Genome-Wide Association Study of Metabolic Syndrome Reveals Primary Genetic Variants at CETP Locus in Indians

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Study Subjects
2.3. Phenotype Definition (MetS)
2.4. Genome-Wide Association Study
2.4.1. Discovery Phase
2.4.2. Replication Phase and Meta-Analysis
2.5. Statistical Power of the Study
2.6. Conditional and Haplotype Association Analysis
2.7. Imputation Analysis
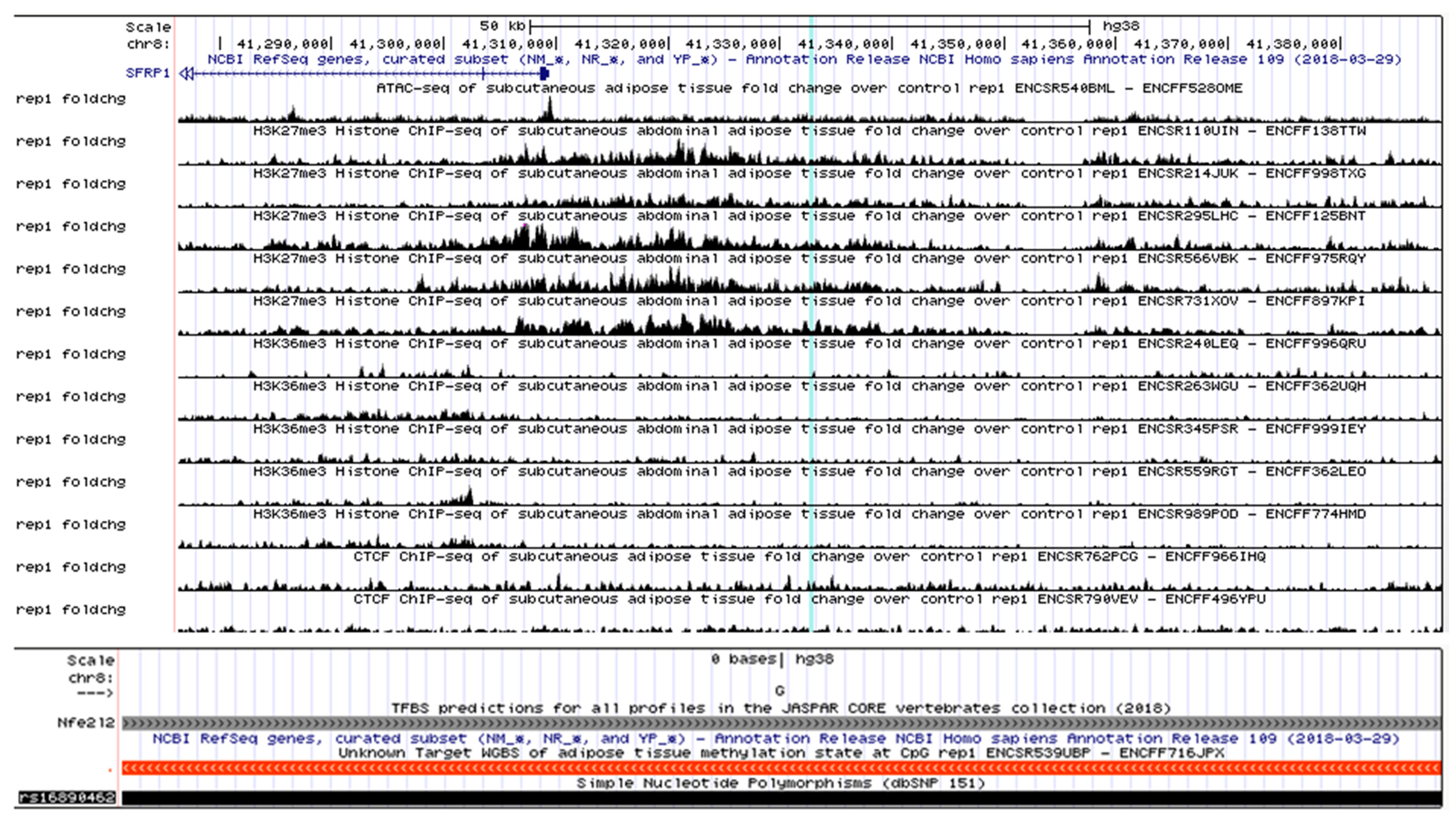
2.8. Overlaying Gene Regulatory Features
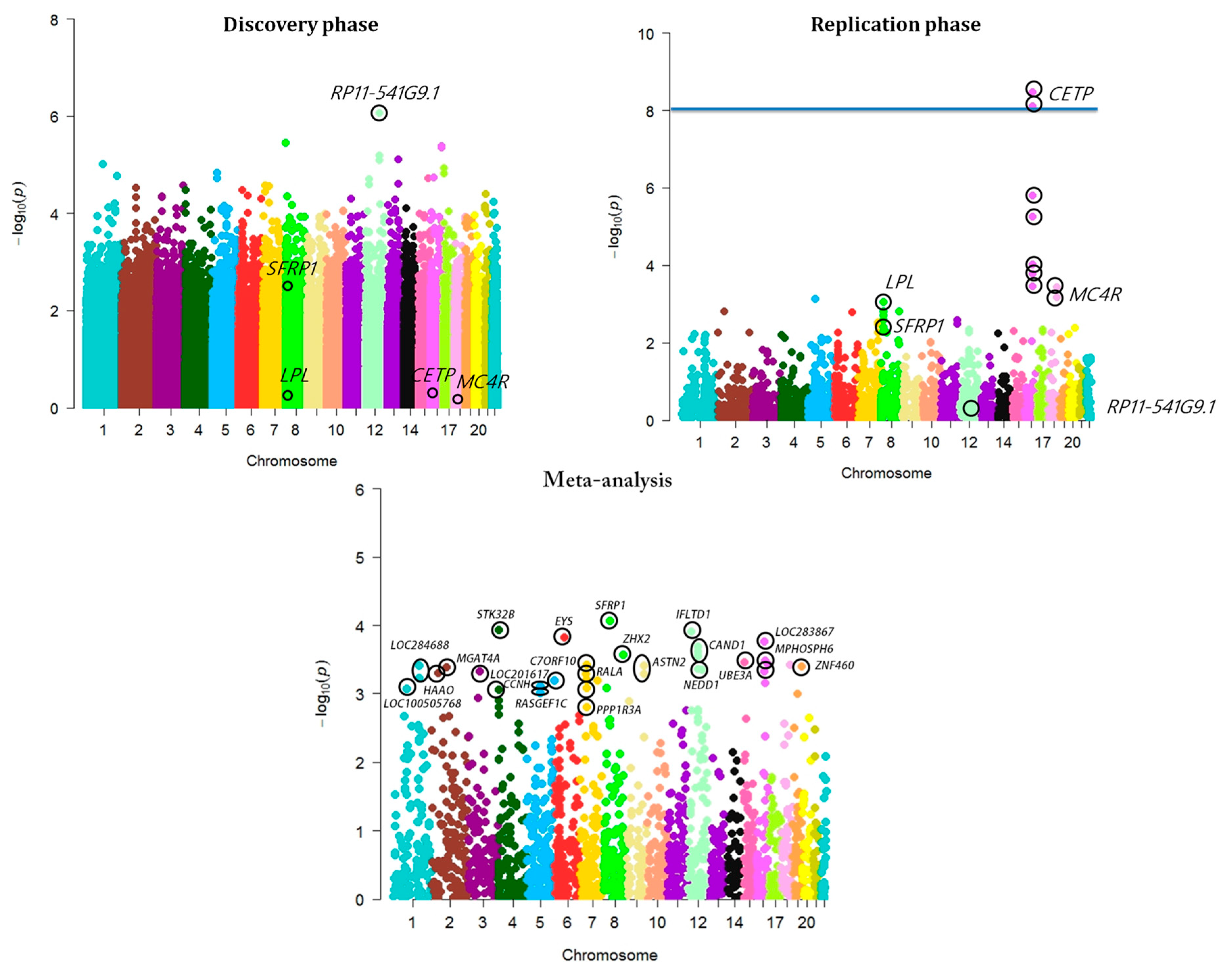
3. Results and Discussion
3.1. Genome-Wide Association Analysis of MetS
3.2. Conditional Analysis of CETP Locus
3.3. Haplotype Association Analysis
3.4. SFRP1—A Novel Sub-GWAS Locus for MetS in Indians
3.5. Imputation Analysis of Novel Locus
3.6. SFRP1, a Biologically Relevant Locus
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Members of INDICO
References
- Alberti, K.G.M.M.; Zimmet, P.; Shaw, J. Metabolic syndrome—A new world-wide definition. A consensus statement from the International Diabetes Federation. Diabet. Med. 2006, 23, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Mottillo, S.; Filion, K.B.; Genest, J.; Joseph, L.; Pilote, L.; Poirier, P.; Rinfret, S.; Schiffrin, E.L.; Eisenberg, M.J. The metabolic syndrome and cardiovascular risk: A systematic review and meta-analysis. J. Am. Coll. Cardiol. 2010, 56, 1113–1132. [Google Scholar] [CrossRef] [PubMed]
- Esposito, K.; Chiodini, P.; Colao, A.; Lenzi, A.; Giugliano, D. Metabolic syndrome and risk of cancer: A systematic review and meta-analysis. Diabetes Care. 2012, 35, 2402–2411. [Google Scholar] [CrossRef] [PubMed]
- Hui, W.S.; Liu, Z.; Ho, S.C. Metabolic syndrome and all-cause mortality: A meta-analysis of prospective cohort studies. Eur. J. Epidemiol. 2010, 25, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Misra, A.; Khurana, L. Obesity and the metabolic syndrome in developing countries. J. Clin. Endocrinol. Metab. 2008, 93, S9–S30. [Google Scholar] [CrossRef] [PubMed]
- Prasad, D.S.; Kabir, Z.; Dash, A.K.; Das, B.C. Prevalence and risk factors for metabolic syndrome in Asian Indians: A community study from urban Eastern India. J. Cardiovasc. Dis. Res. 2012, 3, 204–211. [Google Scholar] [CrossRef] [Green Version]
- Khan, Y.; Lalchandani, A.; Gupta, A.; Khadanga, S.; Kumar, S. Prevalence of metabolic syndrome crossing 40% in Northern India: Time to act fast before it runs out of proportions. J. Fam. Med. Prim. Care 2018, 7, 118–123. [Google Scholar] [CrossRef]
- Tandon, N.; Garg, M.K.; Singh, Y.; Marwaha, R.K. Prevalence of metabolic syndrome among urban Indian adolescents and its relation with insulin resistance (HOMA-IR). J. Pediatr. Endocrinol. Metab. 2013, 26, 1123–1130. [Google Scholar] [CrossRef]
- Joy, T.; Hegele, R.A. Genetics of Metabolic syndrome: Is there a role for phenomics? Curr. Atheroscler. Rep. 2008, 10, 201–208. [Google Scholar] [CrossRef]
