New Monoclonal Antibodies for a Selective Detection of Membrane-Associated and Soluble Forms of Carbonic Anhydrase IX in Human Cell Lines and Biological Samples

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Recombinant Carbonic Anhydrases
2.3. Blood and Tissue Samples
2.4. Generation of Monoclonal Antibodies against Human CA IX Protein
2.5. Enzyme-Linked Immunosorbent Assay
2.5.1. Indirect ELISA
2.5.2. Direct ELISA
2.5.3. Competitive ELISA
2.5.4. Sandwich ELISA
2.6. Isotype Determination of the MAbs
2.7. Epitope Mapping with Overlapping Biotinylated Peptides
2.8. Purification of the MAbs from Hybridoma Supernatants
2.9. Conjugation of the MAbs to Horseradish Peroxidase
2.10. Determination of the Apparent Dissociation Constant
2.11. Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis
2.12. Western Blot
2.13. Flow Cytometry Analysis
2.14. Immunoprecipitation
2.15. Immunofluorescence Staining
2.16. Near-Infrared Fluorescence Western Blot
2.17. Immunohistochemistry
3. Results
3.1. Generation of MAbs against CA IX
3.2. Characterization of MAbs by Various Immunoassays
3.3. Epitope Analysis by a Competitive ELISA and an Indirect ELISA Using Overlapping Peptides
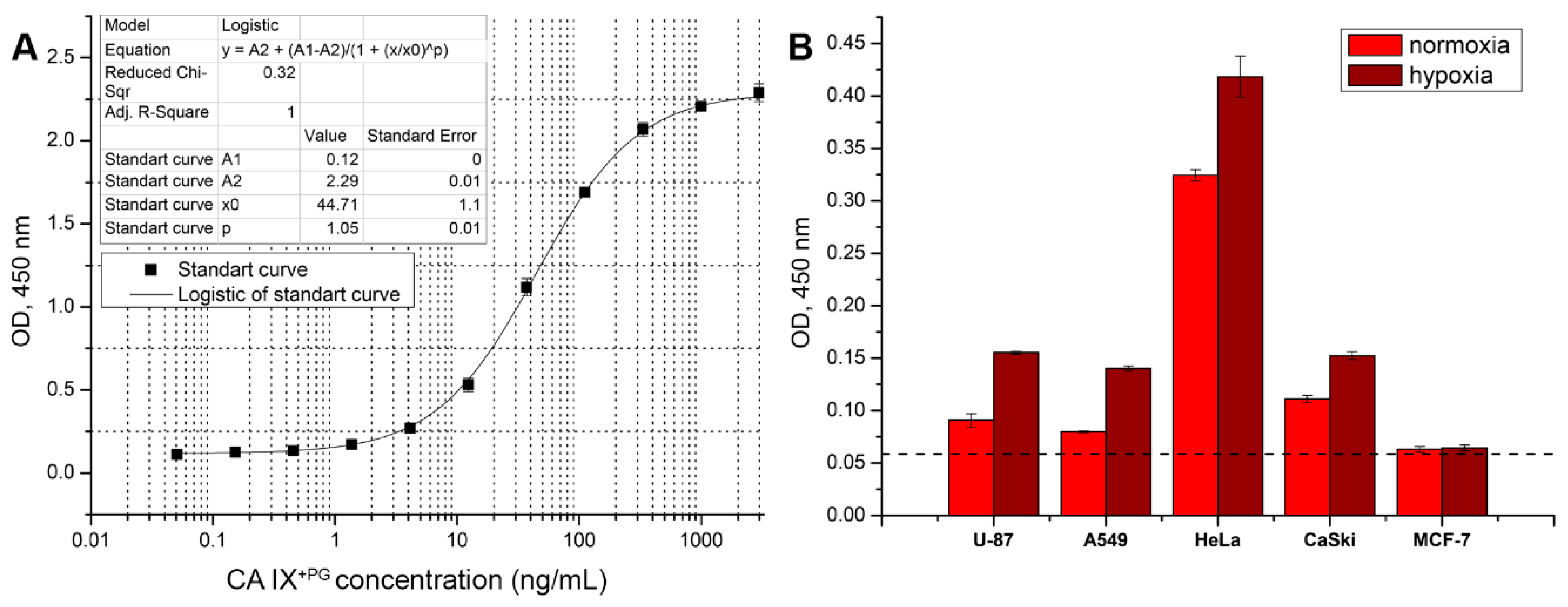
3.4. Application of the MAbs for Studying CA IX Expression in Human Tumor Cells
3.5. Application of the MAbs for the Visualization of CA IX-Positive Tumor Cells by Immunofluorescence
3.6. Development of Sandwich ELISA for Detection of Soluble CA IX
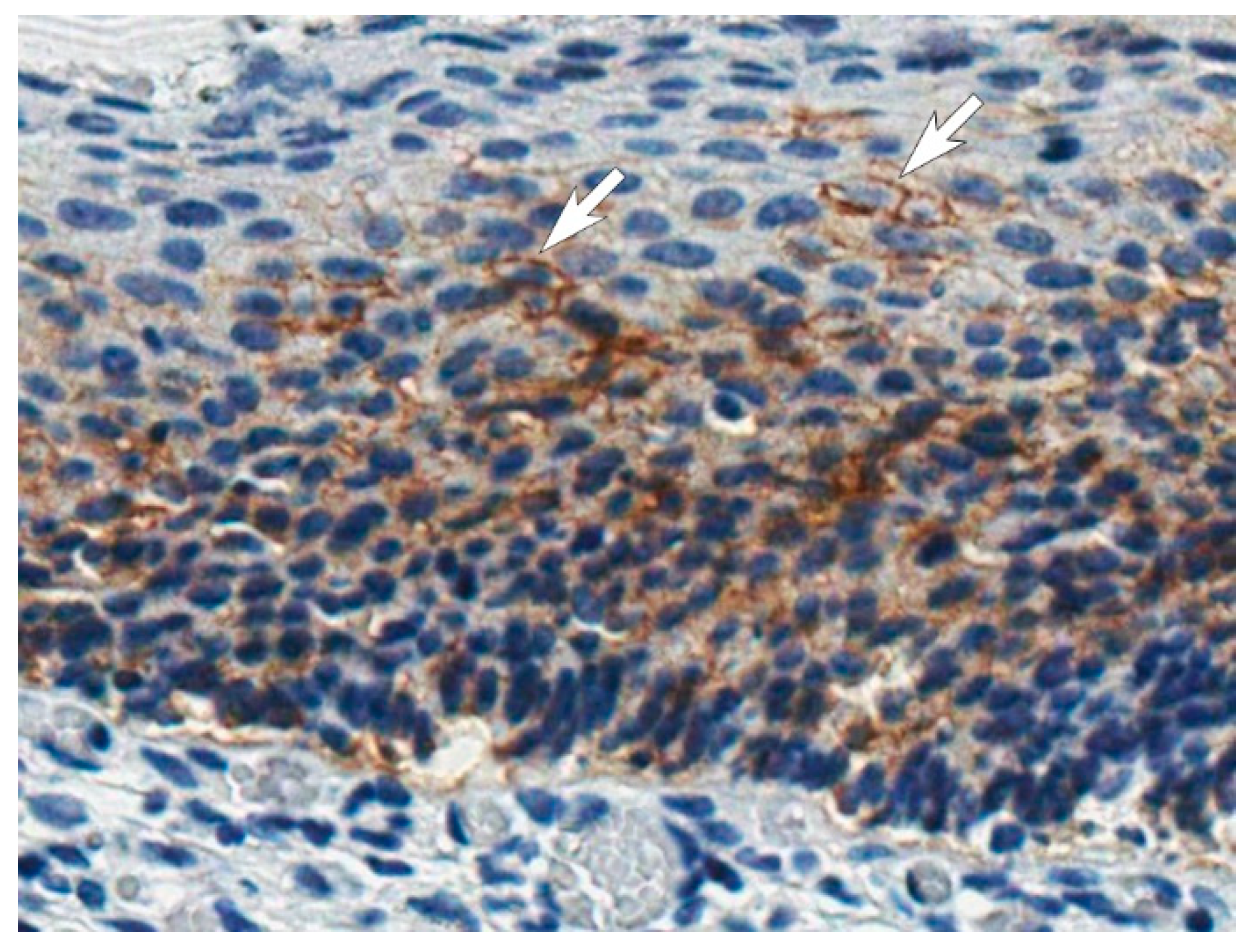
3.7. Application of the MAbs for CA IX Detection in Tissues by the Immunohistochemistry Assay
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| A431 | human epidermoid carcinoma cell line |
| A498 | human renal cell carcinoma cell line |
| A549 | human lung carcinoma cell line |
| aa | amino acid |
| BSA | bovine serum albumin |
| CA | carbonic anhydrase |
| CA IX | carbonic anhydrase IX |
| CA IX+PG | recombinant full-length extracellular domain of CA IX |
| CA IX-PG | recombinant catalytic domain without PG region |
| CA XII | carbonic anhydrase XII |
| CaSki | human cervical squamous cell carcinoma cell line |
| DMEM | Dulbecco’s modified Eagle’s growth medium |
| ELISA | enzyme-linked immunosorbent assay |
| FBS | fetal bovine serum |
| HAT | hypoxanthine/aminopterin/thymidine |
| HEK-293 | human embryonic kidney 293 cell line |
| HeLa | human cervical adenocarcinoma cell line |
| HRP | horseradish peroxidase |
| HT29 | human colorectal carcinoma cell line |
| IFA | immunofluorescence assay |
| IgG | immunoglobulin G |
