Ligand Binding and Signaling of HARE/Stabilin-2


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
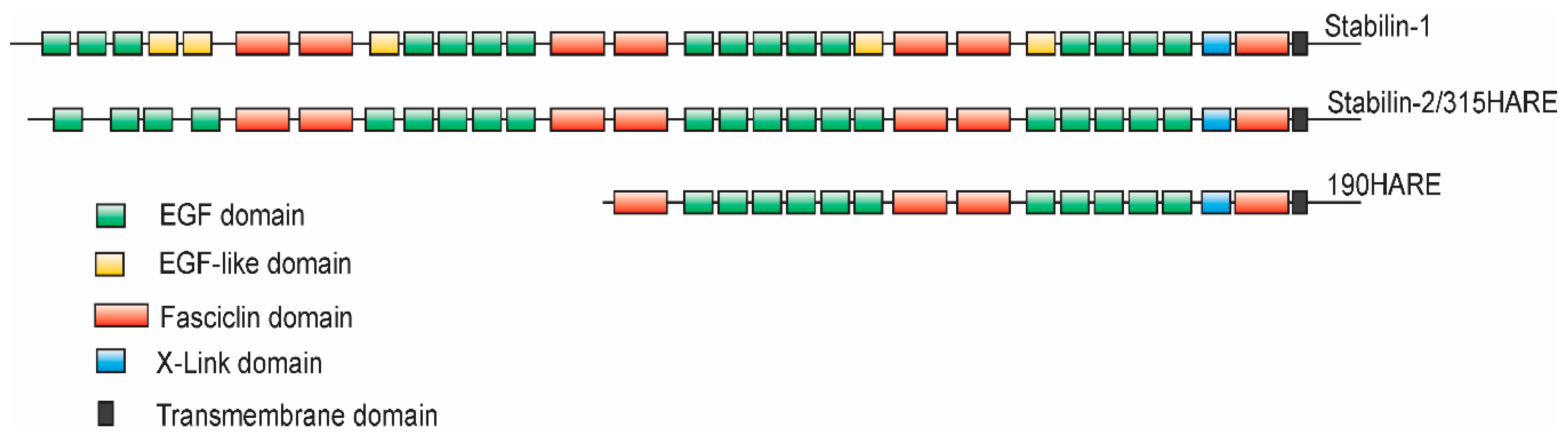
:1. Initial Discovery
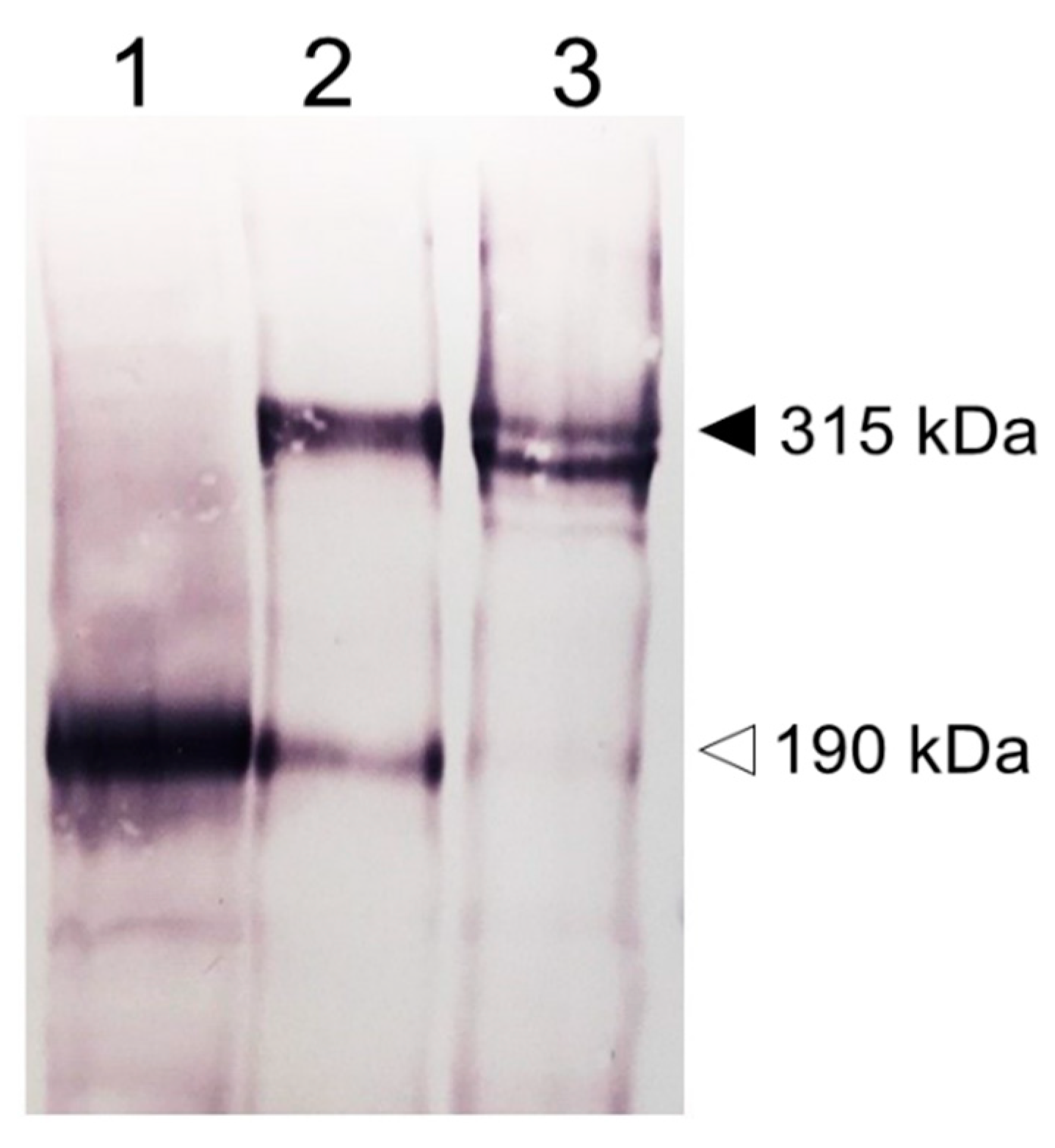
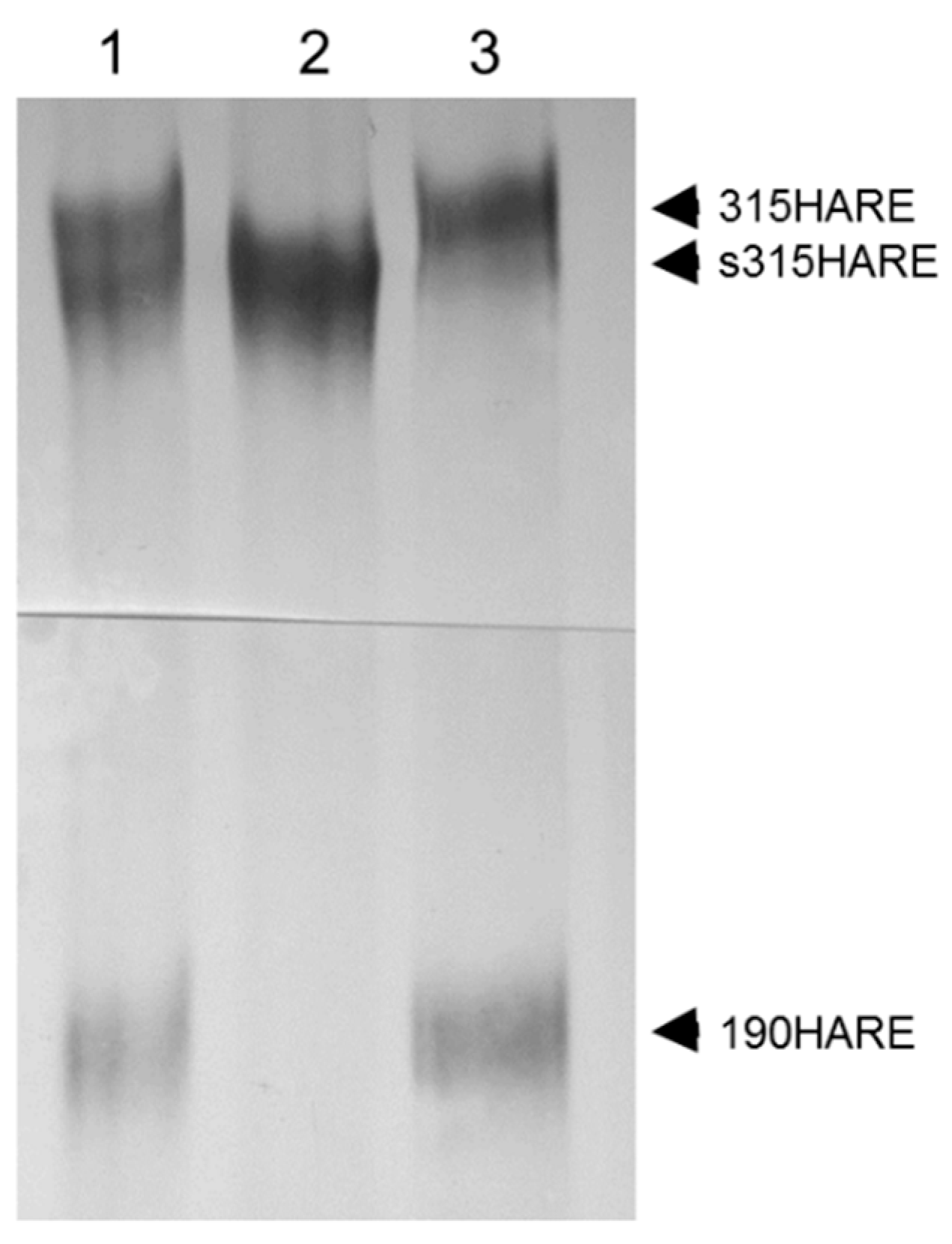
2. Tissue Expression
3. Hyaluronan Binding and Endocytosis
4. Heparin Clearance by Stabilin-1 and Stabilin-2
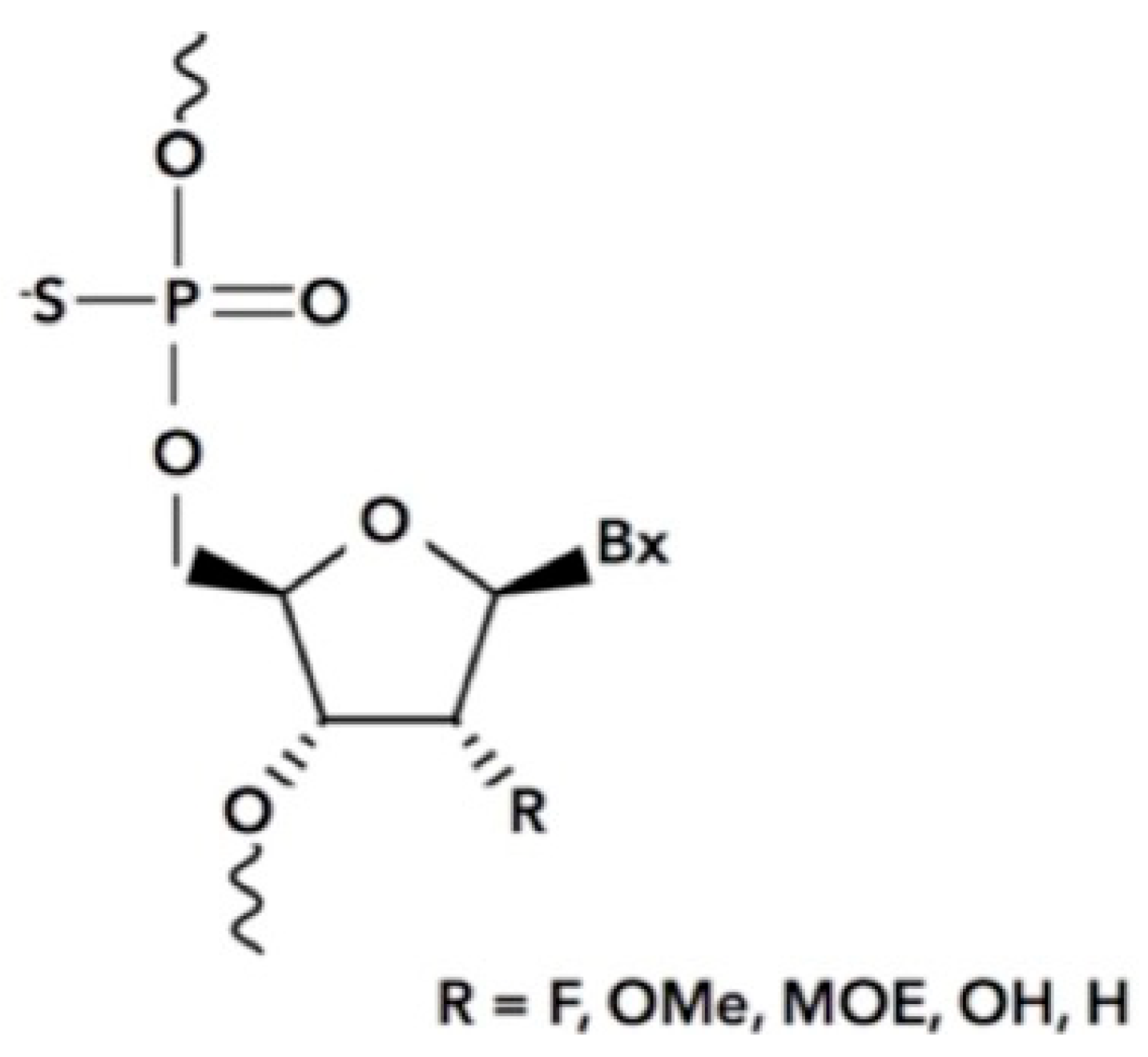
5. Stabilin Interactions with Antisense Oligonucleotides (ASOs)
6. Receptor Signaling
7. Conclusions/Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Goerdt, S. Identification of a novel high molecular weight protein preferentially expressed by sinusoidal endothelial cells in normal human tissues. J. Cell Biol. 1991, 113, 1425–1437. [Google Scholar] [CrossRef] [PubMed]
- Goerdt, S.; Bhardwaj, R.; Sorg, C. Inducible expression of MS-1 high-molecular-weight protein by endothelial cells of continuous origin and by dendritic cells/macrophages in vivo and in vitro. Am. J. Pathol. 1993, 142, 1409–1422. [Google Scholar] [PubMed]
- Politz, O.; Gratchev, A.; McCourt, P.A.G.; Schledzewski, K.; Guillot, P.; Johansson, S.; Svineng, G.; Franke, P.; Kannicht, C.; Kzhyshkowska, J.; et al. Stabilin-1 and -2 constitute a novel family of fasciclin-like hyaluronan receptor homologues. Biochem. J. 2002, 362 Pt 1, 155–164. [Google Scholar]