- Henneman, P.; Aulchenko, Y.S.; Frants, R.R.; Zorkoltseva, I.V.; Zillikens, M.C.; Frolich, M.; Oostra, B.A.; Van Dijk, K.W.; Van Duijn, C.M. Genetic architecture of plasma adiponectin overlaps with the genetics of metabolic syndrome-related traits. Diabetes Care 2010, 33, 908–913. [Google Scholar] [CrossRef]
- Terán-García, M.; Bouchard, C. Genetics of the metabolic syndrome. Appl. Physiol. Nutr. Metab. 2007, 32, 89–114. [Google Scholar] [CrossRef]
- Watanabe, R.M.; Valle, T.; Hauser, E.R.; Ghosh, S.; Eriksson, J.; Kohtamaki, K.; Ehnholm, C.; Tuomilehto, J.; Collins, F.S.; Bergman, R.N.; et al. Familiality of quantitative metabolic traits in Finnish families with non-insulin-dependent diabetes mellitus. Finland-United States Investigation of NIDDM Genetics (FUSION) Study investigators. Hum. Hered. 1999, 49, 159–168. [Google Scholar] [CrossRef]
- Willer, C.J.; Schmidt, E.M.; Sengupta, S.; Peloso, G.M.; Gustafsson, S.; Kanoni, S.; Ganna, A.; Chen, J.; Buchkovich, M.L.; Mora, S.; et al. Discovery and refinement of loci associated with lipid levels. Nat. Genet. 2013, 45, 1274–1283. [Google Scholar] [CrossRef] [Green Version]
- Bandesh, K.; Prasad, G.; Giri, A.K.; Kauser, Y.; Upadhyay, M.; Basu, A.; Tandon, N.; Bharadwaj, D. Genome-wide association study of blood lipids in Indians confirms universality of established variants. J. Hum. Genet. 2019, 64, 573–587. [Google Scholar] [CrossRef]
- Giri, A.K.; Prasad, G.; Bandesh, K.; Parekatt, V.; Mahajan, A.; Banerjee, P.; Kauser, Y.; Chakraborty, S.; Rajashekar, D.; INDICO; et al. Multifaceted genome-wide study identifies novel regulatory loci for body mass index in Indians. BioRxiv 2019. [Google Scholar] [CrossRef]
- Dupuis, J.; Langenberg, C.; Prokopenko, I.; Saxena, R.; Soranzo, N.; Jackson, A.U.; Wheeler, E.; Glazer, N.L.; Bouatia-Naji, N.; Gloyn, A.L.; et al. New genetic loci implicated in fasting glucose homeostasis and their impact on type 2 diabetes risk. Nat. Genet. 2010, 42, 105–116. [Google Scholar] [CrossRef]
- Giri, A.; Hellwege, J.N.; Keaton, J.M.; Park, J.; Qiu, C.; Warren, H.R.; Torstenson, E.S.; Kovesdy, C.P.; Sun, Y.V.; Wilson, O.D.; et al. Trans-ethnic association study of blood pressure determinants in over 750,000 individuals. Nat. Genet. 2019, 51, 51–62. [Google Scholar] [CrossRef]
- Kraja, A.T.; Vaidya, D.; Pankow, J.S.; Goodarzi, M.O.; Assimes, T.L.; Kullo, I.J.; Sovio, U.; Mathias, R.A.; Sun, Y.V.; Franceschini, N.; et al. A bivariate genome-wide approach to metabolic syndrome: STAMPEED Consortium. Diabetes 2011, 60, 1329–1339. [Google Scholar] [CrossRef]
- Carty, C.L.; Bhattacharjee, S.; Haessler, J.; Cheng, I.; Hindorff, L.A.; Aroda, V.; Carlson, C.S.; Hsu, C.-N.; Wilkens, L.; Liu, S.; et al. Comparative Analysis of Metabolic Syndrome Components in over 15,000 African Americans Identifies Pleiotropic Variants: Results from the PAGE Study. Circ. Cardiovasc. Genet. 2014, 7, 505–513. [Google Scholar] [CrossRef]