| IHC | immunohistochemistry |
| IP | immunoprecipitation |
| Jurkat | human T cell leukemia cell line |
| Kd | apparent dissociation constant |
| MAbs | monoclonal antibodies |
| MCF-7 | human breast adenocarcinoma cell line |
| MDA-MB-231 | human triple-negative breast cancer cell line |
| OD | optical density |
| PBS | phosphate-buffered saline |
| PBS-T | 0.1% Tween 20 in PBS buffer |
| PBS-TW | PBS supplemented with 0.05% of Tween 20 |
| PG | proteoglycan region of CA IX |
| PPV | porcine parvovirus |
| PVDF | polyvinylidene difluoride |
| RT | room temperature |
| SDS | sodium dodecyl sulfate |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| TBS-T | Tris buffered saline with 0.5% Tween 20 |
| TMB | 3,3′,5,5′-Tetramethylbenzidine |
| U-87 | human glioblastoma cell line |
| WB | Western blot |
References
- Stratton, M.R.; Campbell, P.J.; Futreal, P.A. The cancer genome. Nature 2009, 458, 719–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertram, J.S. The molecular biology of cancer. Mol. Asp. Med. 2000, 21, 167–223. [Google Scholar] [CrossRef]
- Bhatt, A.N.; Mathur, R.; Farooque, A.; Verma, A.; Dwarakanath, B.S. Cancer biomarkers—Current perspectives. Indian J. Med. Res. 2010, 132, 129–149. [Google Scholar] [PubMed]
- Duffy, M.J. Role of tumor markers in patients with solid cancers: A critical review. Eur. J. Intern. Med. 2007, 18, 175–184. [Google Scholar] [CrossRef]
- Karley, D.; Gupta, D.; Tiwari, A. Biomarker for Cancer: A great Promise for Future. World J. Oncol. 2011, 2, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Soori, G.; Dobleman, T.J.; Xiao, G.G. The application of monoclonal antibodies in cancer diagnosis. Expert Rev. Mol. Diagn. 2014, 14, 97–106. [Google Scholar] [CrossRef]
- Ventola, C.L. Cancer Immunotherapy, Part 3: Challenges and Future Trends. P&T 2017, 42, 514–521. [Google Scholar]
- Chatziharalambous, D.; Lygirou, V.; Latosinska, A.; Stravodimos, K.; Vlahou, A.; Jankowski, V.; Zoidakis, J. Analytical performance of ELISA assays in Urine: One more bottleneck towards biomarker validation and clinical implementation. PLoS ONE 2016, 11, e0149471. [Google Scholar] [CrossRef]
- Warram, J.M.; de Boer, E.; Sorace, A.G.; Chung, T.K.; Kim, H.; Pleijhuis, R.G.; van Dam, G.M.; Rosenthal, E.L. Antibody-based imaging strategies for cancer. Cancer Metastasis Rev. 2014, 33, 809–822. [Google Scholar] [CrossRef] [Green Version]
