeze-thaw cycles without significant effect on metabolites. | |
| Deprez et al. [52] | 2002 | Plasma | NMR | 0–9 month | 4 °C and room temperature | No change in metabolite profile when snap frozen and stored at -80 °C for 9 months. A significant increase was observed in tyrosine, phenylalanine and glycerol for the sample at room temperature. Choline, 3-hydroxybtyurate, acetate, glycerol slightly increase when stored at 4 °C for 3–4 days likely due to enzymatic action. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khadka, M.; Todor, A.; Maner-Smith, K.M.; Colucci, J.K.; Tran, V.; Gaul, D.A.; Anderson, E.J.; Natrajan, M.S.; Rouphael, N.; Mulligan, M.J.; et al. The Effect of Anticoagulants, Temperature, and Time on the Human Plasma Metabolome and Lipidome from Healthy Donors as Determined by Liquid Chromatography-Mass Spectrometry. Biomolecules 2019, 9, 200. https://doi.org/10.3390/biom9050200
Khadka M, Todor A, Maner-Smith KM, Colucci JK, Tran V, Gaul DA, Anderson EJ, Natrajan MS, Rouphael N, Mulligan MJ, et al. The Effect of Anticoagulants, Temperature, and Time on the Human Plasma Metabolome and Lipidome from Healthy Donors as Determined by Liquid Chromatography-Mass Spectrometry. Biomolecules. 2019; 9(5):200. https://doi.org/10.3390/biom9050200
Chicago/Turabian StyleKhadka, Manoj, Andrei Todor, Kristal M. Maner-Smith, Jennifer K. Colucci, ViLinh Tran, David A. Gaul, Evan J. Anderson, Muktha S. Natrajan, Nadine Rouphael, Mark J. Mulligan, and et al. 2019. "The Effect of Anticoagulants, Temperature, and Time on the Human Plasma Metabolome and Lipidome from Healthy Donors as Determined by Liquid Chromatography-Mass Spectrometry" Biomolecules 9, no. 5: 200. https://doi.org/10.3390/biom9050200
APA StyleKhadka, M., Todor, A., Maner-Smith, K. M., Colucci, J. K., Tran, V., Gaul, D. A., Anderson, E. J., Natrajan, M. S., Rouphael, N., Mulligan, M. J., McDonald, C. E., Suthar, M., Li, S., & Ortlund, E. A. (2019). The Effect of Anticoagulants, Temperature, and Time on the Human Plasma Metabolome and Lipidome from Healthy Donors as Determined by Liquid Chromatography-Mass Spectrometry. Biomolecules, 9(5), 200. https://doi.org/10.3390/biom9050200