- Tekola-Ayele, F.; Doumatey, A.P.; Shriner, D.; Bentley, A.R.; Chen, G.; Zhou, J.; Fasanmade, O.; Johnson, T.; Oli, J.; Okafor, G.; et al. Genome-wide association study identifies African-ancestry specific variants for metabolic syndrome. Mol. Genet. Metab. 2015, 116, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Kristiansson, K.; Perola, M.; Tikkanen, E.; Kettunen, J.; Surakka, I.; Havulinna, A.S.; Stančáková, A.; Barnes, C.; Widen, E.; Kajantie, E.; et al. Genome-wide screen for metabolic syndrome susceptibility loci reveals strong lipid gene contribution but no evidence for common genetic basis for clustering of metabolic syndrome traits. Circ. Cardiovasc. Genet. 2012, 5, 242–249. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, D.; Zhou, D.; Li, Z.; Li, Z.; Fang, L.; Yang, M.; Shan, Z.; Li, H.; Chen, J.; et al. Susceptibility loci for metabolic syndrome and metabolic components identified in Han Chinese: A multi-stage genome-wide association study. J. Cell. Mol. Med. 2017, 21, 1106–1116. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, Y.; Park, T. New Common and Rare Variants Influencing Metabolic Syndrome and Its Individual Components in a Korean Population. Sci. Rep. 2018, 8, 5701. [Google Scholar] [CrossRef]
- Lin, E.; Kuo, P.-H.; Liu, Y.-L.; Yang, A.C.; Tsai, S.-J. Detection of susceptibility loci on APOA5 and COLEC12 associated with metabolic syndrome using a genome-wide association study in a Taiwanese population. Oncotarget 2017, 8, 93349–93359. [Google Scholar] [CrossRef] [Green Version]
- Vattikuti, S.; Guo, J.; Chow, C.C. Heritability and genetic correlations explained by common SNPS for metabolic syndrome traits. PLoS Genet. 2012, 8, e1002637. [Google Scholar] [CrossRef]
- Indian Genome Variation Consortium. The Indian Genome Variation database (IGVdb): A project overview. Hum Genet. 2005, 118, 1–11. [Google Scholar] [CrossRef]
- Tabassum, R.; Chauhan, G.; Dwivedi, O.P.; Mahajan, A.; Jaiswal, A.; Kaur, I.; Bandesh, K.; Singh, T.; Mathai, B.J.; Pandey, Y.; et al. Genome-wide association study for type 2 diabetes in Indians identifies a new susceptibility locus at 2q21. Diabetes 2013, 62, 977–986. [Google Scholar] [CrossRef]
- Mohan, V.; Rao, G.H.R. Type 2 Diabetes in South Asains, 1st ed.; South Asian Society on Atherosclerosis and Thrombosis: New Delhi, India, 2007. [Google Scholar]
- Rao, G.H.R.; Thanickachalam, S. Coronary Artery Disease: Risk Promoters, Pathophysiology and Prevention, 1st ed.; South Asian Society on Atherosclerosis and Thrombosis: New Delhi, India, 2005. [Google Scholar]