- Blume-Peytavi, U.; Adler, Y.D.; Geilen, C.C.; Ahmad, W.; Christiano, A.; Goerdt, S.; Orfanos, C.E. Multiple familial cutaneous glomangioma: A pedigree of 4 generations and critical analysis of histologic and genetic differences of glomus tumors. J. Am. Acad. Dermatol. 2000, 42, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Yannariello-Brown, J.; Zhou, B.; Ritchie, D.; Oka, J.A.; Weigel, P.H. A novel ligand blot assay detects different hyaluronan-binding proteins in rat liver hepatocytes and sinusoidal endothelial cells. Biochem. Biophys. Res. Commun. 1996, 218, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Yannariello-Brown, J.; Zhou, B.; Weigel, P.H. Identification of a 175 kDa protein as the ligand-binding subunit of the rat liver sinusoidal endothelial cell hyaluronan receptor. Glycobiology 1997, 7, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Weigel, J.A.; Fauss, L.; Weigel, P.H. Identification of the hyaluronan receptor for endocytosis (HARE). J. Biol. Chem. 2000, 275, 37733–37741. [Google Scholar] [CrossRef]
- Tamura, Y.; Adachi, H.; Osuga, J.I.; Ohashi, K.; Yahagi, N.; Sekiya, M.; Okazaki, H.; Tomita, S.; Iizuka, Y.; Shimano, H. FEEL-1 and FEEL-2 are endocytic receptors for advanced glycation end products. J. Biol. Chem. 2003, 278, 12613–12617. [Google Scholar] [CrossRef]