- De Boer, E.; Warram, J.M.; Tucker, M.D.; Hartman, Y.E.; Moore, L.S.; De Jong, J.S.; Chung, T.K.; Korb, M.L.; Zinn, K.R.; Van Dam, G.M.; et al. In Vivo Fluorescence Immunohistochemistry: Localization of Fluorescently Labeled Cetuximab in Squamous Cell Carcinomas. Sci. Rep. 2015, 5, 10169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventola, C.L. Cancer Immunotherapy, Part 1: Current Strategies and Agents. P&T 2017, 42, 375–383. [Google Scholar]
- Thiry, A.; Dogné, J.M.; Masereel, B.; Supuran, C.T. Targeting tumor-associated carbonic anhydrase IX in cancer therapy. Trends Pharmacol. Sci. 2006, 27, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Mahon, B.P.; Pinard, M.A.; McKenna, R. Targeting carbonic anhydrase IX activity and expression. Molecules 2015, 20, 2323–2348. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.M.; Allison, J.P.; Wolchok, J.D. Monoclonal antibodies in cancer therapy. Cancer Immun. 2012, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Waheed, A.; Sly, W.S. Carbonic anhydrase XII functions in health and disease. Gene 2017, 623, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Leppilampi, M.; Saarnio, J.; Karttunen, T.J.; Kivelä, J.; Pastoreková, S.; Pastorek, J.; Waheed, A.; Sly, W.S.; Parkkila, S. Carbonic anhydrase isozymes IX and XII in gastric tumors. World J. Gastroenterol. 2003, 9, 1398–1403. [Google Scholar] [CrossRef]
- Pastorek, J.; Pastorekova, S. Hypoxia-induced carbonic anhydrase IX as a target for cancer therapy: From biology to clinical use. Semin. Cancer Biol. 2015, 31, 52–64. [Google Scholar] [CrossRef]
- Lau, J.; Lin, K.-S.; Bénard, F. Past, Present, and Future: Development of Theranostic Agents Targeting Carbonic Anhydrase IX. Theranostics 2017, 7, 4322–4339. [Google Scholar] [CrossRef]
- Stillebroer, A.B.; Mulders, P.F.A.; Boerman, O.C.; Oyen, W.J.G.; Oosterwijk, E. Carbonic anhydrase IX in renal cell carcinoma: Implications for prognosis, diagnosis, and therapy. Eur. Urol. 2010, 58, 75–83. [Google Scholar] [CrossRef]
- Oosterwijk, E.; Ruiter, D.J.; Hoedemaeker, P.J.; Pauwels, E.K.; Jonas, U.; Zwartendijk, J.; Warnaar, S.O. Monoclonal antibody G 250 recognizes a determinant present in renal-cell carcinoma and absent from normal kidney. Int. J. Cancer 1986, 38, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Pastorekova, S.; Zavadova, Z.; Kostal, M.; Babusikova, O.; Zavada, J. A novel quasi-viral agent, MaTu, is a two-component system. Virology 1992, 187, 620–626. [Google Scholar] [CrossRef]