- Prasad, D.S.; Kabir, Z.; Dash, A.K.; Das, B.C. Abdominal obesity, an independent cardiovascular risk factor in Indian subcontinent: A clinico epidemiological evidence summary. J. Cardiovasc. Dis. Res. 2011, 2, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Zabaneh, D.; Balding, D.J. A genome-wide association study of the metabolic syndrome in Indian Asian men. PLoS ONE 2010, 5, e11961. [Google Scholar] [CrossRef]
- Basu, A.; Sarkar-Roy, N.; Majumder, P.P. Genomic reconstruction of the history of extant populations of India reveals five distinct ancestral components and a complex structure. Proc. Natl. Acad. Sci. USA 2016, 113, 1594–1599. [Google Scholar] [CrossRef] [Green Version]
- INdian DIabetes Consortium. INDICO: The development of a resource for epigenomic study of Indians undergoing socioeconomic transition. Hugo J. 2011, 5, 65–69. [Google Scholar] [CrossRef]
- Grundy, S.M.; Cleeman, J.I.; Daniels, S.R.; Donato, K.A.; Eckel, R.H.; Franklin, B.A.; Gordon, D.J.; Krauss, R.M.; Savage, P.J.; Smith, S.C., Jr.; et al. Diagnosis and management of the metabolic syndrome: An American Heart Association/National Heart, Lung, and Blood Institute scientific statement: Executive Summary. Crit. Pathw. Cardiol. 2005, 4, 198–203. [Google Scholar] [CrossRef]
- Mahajan, A.; Jaiswal, A.; Tabassum, R.; Podder, A.; Ghosh, S.; Madhu, S.V.; Mathur, S.K.; Tandon, N.; Bharadwaj, D. Elevated levels of C-reactive protein as a risk factor for Metabolic Syndrome in Indians. Atherosclerosis 2012, 220, 275–281. [Google Scholar] [CrossRef]
- Giri, A.K.; Banerjee, P.; Chakraborty, S.; Kauser, Y.; Undru, A.; Roy, S.; Parekatt, V.; Ghosh, S.; Tandon, N.; Bharadwaj, D. Genome wide association study of uric acid in Indian population and interaction of identified variants with Type 2 diabetes. Sci. Rep. 2016, 6, 21440. [Google Scholar] [CrossRef] [Green Version]
- Prasad, G.; Giri, A.K.; Basu, A.; Tandon, N.; Bharadwaj, D. Genomewide association study for C-reactive protein in Indians replicates known associations of common variants. J. Genet. 2019, 98, 20. [Google Scholar] [CrossRef]
- Bandesh, K.; Prasad, G.; Giri, A.K.; Voruganti, V.S.; Butte, N.F.; Cole, S.A.; Comuzzie, A.G.; INDICO; Tandon, N.; Bharadwaj, D. Genomewide association study of C-peptide surfaces key regulatory genes in Indians. J. Genet. 2019, 98, 8. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Turner, S.D. qqman: An R package for visualizing GWAS results using Q-Q and manhattan plots. J. Open Source Softw. 2014. [Google Scholar] [CrossRef]
- Buniello, A.; Macarthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef]
- Watanabe, K.; Stringer, S.; Frei, O.; Mirkov, M.U.; Polderman, T.J.C.; van der Sluis, S.; Andreassen, O.A.; Neale, B.M.; Posthuma, D. A global overview of pleiotropy and genetic architecture in complex traits. BioRxiv 2018. [Google Scholar] [CrossRef]
- Type 2 Diabetes Knowledge Portal. Available online: http://www.type2diabetesgenetics.org/ (accessed on 25 May 2019).
