1. Mesothelia: Broad Definition
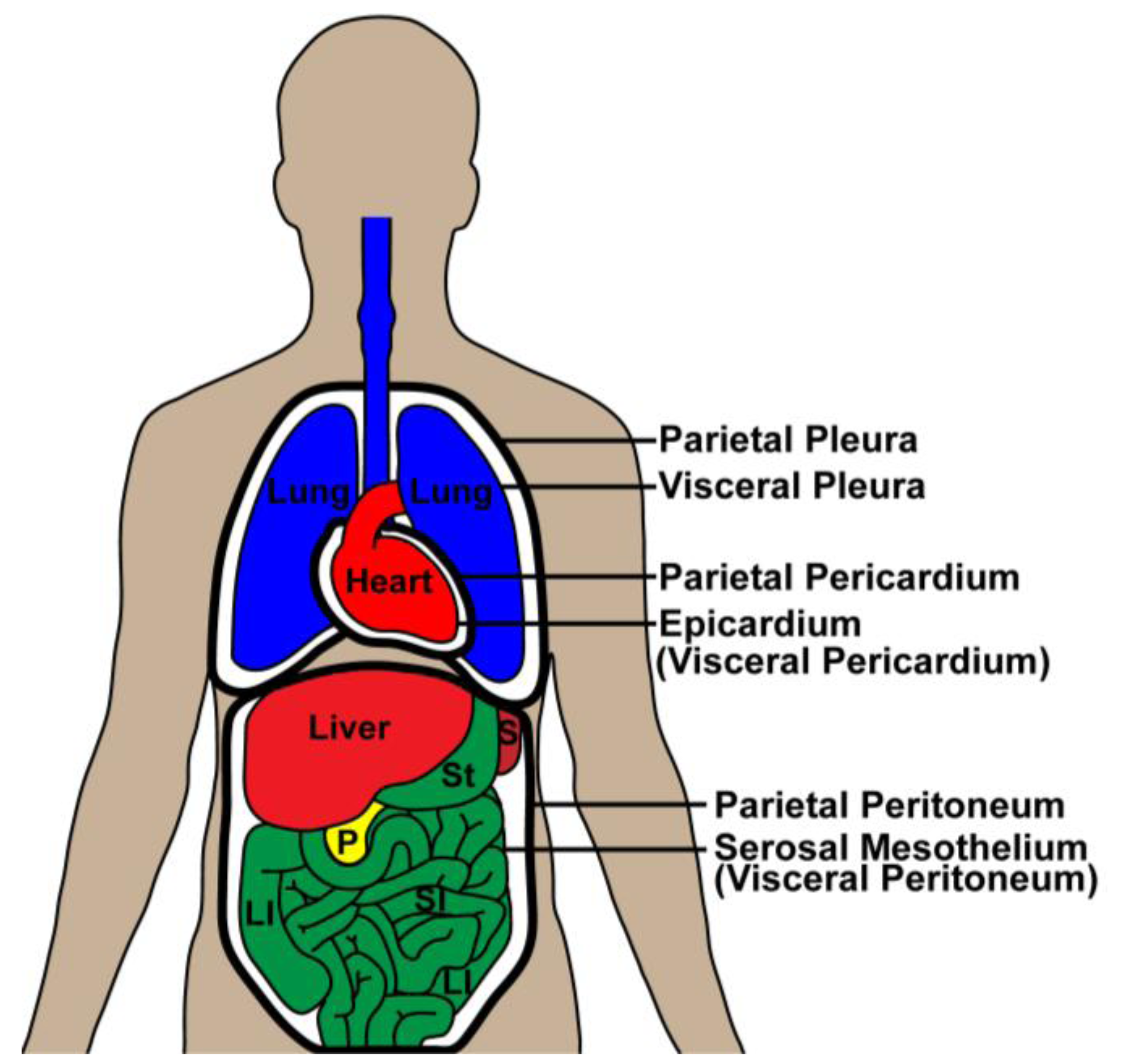
Mesothelia are simple squamous epithelia that line coelomic cavities and organs and form the mesenteries. In mammals, separation of the common embryonic coelom gives rise to pericardial, pleural and peritoneal cavities (discussed in detail below). Mesothelial cells lining the cavities/body walls in these subdivisions are referred to as parietal mesothelia while visceral mesothelia cover the organs. Visceral mesothelium covering the heart is referred to as the epicardium while pleural mesothelium covers lungs. Serosal mesothelium, which is the focus of this review, covers the organs of the alimentary canal within the abdominal cavity/coelom (
Figure 1).
Figure 1.
Mesothelia. Parietal mesothelia (thick black lines) cover the body wall and pericardial sac. Visceral mesothelia cover the organs and mesenteries and form a continuous lining with the parietal mesothelia. LI, large intestine; P, pancreas; S, spleen; SI, small intestine; St, stomach.
Figure 1.
Mesothelia. Parietal mesothelia (thick black lines) cover the body wall and pericardial sac. Visceral mesothelia cover the organs and mesenteries and form a continuous lining with the parietal mesothelia. LI, large intestine; P, pancreas; S, spleen; SI, small intestine; St, stomach.
Below, we describe the histology of adult mesothelium and give a brief overview of heart and intestinal structure. This is followed by a discussion of similarities and differences in basic developmental programs of the two organs as a guidepost for understanding the roles of mesothelia in organ development and function. We will then provide an overview of proepicardium/epicardium differentiation to serve as the departure point for the current discussion of mesothelial differentiation in the gut tube and its associated glands.
2. Adult Mesothelia: Cell Biology, Histology, and Function
Despite the multiple names, mesothelium has a relatively consistent structure in each of the coelomic cavities [
1,
2] (
Figure 2). The structure of mesothelium is that of a simple squamous epithelium; a single layer of thin plate-like cells [
3]. Histologically, there are three broad characteristics that define this epithelium: apical/basolateral polarity, robust cell/cell adhesion, and production of a basement membrane. Mesothelia are polarized with their apical surfaces toward the coelomic space. Apical surface modifications may include microvilli and cilia [
4,
5,
6]. Tight junctions are located on the lateral aspect of the cell and evidenced by the localization of Bves and ZO-1 [
7,
8]. They maintain apical/basal polarity and create a diffusion barrier between the coelomic space and the submesothelial connective tissue of the organ or body wall. E-cadherin confers further cell-cell adhesion and cytokeratins (mostly subtypes 8, 18, and 19) provide structural support [
9,
10] (
Figure 2B). A basement membrane is produced by and underlies the mesothelium separating it from the sub-mesothelial connective tissue space (
Figure 2C).
Figure 2.