- Opavský, R.; Pastoreková, S.; Zelník, V.; Gibadulinová, A.; Stanbridge, E.J.; Závada, J.; Kettmann, R.; Pastorek, J. Human MN/CA9 gene, a novel member of the carbonic anhydrase family: Structure and exon to protein domain relationships. Genomics 1996, 33, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Hewett-Emmett, D.; Tashian, R.E. Functional Diversity, Conservation, and Convergence in the Evolution of the α-, β-, and γ-Carbonic Anhydrase Gene Families. Mol. Phylogenet. Evol. 1996, 5, 50–77. [Google Scholar] [CrossRef]
- Grabmaier, K.; Vissers, J.L.M.; De Weijert, M.C.A.; Oosterwijk-Wakka, J.C.; Van Bokhoven, A.; Brakenhoff, R.H.; Noessner, E.; Mulders, P.A.; Merkx, G.; Figdor, C.G.; et al. Molecular cloning and immunogenicity of renal cell carcinoma-associated antigen G250. Int. J. Cancer 2000, 85, 865–870. [Google Scholar] [CrossRef]
- Oosterwijk, E. Carbonic anhydrase IX: Historical and future perspectives. BJU Int. 2008, 101, 2–7. [Google Scholar] [CrossRef]
- Surfus, J.E.; Hank, J.A.; Oosterwijk, E.; Welt, S.; Lindstrom, M.J.; Albertini, M.R.; Schiller, J.H.; Sondel, P.M. Anti-Renal-Cell Carcinoma Chimeric Antibody G250 Facilitates Antibody-Dependent Cellular Cytotoxicity with In Vitro and In Vivo Interleukin-2-Activated Effectors. J. Immunother. 1996, 19, 184–191. [Google Scholar] [CrossRef] [Green Version]
- McDonald, P.C.; Winum, J.-Y.; Supuran, C.T.; Dedhar, S. Recent developments in targeting carbonic anhydrase IX for cancer therapeutics. Oncotarget 2012, 3, 84–97. [Google Scholar] [CrossRef]
- Chia, S.K.; Wykoff, C.C.; Watson, P.H.; Han, C.; Leek, R.D.; Pastorek, J.; Gatter, K.C.; Ratcliffe, P.; Harris, A.L. Prognostic significance of a novel hypoxia-regulated marker, carbonic anhydrase IX, in invasive breast carcinoma. J. Clin. Oncol. 2001, 19, 3660–3668. [Google Scholar] [CrossRef]
- Proescholdt, M.A.; Mayer, C.; Kubitza, M.; Schubert, T.; Liao, S.-Y.; Stanbridge, E.J.; Ivanov, S.; Oldfield, E.H.; Brawanski, A.; Merrill, M.J. Expression of hypoxia-inducible carbonic anhydrases in brain tumors. Neuro. Oncol. 2005, 7, 465–475. [Google Scholar] [CrossRef]
- Závada, J.; Závadová, Z.; Zat’ovičová, M.; Hyršl, L.; Kawaciuk, I.; Zat’ovicová, M.; Hyrsl, L.; Kawaciuk, I. Soluble form of carbonic anhydrase IX (CA IX) in the serum and urine of renal carcinoma patients. Br. J. Cancer 2003, 89, 1067–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatovicova, M.; Sedlakova, O.; Svastova, E.; Ohradanova, A.; Ciampor, F.; Arribas, J.; Pastorek, J.; Pastorekova, S. Ectodomain shedding of the hypoxia-induced carbonic anhydrase IX is a metalloprotease-dependent process regulated by TACE/ADAM17. Br. J. Cancer 2005, 93, 1267–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murri-Plesko, M.T.; Hulikova, A.; Oosterwijk, E.; Scott, A.M.; Zortea, A.; Harris, A.L.; Ritter, G.; Old, L.; Bauer, S.; Swietach, P.; et al. Antibody inhibiting enzymatic activity of tumour-associated carbonic anhydrase isoform IX. Eur. J. Pharmacol. 