- Pruim, R.J.; Welch, R.P.; Sanna, S.; Teslovich, T.M.; Chines, P.S.; Gliedt, T.P.; Boehnke, M.; Abecasis, G.R.; Willer, C.J.; Frishman, D. LocusZoom: Regional visualization of genome-wide association scan results. Proc. Bioinformat. 2011, 26, 2336–2337. [Google Scholar] [CrossRef]
- Gauderman, W.J. Sample size requirements for association studies of gene-gene interaction. Am. J. Epidemiol. 2002, 5, 478–484. [Google Scholar] [CrossRef]
- Delaneau, O.; Zagury, J.-F.; Marchini, J. Improved whole-chromosome phasing for disease and population genetic studies. Nat. Methods 2013, 10, 5–6. [Google Scholar] [CrossRef]
- Howie, B.N.; Donnelly, P.; Marchini, J. A flexible and accurate genotype imputation method for the next generation of genome-wide association studies. PLoS Genet. 2009, 5, e1000529. [Google Scholar] [CrossRef]
- GTEx Consortium. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Davis, C.A.; Hitz, B.C.; Sloan, C.A.; Chan, E.T.; Davidson, J.M.; Gabdank, I.; Hilton, J.A.; Jain, K.; Baymuradoy, U.K.; Narayanan, A.K.; et al. The Encyclopedia of DNA elements (ENCODE): Data portal update. Nucleic Acids Res. 2018, 46, D794–D801. [Google Scholar] [CrossRef]
- Khan, A.; Fornes, O.; Stigliani, A.; Gheorghe, M.; Castro-Mondragon, J.A.; Van Der Lee, R.; Bessy, A.; Chèneby, J.; Kulkarni, S.R.; Tan, G.; et al. JASPAR 2018: Update of the open-access database of transcription factor binding profiles and its web framework. Nucleic Acids Res. 2018, 46, D260–D266. [Google Scholar] [CrossRef]
- Casper, J.; Zweig, A.S.; Villarreal, C.; Tyner, C.; Speir, M.L.; Rosenbloom, K.R.; Raney, B.J.; Lee, C.M.; Lee, B.T.; Karolchik, D.; et al. The UCSC Genome Browser database: 2018 update. Nucleic Acids Res. 2018, 46, D762–D769. [Google Scholar] [CrossRef]
- Hou, H.; Ma, R.; Guo, H.; He, J.; Hu, Y.; Mu, L.; Yan, Y.; Ma, J.; Li, S.; Zhang, J.; et al. Association between six CETP polymorphisms and metabolic syndrome in uyghur adults from Xinjiang, China. Int. J. Environ. Res. Public Health 2017, 14, E653. [Google Scholar] [CrossRef]
- Machiela, M.J.; Chanock, S.J. LDlink: A web-based application for exploring population-specific haplotype structure and linking correlated alleles of possible functional variants. Bioinformatics 2015, 31, 3555–3557. [Google Scholar] [CrossRef]
- Rebhan, M.; Chalifa-Caspi, V.; Prilusky, J.; Lancet, D. GeneCards: Integrating information about genes, proteins and diseases. Trends Genet. 1997, 13, 163. [Google Scholar] [CrossRef]
- Papp, A.C.; Pinsonneault, J.K.; Wang, D.; Newman, L.C.; Gong, Y.; Johnson, J.A.; Pepine, C.J.; Kumari, M.; Hingorani, A.D.; Talmud, P.J.; et al. Cholesteryl ester transfer protein (CETP) polymorphisms affect mRNA splicing, HDL levels, and sex-dependent cardiovascular risk. PLoS ONE 2012, 7, e31930. [Google Scholar] [CrossRef]
- Lu, H.; Inazu, A.; Moriyama, Y.; Higashikata, T.; Kawashiri, M.A.; Yu, W.; Huang, Z.; Okamura, T.; Mabuchi, H. Haplotype analyses of cholesteryl ester transfer protein gene promoter: A clue to an unsolved mystery of TaqIB polymorphism. J. Mol. Med. 2003, 81, 246–255. [Google Scholar] [CrossRef]
- Frisdal, E.; Klerkx, A.H.E.M.; Le Goff, W.; Tanck, M.W.T.; Lagarde, J.P.; Jukema, J.W.; Kastelein, J.J.P.; Chapman, M.J.; Guerin, M. Functional interaction between -629C/A, -971G/A and -1337C/T polymorphisms in the CETP gene is a major determinant of promoter activity and plasma CETP concentration in the REGRESS Study. Hum. Mol. Genet. 2005, 14, 2607–2618. [Google Scholar] [CrossRef]
- Dachet, C.; Poirier, O.; Cambien, F.; Chapman, J.; Rouis, M. New functional promoter polymorphism, CETP/-629, in cholesteryl ester transfer protein (CETP) gene related to CETP mass and high density lipoprotein cholesterol levels: Role of Sp1/Sp3 in transcriptional regulation. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 507–515. [Google Scholar] [CrossRef]