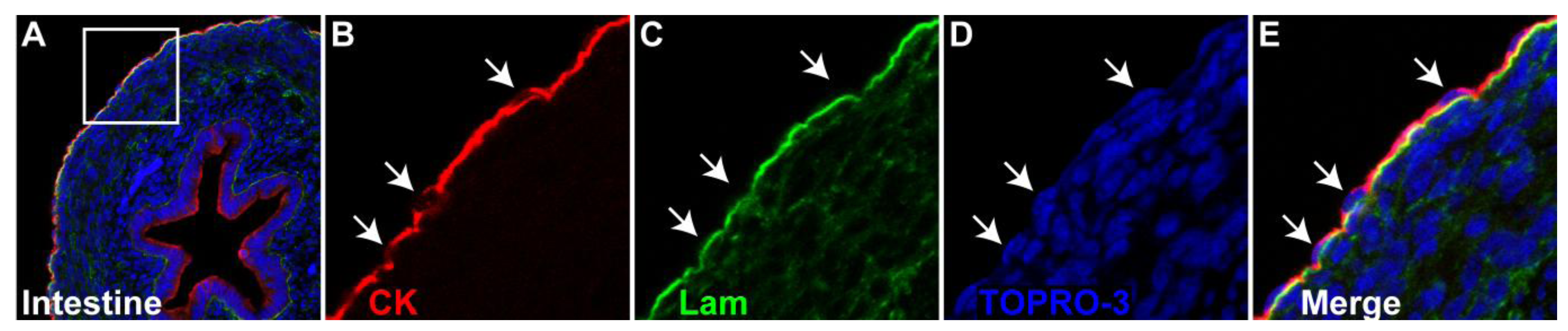
Mesothelial cell structure. (a) Transverse section through a chick embryonic day 8 small intestine. The mesothelium has attained a mature configuration by this developmental stage. (b–e) Magnified view of boxed area shown in A. (b) Mesothelial cells (arrows) are cytokeratin (CK, red) positive. (c) A laminin (lam, green) positive basement membrane underlies the mesothelium. (d) Mesothelial nuclei are flat in shape (arrows). (e) Merge of B–D.
Figure 2.
Mesothelial cell structure. (a) Transverse section through a chick embryonic day 8 small intestine. The mesothelium has attained a mature configuration by this developmental stage. (b–e) Magnified view of boxed area shown in A. (b) Mesothelial cells (arrows) are cytokeratin (CK, red) positive. (c) A laminin (lam, green) positive basement membrane underlies the mesothelium. (d) Mesothelial nuclei are flat in shape (arrows). (e) Merge of B–D.
The most conspicuous function of mesothelia in the adult is the production of a non-adhesive surface allowing relatively frictionless movement of the viscera within the individual coelomic cavity. This is accomplished by secretion of an apical glycosaminoglycan layer and production of a small amount of circulating coelomic fluid to provide lubrication [
11,
12]. On the surface of the diaphragm, there are gaps in the mesothelial monolayer called stomata that can directly connect with submesothelial lymphatic vessels [
6]. Not only is ionic and protein composition of coelomic cavity fluid regulated by mesothelium, but passage of inflammatory cells is also restricted [
13,
14,
15]. Additionally, mesothelium is crucial to the injury response and is thought to both prevent and promote scarring depending on the injury and to promote neovascularization [
16,
17,
18,
19]. These and many other studies have provided growing evidence for the dynamic role of mesothelium in both normal and abnormal physiologic states in the adult [
20].
3. Heart Histology
The human heart is a muscular pump with complex valvular and neuronal components and an elaborate conduction system. Still, the histology of the heart wall is relatively simple. From the lumen outward, the first cell type seen in any part of the vertebrate heart is the endocardial endothelial cell (
Figure 3). This cell rests upon a scant connective tissue layer that merges with the myocardium [
5]. The myocardium is populated by cardiac myocytes and contains abundant fibroblasts, which actually outnumber the cardiomyocytes, as well as smooth muscle and endothelial cells of arteries, veins, and capillaries and migratory cells of the immune system.
On its surface, the heart is covered by the epicardium composed of an outermost mesothelium and underlying connective tissue space inhabited by fibroblasts, adipose cells and matrix. The major coronary vessels run within this connective tissue space with smaller branches diving into the substance of the myocardium [
21]. Each myocyte is invested with a capillary bed. Proximally, the coronary vessels connect with the aorta. There is continuity between the mesothelia of the epicardium and pericardium as well as the mesocardium, the mesentery-like structure that connects the heart to the pericardial sac and dorsal body wall [
22]. All the cells composing the heart, with the exception of neural crest derivatives and migratory cells of the immune system are generated from splanchnic mesoderm. There is no endodermal component to the heart. Murine and avian cardiac structure is similar to the human configuration though notably in the mouse the coronary arteries are embedded within the myocardium near the subepicardial space while the veins reside in the epicardium [
23]. In the avian heart, the arteries are localized to the epicardial connective tissue while the veins are embedded within the myocardium [
24,
25]. In summary, the muscular and connective tissue layer of the heart is bounded by a luminal (endocardium) and surface (mesothelium) epithelium with an arterial supply that is associated closely with the surface mesothelium (
Figure 3A).
Figure 3.