- Irjala, H.; Alanen, K.; Grénman, R.; Heikkilä, P.; Joensuu, H.; Jalkanen, S. Mannose receptor (MR) and common lymphatic endothelial and vascular endothelial receptor (CLEVER)-1 direct the binding of cancer cells to the lymph vessel endothelium. Cancer Res. 2003, 63, 4671–4676. [Google Scholar]
- Letunic, I.; Doerks, T.; Bork, P. SMART: Recent updates, new developments and status in 2015. Nucleic Acids Res. 2015, 43, D257–D260. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Oka, J.A.; Singh, A.; Weigel, P.H. Purification and subunit characterization of the rat liver endocytic hyaluronan receptor. J. Biol. Chem. 1999, 274, 33831–33834. [Google Scholar] [CrossRef] [PubMed]
- Kzhyshkowska, J.; Gratchev, A.; Martens, J.-H.; Pervushina, O.; Mamidi, S.; Johansson, S.; Schledzewski, K.; Hansen, B.; He, X.; Tang, J.; et al. Stabilin-1 localizes to endosomes and the trans-Golgi network in human macrophages and interacts with GGA adaptors. J. Leukoc. Biol. 2004, 76, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Prevo, R.; Banerji, S.; Ni, J.; Jackson, D.G. Rapid plasma membrane-endosomal trafficking of the lymph node sinus and high endothelial venule scavenger receptor/homing receptor stabilin-1 (FEEL-1/CLEVER-1). J. Biol. Chem. 2004, 279, 52580–52592. [Google Scholar] [CrossRef]