- Thompson, A.; Di Angelantonio, E.; Sarwar, N.; Erqou, S.; Saleheen, D.; Dullaart, R.P.F.; Keavney, B.; Ye, Z.; Danesh, J. Association of cholesteryl ester transfer protein genotypes with CETP mass and activity, lipid levels, and coronary risk. JAMA J. Am. Med. Assoc. 2008, 299, 2777–2788. [Google Scholar] [CrossRef]
- van der Harst, P.; Verweij, N. Identification of 64 Novel Genetic Loci Provides an Expanded View on the Genetic Architecture of Coronary Artery Disease. Circ. Res. 2018, 122, 433–443. [Google Scholar] [CrossRef]
- Iwanicka, J.; Iwanicki, T.; Niemiec, P.; Balcerzyk, A.; Krauze, J.; Górczyńska-Kosiorz, S.; Ochalska-Tyka, A.; Grzeszczak, W.; Żak, I. Relationship between CETP gene polymorphisms with coronary artery disease in Polish population. Mol. Biol. Rep. 2018, 45, 1929–1935. [Google Scholar] [CrossRef] [Green Version]
- Inouye, M.; Ripatti, S.; Kettunen, J.; Lyytikäinen, L.P.; Oksala, N.; Laurila, P.P.; Kangas, A.J.; Soininen, P.; Savolainen, M.J.; Viikari, J.; et al. Novel Loci for Metabolic Networks and Multi-Tissue Expression Studies Reveal Genes for Atherosclerosis. PLoS Genet. 2012, 8, e1002907. [Google Scholar] [CrossRef]
- Yang, J.; Benyamin, B.; McEvoy, B.P.; Gordon, S.; Henders, A.K.; Nyholt, D.R.; Madden, P.A.; Heath, A.C.; Martin, N.G.; Montgomery, G.W.; et al. Common SNPs explain a large proportion of the heritability for human height. Nat. Genet. 2010, 42, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Ross, S.E.; Hemati, N.; Longo, K.A.; Bennett, C.N.; Lucas, P.C.; Erickson, R.L.; MacDougald, O.A. Inhibition of adipogenesis by Wnt signaling. Science 2000, 289, 950–953. [Google Scholar] [CrossRef]
- Bennett, C.N.; Hodge, C.L.; MacDougald, O.A.; Schwartz, J. Role of Wnt10b and C/EBPα in spontaneous adipogenesis of 243 cells. Biochem. Biophys. Res. Commun. 2003, 302, 12–16. [Google Scholar] [CrossRef]
- Lagathu, C.; Christodoulides, C.; Tan, C.Y.; Virtue, S.; Laudes, M.; Campbell, M.; Ishikawa, K.; Ortega, F.; Tinahones, F.J.; Fernández-Real, J.M.; et al. Secreted frizzled-related protein 1 regulates adipose tissue expansion and is dysregulated in severe obesity. Int. J. Obes. 2010, 34, 1695–1705. [Google Scholar] [CrossRef] [Green Version]
- Gauger, K.J.; Bassa, L.M.; Henchey, E.M.; Wyman, J.; Bentley, B.; Brown, M.; Shimono, A.; Schneider, S.S. Mice deficient in Sfrp1 exhibit increased adiposity, dysregulated glucose metabolism, and enhanced macrophage infiltration. PLoS ONE 2013, 8, e78320. [Google Scholar] [CrossRef]
- Blumenthal, A.; Ehlers, S.; Lauber, J.; Buer, J.; Lange, C.; Goldmann, T.; Heine, H.; Brandt, E.; Reiling, N. The Wingless homolog WNT5A and its receptor Frizzled-5 regulate inflammatory responses of human mononuclear cells induced by microbial stimulation. Blood 2006, 108, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.; Schaer, D.J.; Bachli, E.B.; Kurrer, M.O.; Schoedon, G. Wnt5A/CaMKII signaling contributes to the inflammatory response of macrophages and is a target for the antiinflammatory action of activated protein C and interleukin-10. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 504–510. [Google Scholar] [CrossRef]
- Barandon, L.; Casassus, F.; Leroux, L.; Moreau, C.; Allières, C.; Lamazière, J.M.D.; Dufourcq, P.; Couffinhal, T.; Duplàa, C. Secreted frizzled-related protein-1 improves postinfarction scar formation through a modulation of inflammatory response. Arterioscler. Thromb. Vasc. Biol. 2011, 31, e80–e87. [Google Scholar] [CrossRef]
- Barandon, L.; Couffinhal, T.; Ezan, J.; Dufourcq, P.; Costet, P.; Alzieu, P.; Leroux, L.; Moreau, C.; Dare, D.; Duplàa, C. Reduction of Infarct Size and Prevention of Cardiac Rupture in Transgenic Mice Overexpressing FrzA. Circulation 2003, 108, 2282–2289. [Google Scholar] [CrossRef] [Green Version]
- Moon, R.T.; Kohn, A.D.; De Ferrari, G.V.; Kaykas, A. WNT and β-catenin signalling: Diseases and therapies. Nat. Rev. Genet. 2004, 5, 691–701. [Google Scholar] [CrossRef]
- DiRocco, D.P.; Kobayashi, A.; Taketo, M.M.; McMahon, A.P.; Humphreys, B.D. Wnt4/β−Catenin Signaling in Medullary Kidney Myofibroblasts. J. Am. Soc. Nephrol. 2013, 24, 1399–1412. [Google Scholar] [CrossRef]