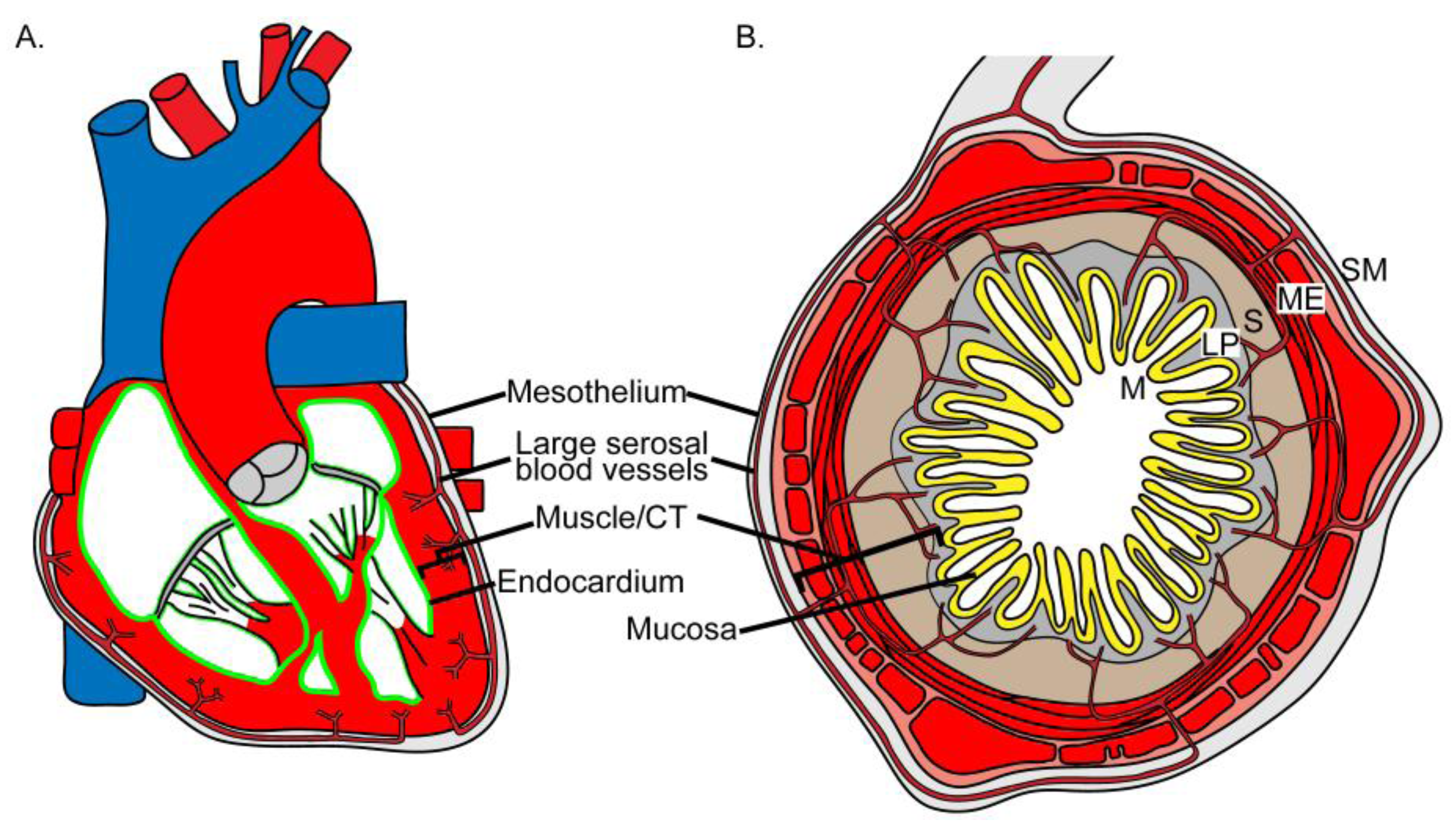
Structure of the human heart and intestine. (a) The lumen of the heart is lined by endocardium (green line), an endothelial cell type. The myocardium composed of myocytes and connective tissue cells forms the majority of the wall (bracket). Mesothelium lines the surface of the heart. The coronary arteries run in the connective tissue layer (light gray) subjacent to the mesothelium. Branches of the coronary arteries penetrate the myocardium. (b) The lumen of the intestine is lined by mucosal epithelium (M, yellow). There are multiple layers of connective tissue and muscle (bracket) underlying the mucosal epithelium. The first connective tissue layer is the lamina propria (LP, gray). The submucosa (S, tan), another connective tissue layer, encircles the lamina propria. The muscularis externus (ME) is composed of two layers of visceral smooth muscle (red). Serosal mesothelium (SM) lines the surface of the intestine. The major blood vessels of the intestine course through the submesothelial connective tissue (light gray) and branches of the vessels penetrate into the intestinal wall.
Figure 3.
Structure of the human heart and intestine. (a) The lumen of the heart is lined by endocardium (green line), an endothelial cell type. The myocardium composed of myocytes and connective tissue cells forms the majority of the wall (bracket). Mesothelium lines the surface of the heart. The coronary arteries run in the connective tissue layer (light gray) subjacent to the mesothelium. Branches of the coronary arteries penetrate the myocardium. (b) The lumen of the intestine is lined by mucosal epithelium (M, yellow). There are multiple layers of connective tissue and muscle (bracket) underlying the mucosal epithelium. The first connective tissue layer is the lamina propria (LP, gray). The submucosa (S, tan), another connective tissue layer, encircles the lamina propria. The muscularis externus (ME) is composed of two layers of visceral smooth muscle (red). Serosal mesothelium (SM) lines the surface of the intestine. The major blood vessels of the intestine course through the submesothelial connective tissue (light gray) and branches of the vessels penetrate into the intestinal wall.
![Jdb 01 00064 g003]()
4. Gut Tube Histology
Structure of the adult vertebrate gastrointestinal (GI) tract with all its associated glands, is highly variable. Still, these differences are actually variations of a basic or conserved structure. Modifications of four generalizable layers account for the variations observed in the stomach and intestines. Understanding the positions and relative contributions of these basic layers is important in understanding gut tube structure and function. The four conserved layers of the gut from the lumen outward are: (1) mucosa, (2) submucosa, (3) muscularis externa and (4) serosa [
5] (
Figure 3B).
These layers vary in composition and thickness along the length of the gut tube but are always present within organs of the alimentary canal housed within the abdominal cavity. The mucosa consists of an epithelial layer bordering the lumen and a subjacent connective tissue layer called the lamina propria. Mucosal epithelium along the entire gut tube and its associated glands is derived from endoderm (further discussion on this topic is given below). This is in contrast to the heart where no endodermal component is found. The major function of the mucosa is secretion and absorption of luminal substances. The submucosa is also rich in connective tissue cells and fibers. Glands, whose epithelial secretory and ductal components are continuous with the mucosal epithelium, may reside within the lamina propria and/or submucosa. Along with the lamina propria, the submucosa provides support for the distention and contraction of the GI wall. Smooth muscle of the muscularis externa responsible for moving luminal content along the alimentary canal is present between the submucosa and the connective tissue layer of the serosa. The outermost layer of the alimentary canal is the serosa with its connective tissue and mesothelium. As stated above, the major function of the serosa in the adult is provision of a smooth, lubricated surface upon which alimentary organs can move within the abdominal cavity.
During development, the major blood vessels to the gut, the celiac and superior and inferior mesenteric arteries, form as unpaired branches of the aorta and course to the gut within a dorsal mesentery. While the celiac and superior mesenteric arteries remain intraperitoneal, the inferior mesenteric artery and much of the territory it supplies becomes secondarily retroperitoneal as development proceeds. In the adult, the large branches of these arteries reside near the surface of the organs of the gastrointestinal tract in the sub-mesothelial space, just like the major vessels of the heart [
22]. As these major vessels traverse the surface of the gut, they send branches down into the substance of the muscularis externa, submucosa, and lamina propria ending in capillary networks serving these regions (
Figure 3B). While capillaries are intimately associated with the epithelial component of the mucosa, they do not penetrate the epithelium under normal conditions.