- Harris, E.N.; Kyosseva, S.V.; Weigel, J.A.; Weigel, P.H. Expression, processing, and glycosaminoglycan binding activity of the recombinant human 315-kDa hyaluronic acid receptor for endocytosis (HARE). J. Biol. Chem. 2007, 282, 2785–2797. [Google Scholar] [CrossRef]
- Weigel, J.A.; Zhang, H.; Bhargava, K.; Keszler, A.; Feix, J.; Hogg, N.; Joseph, J.; Kalyanaraman, B. A blocking antibody to the hyaluronan receptor for endocytosis (HARE) inhibits hyaluronan clearance by perfused liver. J. Biol. Chem. 2003, 278, 9808–9812. [Google Scholar] [CrossRef]
- Falkowski, M.; Schledzewski, K.; Hansen, B.; Goerdt, S. Expression of stabilin-2, a novel fasciclin-like hyaluronan receptor protein, in murine sinusoidal endothelia, avascular tissues, and at solid/liquid interfaces. Histochem. Cell Biol. 2003, 120, 361–369. [Google Scholar] [CrossRef]
- Park, S.-Y.; Yun, Y.; Lim, J.-S.; Kim, M.-J.; Kim, S.-Y.; Kim, J.-E.; Kim, I.-S. Stabilin-2 modulates the efficiency of myoblast fusion during myogenic differentiation and muscle regeneration. Nat. Commun. 2016, 7, 10871. [Google Scholar] [CrossRef] [Green Version]
- GTExPortal, in Webpage. The Broad Institute of MIT and Harvard. 2015. Available online: http://www.gtexportal.org/home/ (accessed on 8 July 2015).