- Matsuyama, M.; Nomori, A.; Nakakuni, K.; Shimono, A.; Fukushima, M. Secreted frizzled-related protein 1 (Sfrp1) regulates the progression of renal fibrosis in a mouse model of obstructive nephropathy. J. Biol. Chem. 2014, 289, 31526–31533. [Google Scholar] [CrossRef]
- Locatelli, F.; Pozzoni, P.; Del Vecchio, L. Renal manifestations in the metabolic syndrome. J. Am. Soc. Nephrol. 2006, 17, S81–S85. [Google Scholar] [CrossRef]
- Johnson, R.J.; Segal, M.S.; Srinivas, T.; Ejaz, A.; Mu, W.; Roncal, C.; Sánchez-Lozada, L.G.; Gersch, M.; Rodriguez-Iturbe, B.; Kang, D.-H.; et al. Essential Hypertension, Progressive Renal Disease, and Uric Acid: A Pathogenetic Link? J. Am. Soc. Nephrol. 2005, 16, 1909–1919. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, S.; Jiang, X.; Wang, Y.H.; Li, F.; Wang, Y.G.; Zheng, Y.; Cai, L. The role of the Nrf2/Keap1 pathway in obesity and metabolic syndrome. Rev. Endocr. Metab. Disord. 2015, 16, 35–45. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Replication Phase | |||||
|---|---|---|---|---|---|
| Gene Region | CHR | Start Base Pair (hg19) | Number of SNPs | p-Value (Lead SNP) | N |
| CETP | 16 | 56988044 | 7 | 3.48 × 10−9 | 4671 |
| MC4R | 18 | 57851097 | 2 | 3.66 × 10−4 | 4666 |
| LPL | 8 | 19919655 | 1 | 8.82 × 10−4 | 4650 |
| Discovery Phase | Replication Phase | Meta-Analysis | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SNP | CHR | Base Position | Nearby Gene | SNP Location | Alleles (Effect/Other) | MAF | N | p-Value | OR | N | p-Value | OR | p-Value | OR | Dir | I | Q |
| rs16890462 | 8 | 41309355 | SFRP1 | Intergenic | A/G | 0.21 | 2156 | 5.48 × 10−3 | 1.29 | 3259 | 5.51 × 10−3 | 1.25 | 8.75 × 10−5 | 1.26 | ++ | 0 | 0.78 |
| rs1530611 | 4 | 5206254 | STK32B | intronic | A/G | 0.29 | 2148 | 6.69 × 10−3 | 1.23 | 3257 | 6.37 × 10−3 | 1.21 | 1.18 × 10−4 | 1.22 | ++ | 0 | 0.89 |
| rs11048180 | 12 | 25735148 | IFLTD1 | intergenic | A/G | 0.08 | 2157 | 5.46 × 10−4 | 0.66 | 3259 | 0.04 | 0.8 | 1.25 × 10−4 | 0.73 | -- | 31.56 | 0.23 |
| rs16896746 | 6 | 66289412 | EYS | intronic | G/A | 0.08 | 2156 | 4.32 × 10−5 | 0.62 | 3259 | 0.1 | 0.86 | 1.51 × 10−4 | 0.73 | -- | 75.4 | 0.04 |
| rs710630 | 12 | 65983583 | CAND1 | intronic | G/A | 0.46 | 2158 | 0.01 | 1.18 | 3259 | 4.61 × 10−3 | 1.19 | 2.08 × 10−4 | 1.19 | ++ | 0 | 0.91 |
| rs710628 | 12 | 65943747 | CAND1 | intergenic | A/G | 0.46 | 2157 | 0.01 | 1.18 | 3258 | 5.53 × 10−3 | 1.19 | 2.47 × 10−4 | 1.18 | ++ | 0 | 0.93 |
| rs7005211 | 8 | 123538147 | ZHX2 | intergenic | G/A | 0.47 | 2155 | 6.69 × 10−3 | 0.83 | 4654 | 8.90 × 10−3 | 0.89 | 2.72 × 10−4 | 0.87 | -- | 0 | 0.33 |
| rs1060350 | 12 | 65992732 | CAND1 | synonymous | G/A | 0.48 | 2157 | 6.15 × 10−3 | 1.21 | 3254 | 0.01 | 1.16 | 2.85 × 10−4 | 1.18 | ++ | 0 | 0.67 |
| rs1152877 | 12 | 65989452 | CAND1 | intronic | G/A | 0.48 | 2155 | 5.50 × 10−3 | 1.22 | 3220 | 0.01 | 1.16 | 2.89 × 10−4 | 1.18 | ++ | 0 | 0.62 |
| rs564210 | 16 | 64314818 | LOC283867 | intergenic | A/G | 0.26 | 2156 | 3.55 × 10−4 | 0.76 | 3251 | 0.1 | 0.89 | 3.30 × 10−4 | 0.83 | -- | 60.29 | 0.11 |
| rs12595506 | 15 | 25744981 | UBE3A | intergenic | G/A | 0.39 | 2156 | 0.01 | 1.19 | 7930 | 4.99 ×10−3 | 1.1 | 3.53 × 10−4 | 1.11 | ++ | 0.22 | 0.32 |
| rs2967379 | 16 | 80770811 | MPHOSPH6 | intergenic | G/A | 0.49 | 2156 | 4.52 × 10−6 | 0.73 | 3258 | 0.4 | 0.96 | 3.69 × 10−4 | 0.84 | -- | 88.69 | 2 × 10−3 |
| rs10499618 | 7 | 40787165 | C7orf10 | intronic | G/A | 0.11 | 2158 | 3.98 × 10−3 | 1.41 | 3257 | 0.03 | 1.26 | 3.82 × 10−4 | 1.32 | ++ | 0 | 0.48 |
| rs1337212 | 9 | 119239170 | ASTN2 | intergenic | A/G | 0.11 | 2157 | 4.53 × 10−4 | 1.55 | 3259 | 0.09 | 1.18 | 3.93 × 10−4 | 1.31 | ++ | 62.74 | 0.1 |
| rs7554931 | 1 | 170365623 | LOC284688 | intergenic | A/G | 0.41 | 2157 | 2.91 × 10−3 | 0.81 | 4673 | 0.01 | 0.9 | 3.98 × 10−4 | 0.87 | -- | 44.4 | 0.18 |
| rs3746228 | 19 | 57804362 | ZNF460 | 3’-UTR | A/G | 0.18 | 2158 | 7.18 × 10−4 | 1.38 | 4529 | 0.03 | 1.13 | 4.02 × 10−4 | 1.18 | ++ | 70.48 | 0.06 |