Thus, similar to the heart, the stomach and intestine have muscular and connective tissue layers bounded by a luminal (mucosal) and surface (mesothelial) epithelium. Vessels of both the cardiac and alimentary systems travel in the submesothelial space and dive into the substance of the organ giving rise to a capillary plexus that is essential for organ function. However, unlike the heart, the luminal epithelium of the GI tract is derived from endoderm and endothelial cells are not found within this layer (
Figure 3). Even from this brief description, it is clear that there are remarkable structural similarities between the heart and GI tract especially between mesothelial components and the associated vascular systems.
5. Coelom Formation, Coelomic Organs, and Mesothelia
Some of the structural variation and similarity can be explained by basic developmental principles of coelom formation and organogenesis in vertebrates. Understanding these principles also establishes the groundwork for a discussion of mesothelial development.
True coelomates contain a fluid filled cavity lined entirely by mesoderm. In vertebrates, this in generated through splitting the lateral plate mesoderm into two subdivisions separated by an intervening space (the coelom) very early in development. The dorsally located lateral plate mesoderm that associates with the ectoderm is called the somatic mesoderm and will form the body wall. The more ventral lateral plate mesoderm (splanchnic mesoderm) which associates with endoderm is destined to build organs of the coelomic cavity (
Figure 4A). Splanchnic mesoderm and endoderm are collectively termed splanchnopleure while somatic mesoderm together with ectoderm is referred to as somatopleure. It is interesting to note that a vascular plexus forms between the endodermal and splanchnic mesodermal layers of the embryo [
20,
26,
27] (
Figure 4A, green). Concurrent with these events, the embryo begins to take on a cylindrical shape with lateral folding (
Figure 4A, arrows). These morphogenetic movements reposition the three germ layers such that a “tube within a tube” is formed (
Figure 4B–C).
It is important to consider that there is variation in the positioning of endoderm relative to splanchnic mesoderm along the anteroposterior axis of the developing coelom. In the anterior region of the embryo, where the heart will form, endoderm is positioned dorsal to the forming coelomic cavity (
Figure 4B). This explains the position of the esophagus and trachea in the dorsal body wall of the adult vertebrate. The dorsal position of the endoderm in the anterior region of the embryo has important implications for heart development. Specifically, the vascular plexus situated between the endoderm and splanchnic mesoderm throughout the rest of the embryo is engaged to contribute to the luminal endocardial layer of the heart tube [
28] (
Figure 4B). Thus, even while blood courses through the heart lumen, the heart primordium itself is avascular throughout the initial stages of organogenesis. More posteriorly, beginning in the region of the forming septum transversum, endoderm is more ventrally deposed and is enveloped by splanchnic mesoderm (
Figure 4C). Given the continuing juxtaposition of endoderm and splanchnic mesoderm in the forming gut, the intervening vascular plexus is retained in this location throughout development.
It is interesting to note that the entire coelomic space is always bounded by mesoderm; indeed, this is the landmark of a true coelomate. Great variation in the structures derived from somatopleure and splanchnopleure is observed in true coelomates and subdivision of the original coelom is highly variable. Still, the basic spatial relationship of somatopleure, coelom, and splanchnopleure is maintained throughout the diversity of true coelomates and the juxtaposition of endoderm and mesoderm is a major step in evolution allowing the embryo to diversify by regionalizing function.
While the coelomic space is bounded by mesoderm, this does not mean that a mesothelium is present throughout development. In fact, the vertebrate coelom and its organs, specifically the heart and elements of the digestive system, are initially devoid of mesothelia [
26,
27,
29,
30]. While work in this area has been principally focused on avian and mammalian embryos, it is likely that other vertebrate classes develop similarly.
Thus, at the end of lateral folding, the heart is composed of two layers—endocardium and splanchnic mesoderm. The gut tube is also composed of two layers—endoderm and splanchnic mesoderm—but contains an endothelial plexus between the two layers. Neither the heart or gut tube is bordered on the coelomic surface by a mesothelium.