- Hare, A.K.; Harris, E.N. Tissue-specific splice variants of HARE/Stabilin-2 are expressed in bone marrow, lymph node, and spleen. Biochem. Biophys. Res. Commun. 2015, 456, 257–261. [Google Scholar] [CrossRef]
- Rodriguez-Martinez, H.; Tienthai, P.; Atikuzzaman, M.; Carrillo, A.V.; Rubér, M.; Alvarez-Rodriguez, M. The ubiquitous hyaluronan: Functionally implicated in the oviduct? Theriogenology 2016, 86, 182–186. [Google Scholar] [CrossRef]
- Euppayo, T.; Punyapornwithaya, V.; Chomdej, S.; Ongchai, S.; Nganvongpanit, K. Effects of hyaluronic acid combined with anti-inflammatory drugs compared with hyaluronic acid alone, in clinical trials and experiments in osteoarthritis: A systematic review and meta-analysis. BMC Musculoskelet. Disord. 2017, 18, 387. [Google Scholar] [CrossRef] [PubMed]
- Balazs, E.A. Fine structure and function of ocular tissues. Int. Ophthalmol. Clin. 1973, 13, 169–187. [Google Scholar] [PubMed]
- Robert, L. Hyaluronan, a truly “youthful” polysaccharide. Its Med Appl. Pathol Biol (Paris) 2015, 63, 32–34. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.B.; El-Ashry, D.; Turley, E.A. Hyaluronan, Cancer-Associated Fibroblasts and the Tumor Microenvironment in Malignant Progression. Front. Cell Dev. Biol. 2018, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.R.; Laurent, T.C.; Laurent, U.B. Hyaluronan: Its nature, distribution, functions and turnover. J. Intern. Med. 1997, 242, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.R.F.; Laurent, T.C.; Pertoft, H.; Baxter, E. Plasma clearance, tissue distribution and metabolism of hyaluronic acid injected intravenously in the rabbit. Biochem. J. 1981, 200, 415–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, J.R.; Appelgren, L.E.; Laurent, T.C. Tissue uptake of circulating hyaluronic acid. A whole body autoradiographic study. Cell Tissue Res. 1983, 233, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.; Fraser, J.E.; Laurent, T.C.; Pertoft, H.; Smedsrød, B. Endothelial cells are a site of uptake and degradation of hyaluronic acid in the liver. Exp. Cell Res. 1983, 144, 223–228. [Google Scholar] [CrossRef]
- Harris, E.N.; Weigel, J.A.; Weigel, P.H. Endocytic function, glycosaminoglycan specificity, and antibody sensitivity of the recombinant human 190-kDa hyaluronan receptor for endocytosis (HARE). J. Biol. Chem. 2004, 279, 36201–36209. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Weigel, J.A.; Saxena, A.; Weigel, P.H. Molecular cloning and functional expression of the rat 175-kDa hyaluronan receptor for endocytosis. Mol. Biol. Cell 2002, 13, 2853–2868. [Google Scholar] [CrossRef] [PubMed]
- McGary, C.T.; Raja, R.H.; Weigel, P.H. Endocytosis of hyaluronic acid by rat liver endothelial cells. Evidence for receptor recycling. Biochem. J. 1989, 257, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Kyosseva, S.V.; Harris, E.N.; Weigel, P.H. The hyaluronan receptor for endocytosis mediates hyaluronan-dependent signal transduction via extracellular signal-regulated kinases. J. Biol. Chem. 2008, 283, 15047–15055. [Google Scholar] [CrossRef] [PubMed]
- Bayliss, M.; Howat, S.; Dudhia, J.; Murphy, J.; Barry, F.; Edwards, J.; Day, A. Up-regulation and differential expression of the hyaluronan-binding protein TSG-6 in cartilage and synovium in rheumatoid arthritis and osteoarthritis. Osteoarthr. Cartil. 2001, 9, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Glant, T.T.; Kamath, R.V.; Bardos, T.; Gal, I.; Szanto, S.; Murad, Y.M.; Sandy, J.D.; Mort, J.S.; Roughley, P.J.; Mikecz, K. Cartilage-specific constitutive expression of TSG-6 protein (product of tumor necrosis factor alpha-stimulated gene 6) provides a chondroprotective, but not antiinflammatory, effect in antigen-induced arthritis. Arthritis Rheum. 2002, 46, 2207–2218. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, D.; Mukhopadhyay, D.; Hascall, V.C.; Day, A.J.; Salustri, A.; Fulop, C. Two distinct populations of tumor necrosis factor-stimulated gene-6 protein in the extracellular matrix of expanded mouse cumulus cell-oocyte complexes. Arch. Biochem. Biophys. 2001, 394, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Blundell, C.D.; Almond, A.; Mahoney, D.J.; DeAngelis, P.L.; Campbell, I.D.; Day, A.J. Towards a structure for a TSG-6.hyaluronan complex by modeling and NMR spectroscopy: Insights into other members of the link module superfamily. J. Biol. Chem. 2005, 280, 18189–18201. [Google Scholar] [PubMed]