2011, 657, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Ahlskog, J.K.J.; Schliemann, C.; Mårlind, J.; Qureshi, U.; Ammar, A.; Pedley, R.B.; Neri, D. Human monoclonal antibodies targeting carbonic anhydrase IX for the molecular imaging of hypoxic regions in solid tumours. Br. J. Cancer 2009, 101, 645–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Lo, A.; Yammanuru, A.; Tallarico, A.S.C.; Brady, K.; Murakami, A.; Barteneva, N.; Zhu, Q.; Marasco, W.A. Unique biological properties of catalytic domain directed human anti-CAIX antibodies discovered through phage-display technology. PLoS ONE 2010, 5, e9625. [Google Scholar] [CrossRef] [PubMed]
- Dudutienė, V.; Matulienė, J.; Smirnov, A.; Timm, D.D.; Zubrienė, A.; Baranauskienė, L.; Morkūnaitė, V.; Smirnovienė, J.; Michailovienė, V.; Juozapaitienė, V.; et al. Discovery and Characterization of Novel Selective Inhibitors of Carbonic Anhydrase IX. J. Med. Chem. 2014, 57, 9435–9446. [Google Scholar] [CrossRef] [PubMed]
- Linkuvienė, V.; Matulienė, J.; Juozapaitienė, V.; Michailovienė, V.; Jachno, J.; Matulis, D. Intrinsic thermodynamics of inhibitor binding to human carbonic anhydrase IX. Biochim. Biophys. Acta Gen. Subj. 2016, 1860, 708–718. [Google Scholar] [CrossRef]
- Dekaminaviciute, D.; Lasickiene, R.; Parkkila, S.; Jogaite, V.; Matuliene, J.; Matulis, D.; Zvirbliene, A. Development and characterization of new monoclonal antibodies against human recombinant CA XII. BioMed Res. Int. 2014, 2014, 309307. [Google Scholar] [CrossRef]
- Köhler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef]
- Harlow, E.; Lane, D. Antibodies: A Laboratory Manual, 1st ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1988. [Google Scholar]
- Armbruster, D.A.; Pry, T. Limit of Blank, Limit of Detection and Limit of Quantitation. Clin. Biochem. Rev. 2008, 29, S49–S52. [Google Scholar]
- Tamošiūnas, P.L.; Petraitytė-Burneikienė, R.; Lasickienė, R.; Akatov, A.; Kundrotas, G.; Sereika, V.; Lelešius, R.; Žvirblienė, A.; Sasnauskas, K. Generation of recombinant porcine parvovirus virus-like particles in Saccharomyces cerevisiae and development of virus-specific monoclonal antibodies. J. Immunol. Res. 2014, 2014, 573531. [Google Scholar] [CrossRef] [PubMed]
- Závada, J.; Závadová, Z.; Pastorek, J.; Biesová, Z.; Jezek, J.; Velek, J. Human tumour-associated cell adhesion protein MN/CA IX: Identification of M75 epitope and of the region mediating cell adhesion. Br. J. Cancer 2000, 82, 1808–1813. [Google Scholar] [CrossRef] [PubMed]