Figure 4.
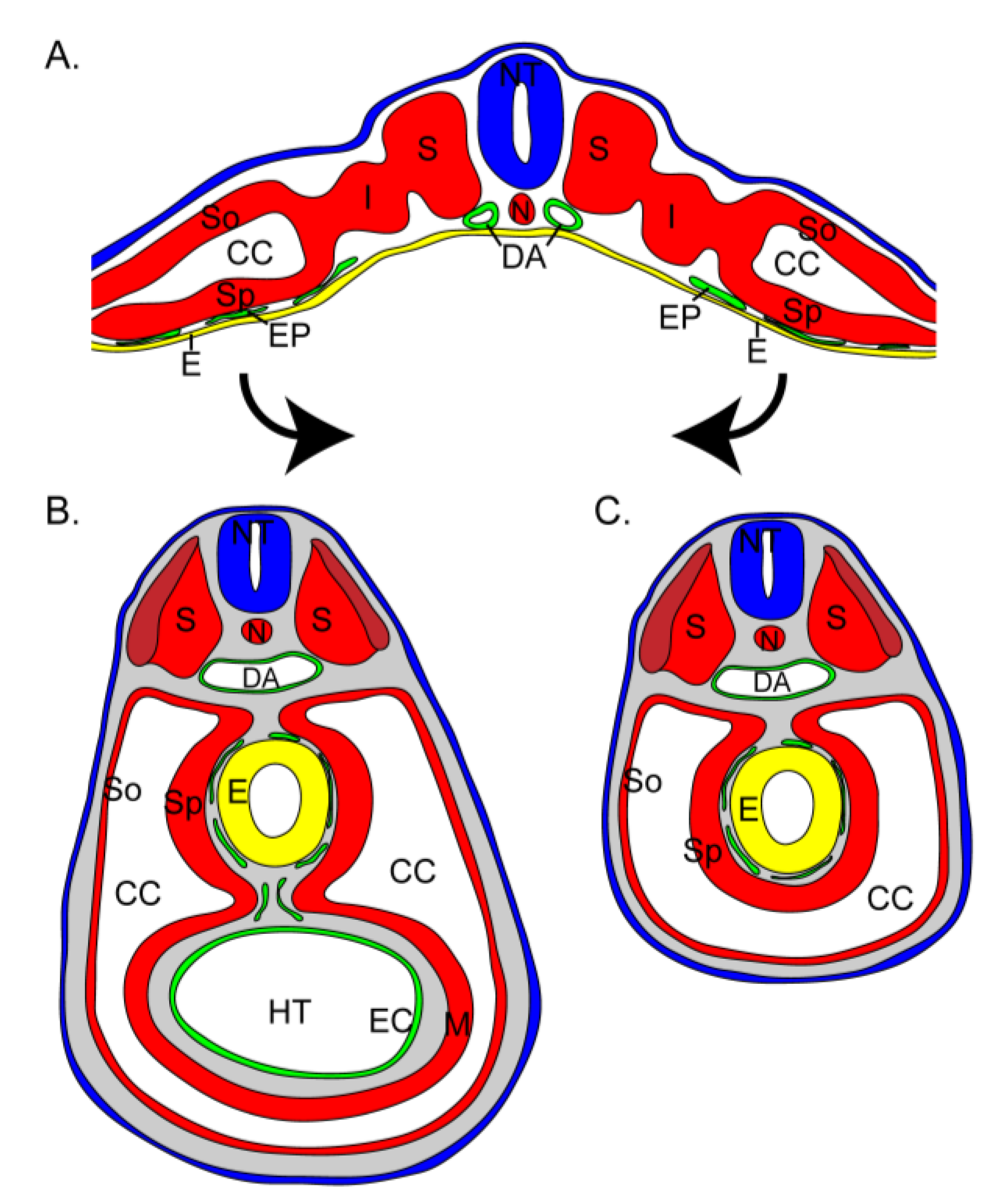
Heart and gut tube formation. (a) Schematic of a transverse section through the posterior region of a HH14 chick embryo demonstrating the relationship of the splanchnic mesoderm and endoderm prior to lateral folding. The lateral plate mesoderm is divided to form the somatic (So) and splanchnic mesoderm (Sp) with an intervening coelomic cavity (CC). An endothelial plexus (EP, green) resides between the splanchnic mesoderm and endoderm (E, yellow). Lateral folding (arrows) will unite the somatopleure and splanchnopleure ventrally. (b) In the anterior region of the HH14 embryo, lateral folding has already been completed. The endoderm has been excluded from the heart tube (HT) and cells of the endothelial plexus have formed the endocardial (EC) lining of the heart tube lumen. Dorsal to the heart tube, the endoderm of the gut tube remains invested by the endothelial plexus and splanchnic mesoderm. (c) Posterior to the heart tube in a HH18 chick embryo, the gut tube is the only structure within the coelomic cavity and includes endoderm, the endothelial plexus, and splanchnic mesoderm. DA, dorsal aorta; I, intermediate mesoderm; N, notochord; NT, neural tube; S, somite.
Figure 4.
Heart and gut tube formation. (a) Schematic of a transverse section through the posterior region of a HH14 chick embryo demonstrating the relationship of the splanchnic mesoderm and endoderm prior to lateral folding. The lateral plate mesoderm is divided to form the somatic (So) and splanchnic mesoderm (Sp) with an intervening coelomic cavity (CC). An endothelial plexus (EP, green) resides between the splanchnic mesoderm and endoderm (E, yellow). Lateral folding (arrows) will unite the somatopleure and splanchnopleure ventrally. (b) In the anterior region of the HH14 embryo, lateral folding has already been completed. The endoderm has been excluded from the heart tube (HT) and cells of the endothelial plexus have formed the endocardial (EC) lining of the heart tube lumen. Dorsal to the heart tube, the endoderm of the gut tube remains invested by the endothelial plexus and splanchnic mesoderm. (c) Posterior to the heart tube in a HH18 chick embryo, the gut tube is the only structure within the coelomic cavity and includes endoderm, the endothelial plexus, and splanchnic mesoderm. DA, dorsal aorta; I, intermediate mesoderm; N, notochord; NT, neural tube; S, somite.
![Jdb 01 00064 g004]()
6. Development of the Epicardium
A review of the literature quickly reveals that relatively little is known about the origin of mesothelia and its “arrival” on organs with the notable exception of cardiac development. We provide a brief overview of proepicardium/epicardium development to inform the current discussion of mesothelial differentiation in the gut tube and its associated glands. Excellent and extensive reviews focused on epicardial development are available to the reader in this edition.
As stated above, as the heart tube begins its characteristic looping, it is devoid of mesothelium. The outer cell layer of the developing heart is the myocardium, which is a polarized epithelium with its basal surface facing the endocardial space and its apical side facing the coelom [
26,
30]. At ED8.5 in mouse and HH stage 14 in chick, a simple squamous epithelial outgrowth from the region of the sinus venosus courses through the pericardial coelom [
29,
31] (
Figure 5A). This structure is termed the proepicardium or PE and is not simply an epithelium but also houses cells within its connective tissue space. These cells may be the product of epithelial to mesenchymal transition (EMT) from the migrating epithelium [
32] or simply cells that travel with this migratory structure. In the avian embryo, the PE extends villi in close approximation to proteoglycan-rich connective tissue fibrils to reach the heart [
31] (
Figure 5B, arrowhead). Apical/basal polarity of the epithelial PE, in the chicken embryo at least, is observed as soon as the PE can be identified and is maintained throughout the life of a healthy bird. Nahirney et al. have shown that once the PE reaches the myocardium, the clusters open (
Figure 5C, arrowhead) such that two sheets of simple squamous epithelium travel on the heart wall (
Figure 5C, arrows) and eventually envelop it [
31] (
Figure 5D). In this way, the polarity of the PE/epicardium is maintained with the apical surface facing the coelomic cavity. Retention of epithelial polarity is essential in positioning the mesothelium for its functions in homeostasis and production of mesenchyme in organogenesis.
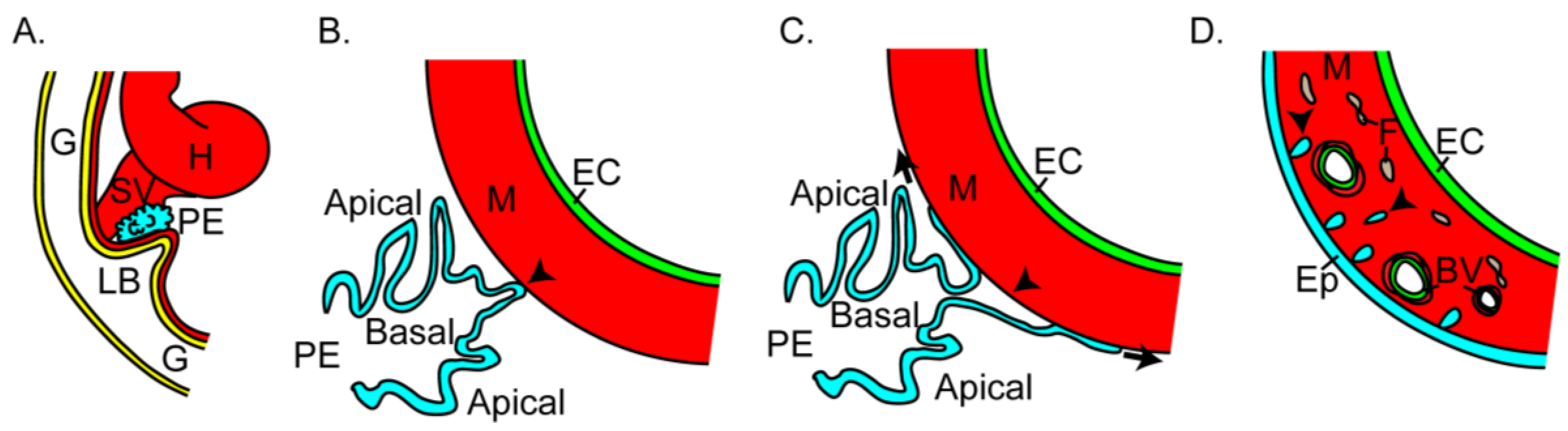
Figure 5.