- Higman, V.A.; Briggs, D.C.; Mahoney, D.J.; Blundell, C.D.; Sattelle, B.M.; Dyer, D.P.; Green, D.E.; DeAngelis, P.L.; Almond, A.; Milner, C.M.; et al. A refined model for the TSG-6 link module in complex with hyaluronan: Use of defined oligosaccharides to probe structure and function. J. Biol. Chem. 2014, 289, 5619–5634. [Google Scholar] [CrossRef]
- Bano, F.; Tammi, M.I.; Kang, D.W.; Harris, E.N.; Richter, R.P. Single-Molecule Unbinding Forces between the Polysaccharide Hyaluronan and Its Binding Proteins. Biophys. J. 2018, 114, 2910–2922. [Google Scholar] [CrossRef] [Green Version]
- Heffelfinger, S.C.; Hawkins, H.H.; Barrish, J.; Taylor, L.; Darlington, G.J. SK HEP-1: A human cell line of endothelial origin. Vitr. Cell. Dev. Biol. 1992, 28, 136–142. [Google Scholar] [CrossRef]
- Weigel, J.A.; Weigel, P.H. Characterization of the recombinant rat 175-kDa hyaluronan receptor for endocytosis (HARE). J. Biol. Chem. 2003, 278, 42802–42811. [Google Scholar] [CrossRef]
- McGary, C.T.; Yannariello-Brown, J.; Kim, D.W.; Stinson, T.C.; Weigel, P.H. Degradation and intracellular accumulation of a residualizing hyaluronan derivative by liver endothelial cells. Hepatology 1993, 18, 1465–1476. [Google Scholar] [CrossRef] [PubMed]
- Smedsrod, B.; Pertoft, H.; Eriksson, S.; Fraser, J.R.; Laurent, T.C. Studies in vitro on the uptake and degradation of sodium hyaluronate in rat liver endothelial cells. Biochem. J. 1984, 223, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Smedsrod, B. Cellular Events in the Uptake and Degradation of Hyaluronan. Adv. Drug Deliv. Rev. 1991, 7, 265–278. [Google Scholar] [CrossRef]
- Laremore, T.N.; Zhang, F.; Dordick, J.S.; Liu, J.; Linhardt, R.J. Recent progress and applications in glycosaminoglycan and heparin research. Curr. Opin. Chem. Biol. 2009, 13, 633–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabenstein, D.L. Heparin and heparan sulfate: Structure and function. Nat. Prod. Rep. 2002, 19, 312–331. [Google Scholar] [CrossRef] [PubMed]
- Kwaan, H.C.; Hatem, A. Effect of lung and gut heparin on experimental arterial thrombosis. Adv. Exp. Med. Biol. 1975, 52, 263–268. [Google Scholar]
- Barrowcliffe, T.W. History of heparin. Handb. Exp. Pharmacol. 2012, 207, 3–22. [Google Scholar]
- Harris, E.N.; Baggenstoss, B.A.; Weigel, P.H. Rat and human HARE/stabilin-2 are clearance receptors for high- and low-molecular-weight heparins. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1191–G1199. [Google Scholar] [CrossRef] [Green Version]
- Praaning, D.P.P.-V.; Brouwer, A.; Knook, D.L. Clearance capacity of rat liver Kupffer, Endothelial, and parenchymal cells. Gastroenterology 1981, 81, 1036–1044. [Google Scholar]
- Oie, C.I.; Olsen, R.; Smedsrod, B.; Hansen, J.B. Liver sinusoidal endothelial cells are the principal site for elimination of unfractionated heparin from the circulation. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G520–G528. [Google Scholar] [CrossRef] [Green Version]
- Harris, E.N.; Weigel, J.A.; Weigel, P.H. The human hyaluronan receptor for endocytosis (HARE/Stabilin-2) is a systemic clearance receptor for heparin. J. Biol. Chem. 2008, 283, 17341–17350. [Google Scholar] [CrossRef] [PubMed]
- Pempe, E.H.; Xu, Y.; Gopalakrishnan, S.; Liu, J.; Harris, E.N. Probing structural selectivity of synthetic heparin binding to stabilin protein receptors. J. Biol. Chem. 2012, 287, 20774–20783. [Google Scholar] [CrossRef] [PubMed]
- Mulloy, B.; Heath, A.; Shriver, Z.; Jameison, F.; Al Hakim, A.; Morris, T.S.; Szajek, A.Y. USP compendial methods for analysis of heparin: Chromatographic determination of molecular weight distributions for heparin sodium. Anal. Bioanal. Chem. 2014, 406, 4815–4823. [Google Scholar] [CrossRef] [PubMed]
- Volpi, N.; Maccari, F.; Suwan, J.; Linhardt, R.J. Electrophoresis for the analysis of heparin purity and quality. Electrophoresis 2012, 33, 1531–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petitou, M. Chemical Synthesis of Heparin. In Heparin: Chemical and Biological Properties, Clinical Applications; Lane, D.A., Lindahl, U., Eds.; CRC Press: Boca Raton, FL, USA, 1989; pp. 65–79. [Google Scholar]
- Lindahl, U. Approaches to the synthesis of heparin. Haemostasis 1990, 20 (Suppl. 1), 146–153. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Xu, Y.; Chen, M.; Weïwer, M.; Zhou, X.; Bridges, A.S.; DeAngelis, P.L.; Zhang, Q.; Linhardt, R.J.; Liu, J. Chemoenzymatic design of heparan sulfate oligosaccharides. J. Biol. Chem. 2010, 285, 34240–34249. [Google Scholar] [CrossRef]