- Swinson, D.E.B.; Jones, J.L.; Richardson, D.; Wykoff, C.; Turley, H.; Pastorek, J.; Taub, N.; Harris, A.L.; O’Byrne, K.J. Carbonic anhydrase IX expression, a novel surrogate marker of tumor hypoxia, is associated with a poor prognosis in non-small-cell lung cancer. J. Clin. Oncol. 2003, 21, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.K.H. The history of monoclonal antibody development—Progress, remaining challenges and future innovations. Ann. Med. Surg. 2014, 3, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.G. Quality Issues of Research Antibodies. Anal. Chem. Insights 2016, 11, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Ecker, D.M.; Jones, S.D.; Levine, H.L. The therapeutic monoclonal antibody market. MAbs 2015, 7, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Bordeaux, J.; Welsh, A.; Agarwal, S.; Killiam, E.; Baquero, M.; Hanna, J.; Anagnostou, V.; Rimm, D. Antibody validation. Biotechniques 2010, 48, 197–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Lomelino, C.; Mboge, M.; Frost, S.; McKenna, R. Cancer Drug Development of Carbonic Anhydrase Inhibitors beyond the Active Site. Molecules 2018, 23, 1045. [Google Scholar] [CrossRef]
- Zatovicova, M.; Jelenska, L.; Hulikova, A.; Ditte, P.; Ditte, Z.; Csaderova, L.; Svastova, E.; Schmalix, W.; Boettger, V.; Bevan, P.; et al. Monoclonal antibody G250 targeting CA Ⅸ: Binding specificity, internalization and therapeutic effects in a non-renal cancer model. Int. J. Oncol. 2014, 45, 2455–2467. [Google Scholar] [CrossRef]
- Vidlickova, I.; Dequiedt, F.; Jelenska, L.; Sedlakova, O.; Pastorek, M.; Stuchlik, S.; Pastorek, J.; Zatovicova, M.; Pastorekova, S. Apoptosis-induced ectodomain shedding of hypoxia-regulated carbonic anhydrase IX from tumor cells: A double-edged response to chemotherapy. BMC Cancer 2016, 16, 239. [Google Scholar] [CrossRef]
- Leppilampi, M.; Koistinen, P.; Savolainen, E.-R.; Hannuksela, J.; Parkkila, A.-K.; Niemelä, O.; Pastoreková, S.; Pastorek, J.; Waheed, A.; Sly, W.S.; et al. The expression of carbonic anhydrase II in hematological malignancies. Clin. Cancer Res. 2002, 8, 2240–2245. [Google Scholar] [PubMed]