Proepicardium (PE) development. (a) The PE (blue) forms external to the heart tube as an outgrowth from the region of the sinus venosus (SV) and liver bud (LB). (b) The PE epithelium is polarized with apical toward the pericardial space. The PE crosses the pericardial cavity to contact (arrowhead) the myocardium (M). (c) At points of contact with the myocardium, the PE epithelial sheet breaks open (arrowheads) to preserve apical-basal polarity and migrate over the heart tube (arrows). (d) During migration and after a continuous epicardium (Ep) is formed, cells leave the epicardium (arrowheads) via epithelial to mesenchymal transition (EMT) to migrate into the myocardium and generate fibroblasts (F) and the coronary blood vessels (BV). EC, endocardium; G, gut tube; H, heart tube.
Figure 5.
Proepicardium (PE) development. (a) The PE (blue) forms external to the heart tube as an outgrowth from the region of the sinus venosus (SV) and liver bud (LB). (b) The PE epithelium is polarized with apical toward the pericardial space. The PE crosses the pericardial cavity to contact (arrowhead) the myocardium (M). (c) At points of contact with the myocardium, the PE epithelial sheet breaks open (arrowheads) to preserve apical-basal polarity and migrate over the heart tube (arrows). (d) During migration and after a continuous epicardium (Ep) is formed, cells leave the epicardium (arrowheads) via epithelial to mesenchymal transition (EMT) to migrate into the myocardium and generate fibroblasts (F) and the coronary blood vessels (BV). EC, endocardium; G, gut tube; H, heart tube.
It is of interest to note that when the PE can first be identified, its epithelial component expresses gene products characteristic of a mesothelium including Tbx18, Wt1, and Tcf21 [
33,
34,
35]. Thus, before these cells come in contact with their target organ the heart, they appear to have already differentiated, or at least partially so, into a mesothelium.
As the epicardium advances over the developing myocardium, the process of EMT begins. EMT is well underway before the epicardium fully covers the heart and several laboratories have demonstrated the propensity of the epicardium to form mesenchymal cells [
8,
33,
36,
37]. The distribution of epicardially-derived mesenchyme is not even throughout the heart with the bulk observed at the atrioventricular junction where major vessels will develop at subsequent stages. Other myocardial regions are not so heavily populated by EMT-derived cells [
8].
The identity of specific EMT-derived progeny is firmly established while that of others is currently in debate. Direct labeling of the PE with vital dyes and retroviral lineage tracers in avian embryos has demonstrated that precursors of the epicardium, vascular endothelial and smooth muscle cells as well as pericytes and fibroblasts are housed within the PE [
32,
38,
39]. Conversely, ablation of the PE results in loss of the coronary vasculature [
40]. Previous studies tracing the Wt1 and Tbx18 proepicardial lineages suggested that endothelial cells may not be the descendants of the epithelial PE but potentially arise from its mesenchymal component [
34,
35]. A more recent study determined that endothelial cells are indeed derived from specific compartments of the PE that express
Scleraxis and
Semaphorin3D [
41]. Taken together, studies on the PE/epicardium suggest that these simple squamous epithelial structures give rise to the vast majority of cells comprising the coronary vessels and cardiac fibroblasts.
Another important aspect to generation of the coronary arterial system is the growth and maturation of the proximal portion of these vessels. Manner demonstrated that the proximal ends of coronary vessels move within the epicardial space toward the aorta [
42]. These vessels penetrate the tunica media of the aorta and through apoptotic events around the future coronary orifice establish continuity with the aorta and systemic circulation [
43]. Prior to the establishment of the final coronary arterial pathway, intermittent passage of blood is observed through transient patency of coronary sprouts with the aorta [
44]. This mechanism of vasculogenesis in developing organs has only been reported for the heart.
To summate, the heart, which is housed within the pericardial cavity is initially devoid of a mesothelium and blood vessels. The PE migrates to and over the heart tube with a concurrent EMT-driven production of freely migratory cells. These cells differentiate into vascular endothelium and smooth muscle as well as cardiac fibroblasts and possibly cardiac myocytes. Thus, the PE is the major source of cells for the developing coronary system as well as other fibroblastic and parenchymal cell types in the entire organ.
New and interesting studies will be published on the regulation of PE/epicardial control of cardiac development but it appears that the major roadmap of development is understood. An intriguing question arises, “Is this developmental process conserved in the formation and development of other coelomic organs?” In contemplating this question, we became particularly interested in the GI tract due to the similar structural arrangement of the intestinal and cardiac vasculature, as described above.
7. Development of the Serosal Mesothelium
As noted earlier, the alimentary canal arises from two embryonic structures: endoderm and splanchnic mesoderm. Later, neural crest cells invade and contribute to elements of the enteric plexus [
45]. With lateral folding, a complete tube is suspended from the dorsal body wall by a dorsal mesentery not unlike the early heart. The epithelial endoderm is maintained as the GI mucosal epithelium but also buds into the surrounding mesoderm in highly regulated processes that produce the epithelial components of glands such as the liver and pancreas. The epithelial connections between the intestinal mucosa and the glandular epithelium of these exocrine organs via continuous ductal systems are maintained throughout life. Concurrent with this epithelial morphogenesis, cells of the splanchnic mesoderm undergo complex remodeling to produce connective tissue, visceral smooth muscle, and vascular components of the gastrointestinal wall and glands. Excellent reviews examine the signaling and genetic control of this process [
46,
47]. For purposes of the current discussion, it is important to note that stromal elements of the lamina propria and submucosa along with those same constituents in associated glands are thought to be progeny of the splanchnic mesoderm. Further visceral smooth muscle of the muscularis externa is also descendent from splanchnic mesoderm. As noted above, neural crest cells migrate into the GI tract to produce neurons and glia of the enteric plexus [
45].