- Linhardt, R.J.; Liu, J. Synthetic heparin. Curr. Opin. Pharmacol. 2012, 12, 217–219. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Cai, C.; Chandarajoti, K.; Hsieh, P.H.; Li, L.; Pham, T.Q.; Sparkenbaugh, E.M.; Sheng, J.; Key, N.S.; Pawlinski, R.; et al. Homogeneous low-molecular-weight heparins with reversible anticoagulant activity. Nat. Chem. Biol. 2014, 10, 248–250. [Google Scholar] [CrossRef]
- Bhaskar, U.; Li, G.; Fu, L.; Onishi, A.; Suflita, M.; Dordick, J.S.; Linhardt, R.J. Combinatorial one-pot chemoenzymatic synthesis of heparin. Carbohydr. Polym. 2015, 122, 399–407. [Google Scholar] [CrossRef]
- Joice, A.; Raman, K.; Mencio, C.; Quintero, M.V.; Brown, S.; Nguyen, T.K.; Kuberan, B. Enzymatic synthesis of heparan sulfate and heparin. Methods Mol. Biol. 2015, 1229, 11–19. [Google Scholar]
- Lane, R.S.; Haller, F.M.; Chavaroche, A.A.E.; Almond, A.; DeAngelis, P.L. Heparosan-coated liposomes for drug delivery. Glycobiology 2017, 27, 1062–1074. [Google Scholar] [CrossRef] [PubMed]
- DeAngelis, P.L. Heparosan, a promising ‘naturally good’ polymeric conjugating vehicle for delivery of injectable therapeutics. Expert Opin. Drug Deliv. 2015, 12, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.M.; Xu, Y.; Kudrna, K.M.; Hass, B.E.; Kellar, B.M.; Egger, A.W.; Liu, J.; Harris, E.N. 3-O sulfation of heparin leads to hepatotropism and longer circulatory half-life. Thromb. Res. 2018, 167, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, D.J.; Mulloy, B.; Forster, M.J.; Blundell, C.D.; Fries, E.; Milner, C.M.; Day, A.J. Characterization of the interaction between tumor necrosis factor-stimulated gene-6 and heparin: Implications for the inhibition of plasmin in extracellular matrix microenvironments. J. Biol. Chem. 2005, 280, 27044–27055. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.N.; Weigel, P.H. The ligand-binding profile of HARE: Hyaluronan and chondroitin sulfates A, C, and D bind to overlapping sites distinct from the sites for heparin, acetylated low-density lipoprotein, dermatan sulfate, and CS-E. Glycobiology 2008, 18, 638–648. [Google Scholar] [CrossRef]
- Crooke, S.T.; Witztum, J.L.; Bennett, C.F.; Baker, B.F. RNA-Targeted Therapeutics. Cell Metab. 2018, 27, 714–739. [Google Scholar] [CrossRef]
- Miller, C.M.; Harris, E.N. Antisense Oligonucleotides: Treatment Strategies and Cellular Internalization. RNA Dis. 2016, 3, 1393. [Google Scholar]
- Seth, P.P.; Tanowitz, M.; Bennett, C.F. Selective tissue targeting of synthetic nucleic acid drugs. J. Clin. Investig. 2019, 129, 915–925. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K.; Prakash, T.P.; Donner, A.J.; Kinberger, G.A.; Gaus, H.J.; Low, A.; Ostergaard, M.E.; Bell, M.; Swayze, E.E.; Seth, P.P. Characterizing the effect of GalNAc and phosphorothioate backbone on binding of antisense oligonucleotides to the asialoglycoprotein receptor. Nucleic Acids Res. 2017, 45, 2294–2306. [Google Scholar] [CrossRef]
- Ammala, C.; Drury, W.J., 3rd; Knerr, L.; Ahlstedt, I.; Stillemark-Billton, P.; Wennberg-Huldt, C.; Andersson, E.M.; Valeur, E.; Jansson-Lofmark, R.; Janzen, D.; et al. Targeted delivery of antisense oligonucleotides to pancreatic beta-cells. Sci. Adv. 2018, 4, eaat3386. [Google Scholar] [CrossRef]
- Geary, R.S.; Norris, D.; Yu, R.; Bennett, C.F. Pharmacokinetics, biodistribution and cell uptake of antisense oligonucleotides. Adv. Drug Deliv. Rev. 2015, 87, 46–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanowitz, M.; Hettrick, L.; Revenko, A.; Kinberger, G.A.; Prakash, T.P.; Seth, P.P. Asialoglycoprotein receptor 1 mediates productive uptake of N-acetylgalactosamine-conjugated and unconjugated phosphorothioate antisense oligonucleotides into liver hepatocytes. Nucleic Acids Res. 2017, 45, 12388–12400. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.; Crooke, R.M.; Graham, M.J.; Lemonidis, K.M.; Lougheed, M.; Murray, S.F.; Witchell, D.; Steinbrecher, U.; Bennett, C.F. Phosphorothioate oligodeoxynucleotides distribute similarly in class A scavenger receptor knockout and wild-type mice. J. Pharmacol. Exp. Ther. 2000, 292, 489–496. [Google Scholar] [PubMed]
- Wang, S.; Allen, N.; Vickers, T.A.; Revenko, A.S.; Sun, H.; Liang, X.H.; Crooke, S.T. Cellular uptake mediated by epidermal growth factor receptor facilitates the intracellular activity of phosphorothioate-modified antisense oligonucleotides. Nucleic Acids Res. 2018, 46, 3579–3594. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.M.; Donner, A.J.; Blank, E.E.; Egger, A.W.; Kellar, B.M.; Ostergaard, M.E.; Seth, P.P.; Harris, E.N. Stabilin-1 and Stabilin-2 are specific receptors for the cellular internalization of phosphorothioate-modified antisense oligonucleotides (ASOs) in the liver. Nucleic Acids Res. 2016, 44, 2782–2794. [Google Scholar] [CrossRef] [PubMed]