- Ivanova, L.; Zandberga, E.; Siliņa, K.; Kalniņa, Z.; Ābols, A.; Endzeliņš, E.; Vendina, I.; Romanchikova, N.; Hegmane, A.; Trapencieris, P.; et al. Prognostic relevance of carbonic anhydrase IX expression is distinct in various subtypes of breast cancer and its silencing suppresses self-renewal capacity of breast cancer cells. Cancer Chemother. Pharmacol. 2015, 75, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Kowalewska, M.; Radziszewski, J.; Kulik, J.; Barathova, M.; Nasierowska-Guttmajer, A.; Bidziński, M.; Pastorek, J.; Pastorekova, S.; Siedlecki, J.A. Detection of carbonic anhydrase 9-expressing tumor cells in the lymph nodes of vulvar carcinoma patients by RT-PCR. Int. J. Cancer 2005, 116, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.-J.; Rho, S.B.; Jung, D.C.; Han, I.-O.; Oh, E.-S.; Kim, J.-Y. Carbonic anhydrase IX (CA9) modulates tumor-associated cell migration and invasion. J. Cell Sci. 2011, 124, 1077–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Said, H.M.; Staab, A.; Hagemann, C.; Vince, G.H.; Katzer, A.; Flentje, M.; Vordermark, D. Distinct patterns of hypoxic expression of carbonic anhydrase IX (CA IX) in human malignant glioma cell lines. J. Neurooncol. 2006, 81, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Takacova, M.; Hlouskova, G.; Zatovicova, M.; Benej, M.; Sedlakova, O.; Kopacek, J.; Pastorek, J.; Lacik, I.; Pastorekova, S. Encapsulation of anti-carbonic anhydrase IX antibody in hydrogel microspheres for tumor targeting. J. Enzym. Inhib. Med. Chem. 2016, 31, 110–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide No. | Peptide Sequence | Peptide No. | Peptide Sequence |
|---|---|---|---|
| #1 | Biotin-SGSGLVPVHPQRLPRMQED-OH | #16 | Biotin-SGSGLGGGSSGEDD-OH |
| #2 | Biotin-SGSGPQRLPRMQEDSPLGG-OH | #17 | Biotin-SGSGGGSSGEDDPL-OH |
| #3 | Biotin-SGSGRMQEDSPLGGGSSGE-OH | #18 | Biotin-SGSGSSGEDDPLGE-OH |
| #4 | Biotin-SGSGSPLGGGSSGEDDPLG-OH | #19 | Biotin-SGSGGEDDPLGEED-OH |
| #5 | Biotin-SGSGGSSGEDDPLGEED-OH | #20 | Biotin-SGSGDDPLGEEDLP-OH |
| #6 | Biotin-SGSGDDPLGEEDLPSEEDS-OH | #21 | Biotin-SGSGPLGEEDLPSE-OH |
| #7 | Biotin-SGSGSEEDLPSEEDSPREED-OH | #22 | Biotin-SGSGGEEDLPSEED-OH |
| #8 | Biotin-SGSGSEEDSPREEDPPGEE-OH | #23 | Biotin-SGSGEDLPSEEDSP-OH |
| #9 | Biotin-SGSGPREEDPPGEEDLPGE-OH | #24 | Biotin-SGSGLPSEEDSPRE-OH |
| #10 | Biotin-SGSGPPGEEDLPGEEDLPG-OH | #25 | Biotin-SGSGDDPLGEED-OH |
| #11 | Biotin-SGSGDLPGEEDLPGEED-OH | #26 | Biotin-SGSGDDPLGEE-OH |
| #12 | Biotin-SGSGEDLPGEEDLPEVKPK-OH | #27 | Biotin-SGSGDDPLGE-OH |
| #13 | Biotin-SGSGSEEDLPEVKPKSEEEG-OH | #28 | Biotin-SGSGGEDDPLG-OH |
| #14 | Biotin-SGSGEVKPKSEEEGSLKLED-OH | ||
| #15 | Biotin-SGSGSEEEGSLKLEDLPTV-OH. |
| MAb Clone | IgG Subtype | Apparent Kd Value, M | MAbs Reactivity with CA IX 1 | |||||
|---|---|---|---|---|---|---|---|---|
| ELISA CA IX−PG | ELISA CA IX+PG | WB CA IX−PG | WB CA IX+PG | FC 2 | IFA 3 | |||