| rs885036 | 2 | 98671225 | MGAT4A | intronic | G/A | 0.49 | 2157 | 2.96 × 10−5 | 1.35 | 3254 | 0.3 | 1.07 | 4.18 × 10−4 | 1.18 | ++ | 83.49 | 0.01 |
| rs1066396 | 12 | 66005634 | CAND1 | intergenic | G/A | 0.48 | 2157 | 6.14 × 10−3 | 1.21 | 3254 | 0.02 | 1.15 | 4.37 × 10−4 | 1.17 | ++ | 0 | 0.59 |
| rs11108860 | 12 | 96081536 | NEDD1 | intergenic | G/A | 0.04 | 2157 | 8.72 × 10−7 | 0.45 | 3259 | 0.9 | 1.01 | 4.52 × 10−4 | 0.66 | -+ | 91.59 | 6 × 10−4 |
| rs4677119 | 3 | 72291958 | LOC201617 | intergenic | A/G | 0.33 | 2158 | 5.76 × 10−3 | 0.81 | 4653 | 0.01 | 0.89 | 4.80 × 10−4 | 0.87 | -- | 22.01 | 0.26 |
| rs9309089 | 2 | 43028132 | HAAO | intergenic | A/G | 0.35 | 2158 | 0.1 | 1.12 | 4623 | 1.54 × 10−3 | 1.15 | 5 × 10−4 | 1.14 | ++ | 0 | 0.72 |
| rs6948816 | 7 | 39661439 | RALA | intronic | A/G | 0.03 | 2158 | 5.36 × 10−3 | 0.62 | 3257 | 0.03 | 0.69 | 5.13 × 10−4 | 0.65 | -- | 0 | 0.62 |
| rs10983653 | 9 | 119237233 | ASTN2 | intergenic | A/G | 0.11 | 2158 | 6.15 × 10−4 | 1.53 | 3259 | 0.09 | 1.19 | 5.14 × 10−4 | 1.31 | ++ | 60.43 | 0.11 |
| rs1333144 | 1 | 170364401 | LOC284688 | intergenic | G/A | 0.41 | 2157 | 5.40 × 10−3 | 0.82 | 4654 | 0.02 | 0.9 | 5.92 × 10−4 | 0.88 | -- | 27.92 | 0.24 |
| rs17530234 | 7 | 40783104 | C7orf10 | intronic | G/A | 0.11 | 2155 | 5.49 × 10−3 | 1.38 | 3258 | 0.03 | 1.24 | 6.25 × 10−4 | 1.3 | ++ | 0 | 0.49 |
| rs2462683 | 7 | 112971889 | PPP1R3A | intergenic | G/A | 0.34 | 2158 | 6.34 × 10−4 | 1.3 | 3257 | 0.1 | 1.11 | 6.51 × 10−4 | 1.18 | ++ | 59.06 | 0.12 |
| rs11749727 | 5 | 179540965 | RASGEF1C | intronic | G/A | 0.44 | 2156 | 6.75 × 10−3 | 1.21 | 3258 | 0.03 | 1.15 | 6.59 × 10−4 | 1.17 | ++ | 0 | 0.55 |
| rs475479 | 16 | 64324819 | LOC283867 | intergenic | G/A | 0.26 | 2158 | 5.41 × 10−4 | 0.76 | 3259 | 0.1 | 0.9 | 7.12 × 10−4 | 0.83 | -- | 61.71 | 0.12 |
| rs35814902 | 5 | 86835416 | CCNH | intergenic | A/G | 0.32 | 2158 | 5.04 × 10−3 | 1.24 | 4573 | 0.02 | 1.11 | 8.01 × 10−4 | 1.13 | ++ | 38.95 | 0.2 |
| rs17529882 | 7 | 40761636 | C7orf10 | intronic | G/A | 0.09 | 2157 | 2.10 × 10−3 | 1.48 | 3259 | 0.08 | 1.22 | 8.28 × 10−4 | 1.32 | ++ | 26.33 | 0.24 |
| rs17456070 | 1 | 87599332 | LOC100505768 | intronic | G/A | 0.30 | 2155 | 0.05 | 0.86 | 3257 | 5.98 ×10−3 | 0.83 | 8.56 × 10−4 | 0.84 | -- | 0 | 0.74 |
| rs12650617 | 4 | 5238437 | STK32B | intronic | A/G | 0.22 | 2157 | 3.35 × 10−5 | 1.45 | 3245 | 0.3 | 1.07 | 8.81 × 10−4 | 1.21 | ++ | 85.49 | 8.70 × 10−3 |
| rs13177543 | 5 | 86842168 | CCNH | intergenic | A/G | 0.32 | 2158 | 5.04 × 10−3 | 1.24 | 4658 | 0.03 | 1.1 | 9.93 × 10−4 | 1.13 | ++ | 42.89 | 0.18 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prasad, G.; Bandesh, K.; Giri, A.K.; Kauser, Y.; Chanda, P.; Parekatt, V.; INDICO; Mathur, S.; Madhu, S.V.; Venkatesh, P.; et al. Genome-Wide Association Study of Metabolic Syndrome Reveals Primary Genetic Variants at CETP Locus in Indians. Biomolecules 2019, 9, 321. https://doi.org/10.3390/biom9080321
Prasad G, Bandesh K, Giri AK, Kauser Y, Chanda P, Parekatt V, INDICO, Mathur S, Madhu SV, Venkatesh P, et al. Genome-Wide Association Study of Metabolic Syndrome Reveals Primary Genetic Variants at CETP Locus in Indians. Biomolecules. 2019; 9(8):321. https://doi.org/10.3390/biom9080321
Chicago/Turabian StylePrasad, Gauri, Khushdeep Bandesh, Anil K. Giri, Yasmeen Kauser, Prakriti Chanda, Vaisak Parekatt, INDICO, Sandeep Mathur, Sri Venkata Madhu, Pradeep Venkatesh, and et al. 2019. "Genome-Wide Association Study of Metabolic Syndrome Reveals Primary Genetic Variants at CETP Locus in Indians" Biomolecules 9, no. 8: 321. https://doi.org/10.3390/biom9080321
APA StylePrasad, G., Bandesh, K., Giri, A. K., Kauser, Y., Chanda, P., Parekatt, V., INDICO, Mathur, S., Madhu, S. V., Venkatesh, P., Bhansali, A., Marwaha, R. K., Basu, A., Tandon, N., & Bharadwaj, D. (2019). Genome-Wide Association Study of Metabolic Syndrome Reveals Primary Genetic Variants at CETP Locus in Indians. Biomolecules, 9(8), 321. https://doi.org/10.3390/biom9080321