In 1980 Meier answered a fundamental question, “When is the mesothelium first seen in gut development?” He demonstrated that splanchnic mesoderm was initially devoid of a histologically identifiable mesothelium. We later demonstrated that the gut tube did not initially express molecular markers (
Wt1, cytokeratin) of the mesothelial phenotype [
27,
48]. It was interesting to note that in the mouse, expression of
Wt1, a widely used marker for mesothelial cells, was first seen at the root of the dorsal mesentery of the intestine and subsequently spread ventrally over the gut tube. We postulated that a wave of migratory cells, like those of the PE swept down upon the developing gut tube though it should be noted this has never been tested.
In a recently published communication, we detailed the formation of mesothelium on the gut wall in the avian embryo [
26]. Using the quail embryo as a developmental model, we confirmed Meier’s finding that splanchnic mesoderm is devoid of mesothelium. With the split of lateral mesoderm to form the coelom, the gut organ primordium is composed solely of endoderm, splanchnic mesoderm and the aforementioned vascular plexus which is situated between their basement membranes (
Figure 6A–B). Our present data suggest that cells of the splanchnic mesoderm lose their epithelial nature and move into the space between juxtaposed basement membranes forming a primary gut mesenchyme (
Figure 6C). This mesenchyme will give rise to cellular components of the lamina propria, submucosa, and muscularis externa. All these cells are non-epithelial and are grouped as “connective tissue/stromal/smooth muscle” cells. The inner endodermally-associated basement membrane is definitively retained throughout development. The outer mesodermally-associated basement membrane, while always present, is temporally fragmented during the course of organogenesis (
Figure 6C). We emphasize the positioning and constancy of these two basement membranes as they represent important landmarks for an understanding of intestinal and, specifically, mesothelial development. These are the only two continuous basement membranes in the entirety of the developing and adult gut tube. The inner basement membrane is associated with the mucosal epithelium and with its precursor embryonic endoderm. The outer basement membrane is originally associated with splanchnic mesoderm but where is its position in the adult and where is the mesothelium in relationship to this important landmark?
The outer epithelium is highly dynamic during this initial timeframe. The epithelium itself changes shape and thickness with many cells moving from the surface into the mesenchymal space between the two basement membranes. At these times, this outer epithelium ranges from columnar to pseudostratified to stratified but is clearly not simple squamous in appearance. Only at embryonic day 6 of chick or quail development is definitive mesothelium seen on the forming gut tube [
26,
27]. After this time and extending throughout adulthood, the simple squamous mesothelium is the only cell type seen external to the outer basement membrane. Once the mesothelium has been established, we have shown in the mouse that EMT from this epithelium ensues [
48]. With time, a connective tissue space is established subjacent to the basement membrane with an embedded vasculature. This is the same arrangement observed in the heart with its epicardium. Thus, at the end of development, the surface of the heart and GI tract have the same arrangement of mesothelium and underlying connective tissue with a vascular supply. But, is there a “gut PE”?
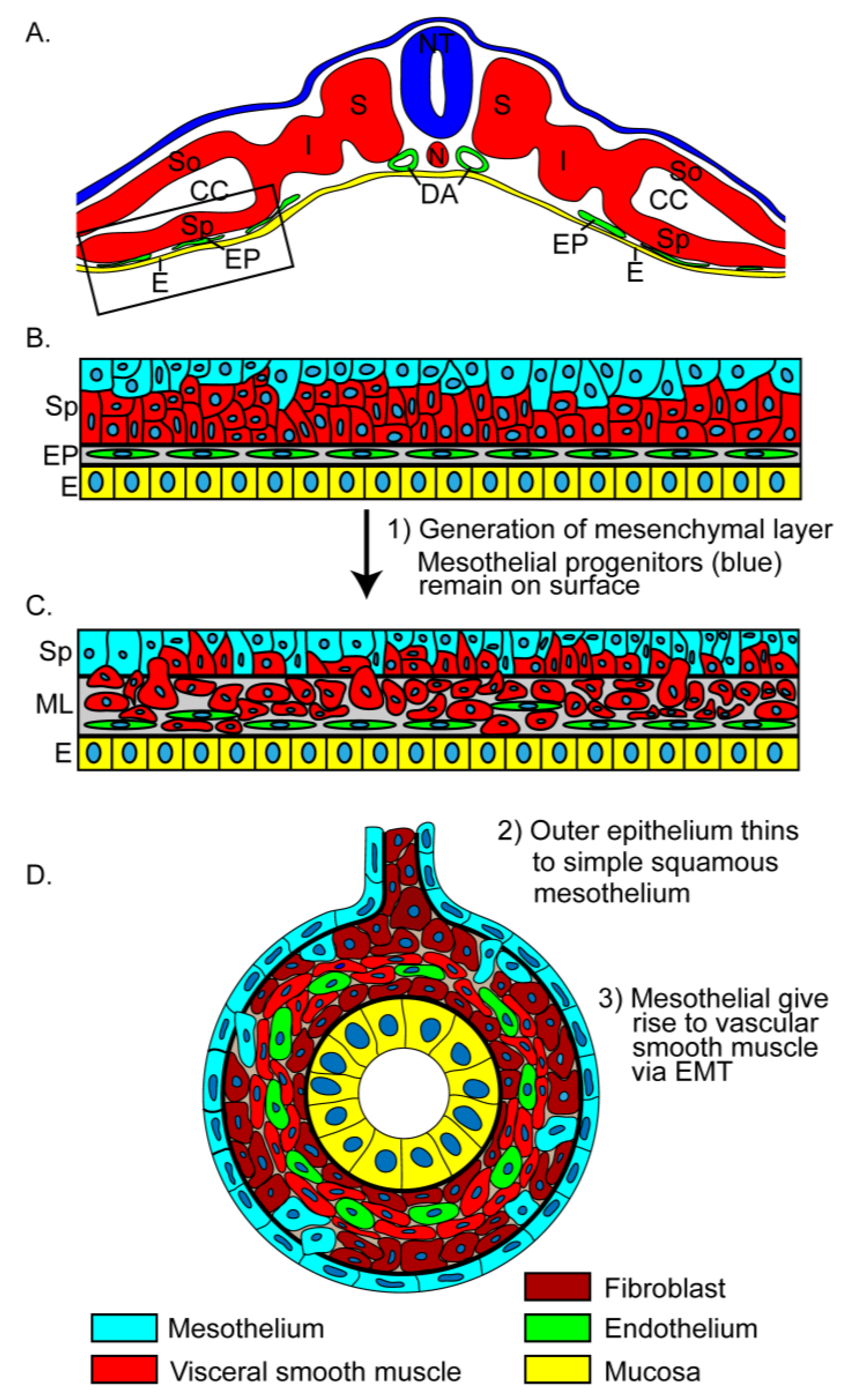
Figure 6.