- Gaus, H.; Miller, C.M.; Seth, P.P.; Harris, E.N. Structural Determinants for the Interactions of Chemically Modified Nucleic Acids with the Stabilin-2 Clearance Receptor. Biochemistry 2018, 57, 2061–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirie, E.; Ray, S.; Pan, C.; Fu, W.; Powers, A.F.; Polikoff, D.; Miller, C.M.; Kudrna, K.M.; Harris, E.N.; Lusis, A.J.; et al. Mouse genome-wide association studies and systems genetics uncover the genetic architecture associated with hepatic pharmacokinetic and pharmacodynamic properties of a constrained ethyl antisense oligonucleotide targeting Malat1. PLoS Genet. 2018, 14, e1007732. [Google Scholar] [CrossRef]
- Miller, C.M.; Wan, W.B.; Seth, P.P.; Harris, E.N. Endosomal Escape of Antisense Oligonucleotides Internalized by Stabilin Receptors Is Regulated by Rab5C and EEA1 During Endosomal Maturation. Nucleic Acids Ther. 2018, 28, 86–96. [Google Scholar] [CrossRef]
- Linnane, E.; Davey, P.; Zhang, P.; Puri, S.; Edbrooke, M.; Chiarparin, E.; Revenko, A.S.; Macleod, A.R.; Norman, J.C.; Ross, S.J. Differential uptake, kinetics and mechanisms of intracellular trafficking of next-generation antisense oligonucleotides across human cancer cell lines. Nucleic Acids Res. 2019, 47, 4375–4392. [Google Scholar] [CrossRef]
- Park, S.Y.; Kang, K.B.; Thapa, N.; Kim, S.Y.; Lee, S.J.; Kim, I.S. Requirement of adaptor protein GULP during stabilin-2-mediated cell corpse engulfment. J. Biol. Chem. 2008, 283, 10593–10600. [Google Scholar] [CrossRef]
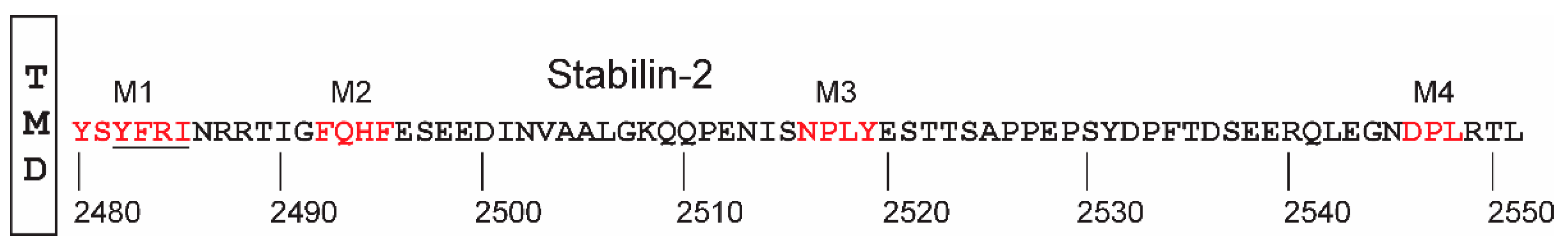
- Pandey, M.S.; Harris, E.N.; Weigel, J.A.; Weigel, P.H. The cytoplasmic domain of the hyaluronan receptor for endocytosis (HARE) contains multiple endocytic motifs targeting coated pit-mediated internalization. J. Biol. Chem. 2008, 283, 21453–21461. [Google Scholar] [CrossRef] [PubMed]
- Handel, T.M.; Johnson, Z.; Crown, S.E.; Lau, E.K.; Proudfoot, A.E. Regulation of protein function by glycosaminoglycans—As exemplified by chemokines. Annu. Rev. Biochem. 2005, 74, 385–410. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.S.; Baggenstoss, B.A.; Washburn, J.; Harris, E.N.; Weigel, P.H. The hyaluronan receptor for endocytosis (HARE) activates NF-kappaB-mediated gene expression in response to 40-400-kDa, but not smaller or larger, hyaluronans. J. Biol. Chem. 2013, 288, 14068–14079. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.N.; Parry, S.; Sutton-Smith, M.; Pandey, M.S.; Panico, M.; Morris, H.R.; Haslam, S.M.; Dell, A.; Weigel, P.H. N-Glycans on the link domain of human HARE/Stabilin-2 are needed for hyaluronan binding to purified ecto-domain, but not for cellular endocytosis of hyaluronan. Glycobiology 2010, 20, 991–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, M.S.; Weigel, P.H. Hyaluronic Acid Receptor for Endocytosis (HARE)-Mediated Endocytosis of Hyaluronan, Heparin, Dermatan Sulfate and Acetylated LDL, but not Chondroitin Sulfate Types A, C, D or E Activates NF-kB-Regulated Gene Expression. J. Biol. Chem. 2014, 289, 1756–1767. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.S.; Miller, C.M.; Harris, E.N.; Weigel, P.H. Activation of ERK and NF-kappaB during HARE-Mediated Heparin Uptake Require Only One of the Four Endocytic Motifs. PLoS ONE 2016, 11, e0154124. [Google Scholar] [CrossRef] [PubMed]
- Weigel, P.H.; Baggenstoss, B.A. What is special about 200 kDa hyaluronan that activates hyaluronan receptor signaling? Glycobiology 2017, 27, 868–877. [Google Scholar] [CrossRef] [Green Version]
- Hammoutene, A.; Rautou, P.E. Role of liver sinusoidal endothelial cells in non-alcoholic fatty liver disease. J. Hepatol. 2019, 70, 1278–1291. [Google Scholar] [CrossRef] [Green Version]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harris, E.N.; Cabral, F. Ligand Binding and Signaling of HARE/Stabilin-2. Biomolecules 2019, 9, 273. https://doi.org/10.3390/biom9070273
Harris EN, Cabral F. Ligand Binding and Signaling of HARE/Stabilin-2. Biomolecules. 2019; 9(7):273. https://doi.org/10.3390/biom9070273
Chicago/Turabian StyleHarris, Edward N., and Fatima Cabral. 2019. "Ligand Binding and Signaling of HARE/Stabilin-2" Biomolecules 9, no. 7: 273. https://doi.org/10.3390/biom9070273
APA StyleHarris, E. N., & Cabral, F. (2019). Ligand Binding and Signaling of HARE/Stabilin-2. Biomolecules, 9(7), 273. https://doi.org/10.3390/biom9070273