| A3 | IgG1 | 1.31 × 10−10 | + | + | − | − | + | + |
| F12 | 5.12 × 10−10 | + | + | − | − | + | + | |
| F8 | 3.29 × 10−10 | + | + | − | − | + | + | |
| F7 | 1.42 × 10−10 | + | + | − | − | + | + | |
| F4 | 6.5 × 10−10 | + | + | − | − | + | + | |
| D8 | 6.17 × 10−10 | + | + | − | − | + | + | |
| C9 | 2.51 × 10−10 | + | + | − | − | + | + | |
| G8 | 2.14 × 10−10 | + | + | − | − | + | ± | |
| H7 | 8.85 × 10−10 | − | + | − | + | + | + | |
| E3 | 1.75 × 10−10 | ± | + | − | − | − | ± | |
| A10 | 1.75 × 10−10 | ± | + | − | − | − | + | |
| H11 | 1.71 × 10−10 | ± | + | − | − | − | ± | |
| D3 | 2.2 × 10−10 | ± | + | − | − | − | ± | |
| M75 | 9.77 × 10−10 | − | + | − | + | ND 4 | ND | |
| Cell Line | Origin | Flow Cytometry | IP/WB | ||
|---|---|---|---|---|---|
| Normoxia | Hypoxia | Normoxia | Hypoxia | ||
| A549 | Lung carcinoma | − | + | − | + |
| U-87 | Glioblastoma | + | + | + | + |
| A498 | Renal cell carcinoma | − | − | − | − |
| Jurkat | T cell leukemia | − | − | − | − |
| HeLa | Cervical adenocarcinoma | − | + | − | + |
| A431 | Epidermoid carcinoma | − | + | − | + |
| CaSki | Cervical squamous cell carcinoma | + | + | + | + |
| MCF-7 | Breast adenocarcinoma | − | + | − | + |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stravinskiene, D.; Imbrasaite, A.; Petrikaite, V.; Matulis, D.; Matuliene, J.; Zvirbliene, A. New Monoclonal Antibodies for a Selective Detection of Membrane-Associated and Soluble Forms of Carbonic Anhydrase IX in Human Cell Lines and Biological Samples. Biomolecules 2019, 9, 304. https://doi.org/10.3390/biom9080304
Stravinskiene D, Imbrasaite A, Petrikaite V, Matulis D, Matuliene J, Zvirbliene A. New Monoclonal Antibodies for a Selective Detection of Membrane-Associated and Soluble Forms of Carbonic Anhydrase IX in Human Cell Lines and Biological Samples. Biomolecules. 2019; 9(8):304. https://doi.org/10.3390/biom9080304
Chicago/Turabian StyleStravinskiene, Dovile, Aiste Imbrasaite, Vilma Petrikaite, Daumantas Matulis, Jurgita Matuliene, and Aurelija Zvirbliene. 2019. "New Monoclonal Antibodies for a Selective Detection of Membrane-Associated and Soluble Forms of Carbonic Anhydrase IX in Human Cell Lines and Biological Samples" Biomolecules 9, no. 8: 304. https://doi.org/10.3390/biom9080304
APA StyleStravinskiene, D., Imbrasaite, A., Petrikaite, V., Matulis, D., Matuliene, J., & Zvirbliene, A. (2019). New Monoclonal Antibodies for a Selective Detection of Membrane-Associated and Soluble Forms of Carbonic Anhydrase IX in Human Cell Lines and Biological Samples. Biomolecules, 9(8), 304. https://doi.org/10.3390/biom9080304