Intestinal mesothelial development. (a) Schematic depicting an early embryo yet to undergo lateral folding. (b) The splanchnopleure of the intestinal anlage (boxed area in A) is composed of splanchnic mesoderm (Sp) and endoderm (E) with an intervening endothelial plexus (EP). The mesothelial progenitors (blue) are distributed primarily near the surface of the stratified splanchnic mesoderm while fibroblast and visceral smooth muscle progenitors are deep (red). (c) Cells of the splanchnic mesoderm undergo EMT and invade the mesenchymal space joining the endothelial plexus to form the mesenchymal layer (ML). The mesodermal basement membrane breaks down through this process (broken black line). (d) A simple squamous mesothelium (blue) is present on the surface by day 6 of chick development and E11.5 of mouse development. Mesothelial cells undergo EMT to give rise to vascular smooth muscle. CC, coelomic cavity; DA, dorsal aorta; E, endoderm; I, intermediate mesoderm; N, notochord; NT, neural tube; S, somite.
Figure 6.
Intestinal mesothelial development. (a) Schematic depicting an early embryo yet to undergo lateral folding. (b) The splanchnopleure of the intestinal anlage (boxed area in A) is composed of splanchnic mesoderm (Sp) and endoderm (E) with an intervening endothelial plexus (EP). The mesothelial progenitors (blue) are distributed primarily near the surface of the stratified splanchnic mesoderm while fibroblast and visceral smooth muscle progenitors are deep (red). (c) Cells of the splanchnic mesoderm undergo EMT and invade the mesenchymal space joining the endothelial plexus to form the mesenchymal layer (ML). The mesodermal basement membrane breaks down through this process (broken black line). (d) A simple squamous mesothelium (blue) is present on the surface by day 6 of chick development and E11.5 of mouse development. Mesothelial cells undergo EMT to give rise to vascular smooth muscle. CC, coelomic cavity; DA, dorsal aorta; E, endoderm; I, intermediate mesoderm; N, notochord; NT, neural tube; S, somite.
![Jdb 01 00064 g006]()
To determine whether gut mesothelium arises from an exogenous source like that seen in the heart, we employed a chick/quail chimeric system where segments of quail splanchnopleure (endoderm plus splanchnic mesoderm) were transplanted into a host chick coelomic cavity. Interestingly, mesothelium was generated from the graft tissue and not the host, meaning that mesothelial progenitors were broadly positioned and intrinsic to the organ primordium, and that a “gut PE” does not account for these cells [
27]. This was substantiated by DNA electroporation and long term retroviral experiments labeling surface cells of splanchnic mesoderm. Taken together, our recently published data suggest that the mesothelium of the avian gut tube is derived by a mechanism completely different from that seen in the heart.
10. Lineages Generated From Epicardial and Serosal Mesothelia
Numerous studies have analyzed the cell types generated by the PE/epicardium. In studies of heart organogenesis, these cells generated through EMT include vascular smooth muscle, fibroblasts, myofibroblasts, endothelium, and possibly cardiac myocytes, as well as the epicardium itself (see above and other articles in this issue). Lineage tracing studies of proepicardial cells have been conducted using direct retroviral labeling as well as genetic techniques. Results vary with these two methodologies as one labels a “location” while the other labels an “expressing cell”. Results from the two labeling strategies demonstrate the presence of both endothelial and vascular smooth muscle progenitors within the PE [
32,
38,
39,
40,
41,
48,
53,
54] although positioning of these progenitors within the PE is still a matter of question.
In reflecting on the differentiation of mesothelial cells in the developing gut tube, the aforementioned types of experiment are difficult to interpret in the context of gut organogenesis. First, no one has proven that gut mesothelium is the only population to express the currently available mesothelial genetic markers (such as
Wt1,
Tbx18, or
Gata4). In Wilm
et al [
48], we demonstrated that the
Wt1 genetic lineage gives rise to the majority of vascular smooth muscle of the intestines. However, our recent data suggest that
Wt1, an often used marker, is expressed in splanchnic mesoderm prior to generation of a definitive mesothelium [
27]. This is also observed in a recent study genetically tracing a mesothelin-positive lineage throughout the coelomic cavity in the mouse. This particular gene, though associated with differentiated mesothelium, was demonstrated to also label a progenitor population, termed Fibroblast and Smooth Muscle Cell (FSMC) precursors, that embryonically gives rise not only to mesothelium but also the vast majority of visceral smooth muscle of the intestines and bladder [
55]. Going forward, it will be important to carefully characterize the expression of “mesothelial-specific” genes before standard Cre tracing systems can be used to study mesothelially-derived cell lineages in the gut tube, lungs, and associated glands. This would include any work from our own laboratory. Retroviral, vital dye, or DNA electroporation-based labeling of cells at or near the surface of intestinal splanchnic mesoderm marks definitive mesothelium as well as vascular smooth muscle and stromal cells within the connective tissue if the embryo is allowed to grow [
27]. At the time of labeling in these experiments, the splanchnic mesoderm is devoid of a definitive mesothelial cell layer and is characterized by stratified and pseudostratified epithelium. Thus, while these experiments definitively demonstrate that mesothelial progenitors reside within the gut primordium identifying labeled cells as strict descendants of the gut mesothelium is problematic.
Fundamental differences in mesothelial development in the heart and gut account for the difficulty in interpretation of mesothelial descendants in the developing GI tract. First, the mesothelial precursor of the heart, the PE, has characteristics of a mesothelium before it arrives at its target organ. Thus, expression analysis of genes to be used for lineage-tracing can be more easily conducted. Still, even now, controversy in data interpretation has been experienced. Conversely, precursors of gut mesothelium arise broadly within the developing organ and are heterogeneously dispersed or mixed within a population of cells with differing potentials. Thus going forward, we would submit that experimental models examining mesothelially-derived cell lineages in the gut tube explore timeframes when only definitive mesothelial cells can be identified and labeled. This would entail determining when mesothelial-specific genes are expressed only in the serosal mesothelium and activating Cre expression at that time. Viral- and vital-labeling would again be conducted at those time points. Indeed, our own study [
48] should be revisited using temporally-activated Cre expression to ensure that only mesothelial progeny are being labeled. This strategy, in strictest terms, might be effective in identifying descendants of visceral mesothelia.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





