
Pigment Epithelium-Derived Factor (PEDF) is a Determinant of Stem Cell Fate: Lessons from an Ultra-Rare Disease

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Osteogenesis Imperfecta Type VI
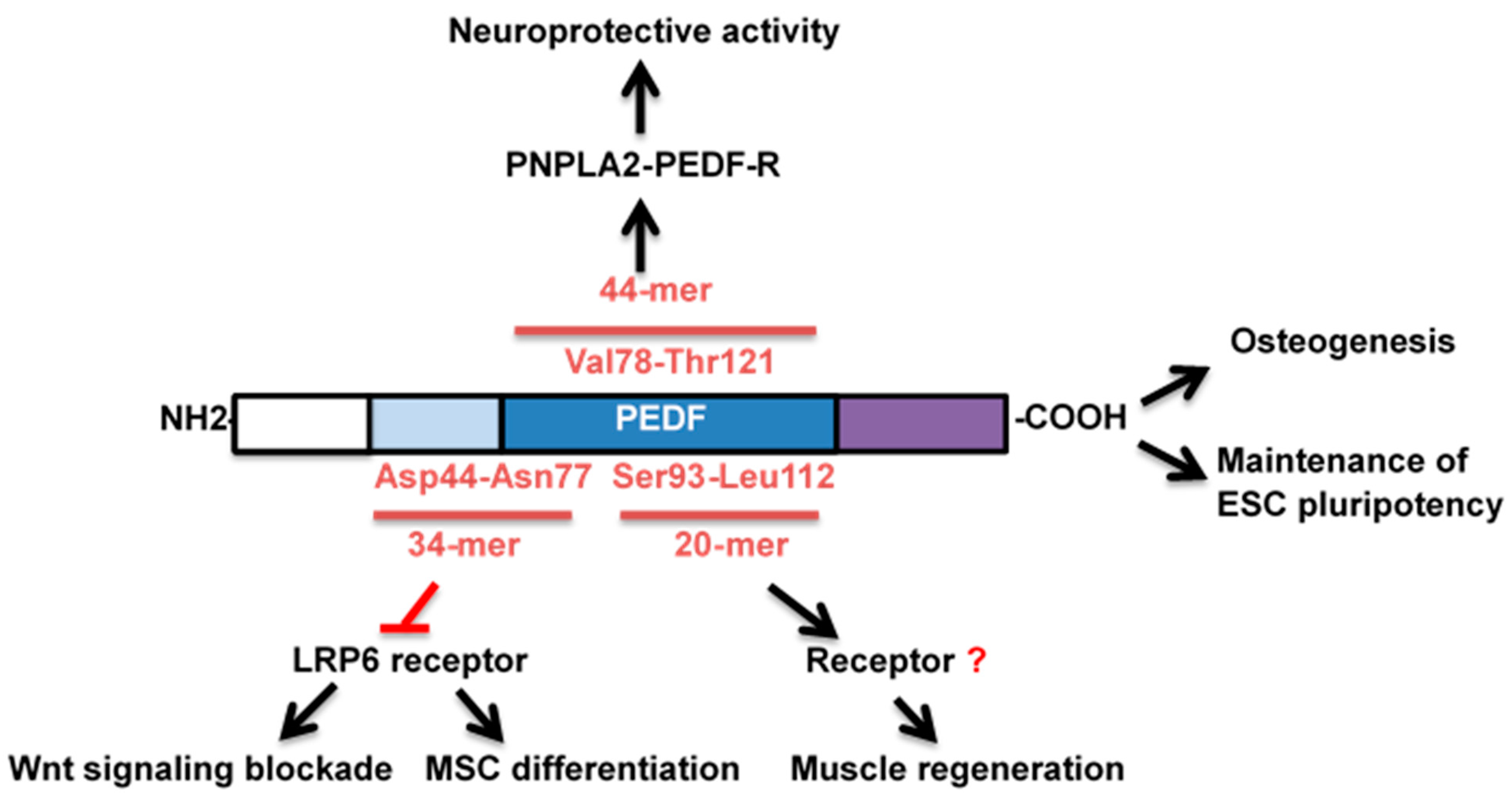
3. Historical Perspectives & PEDF Signaling

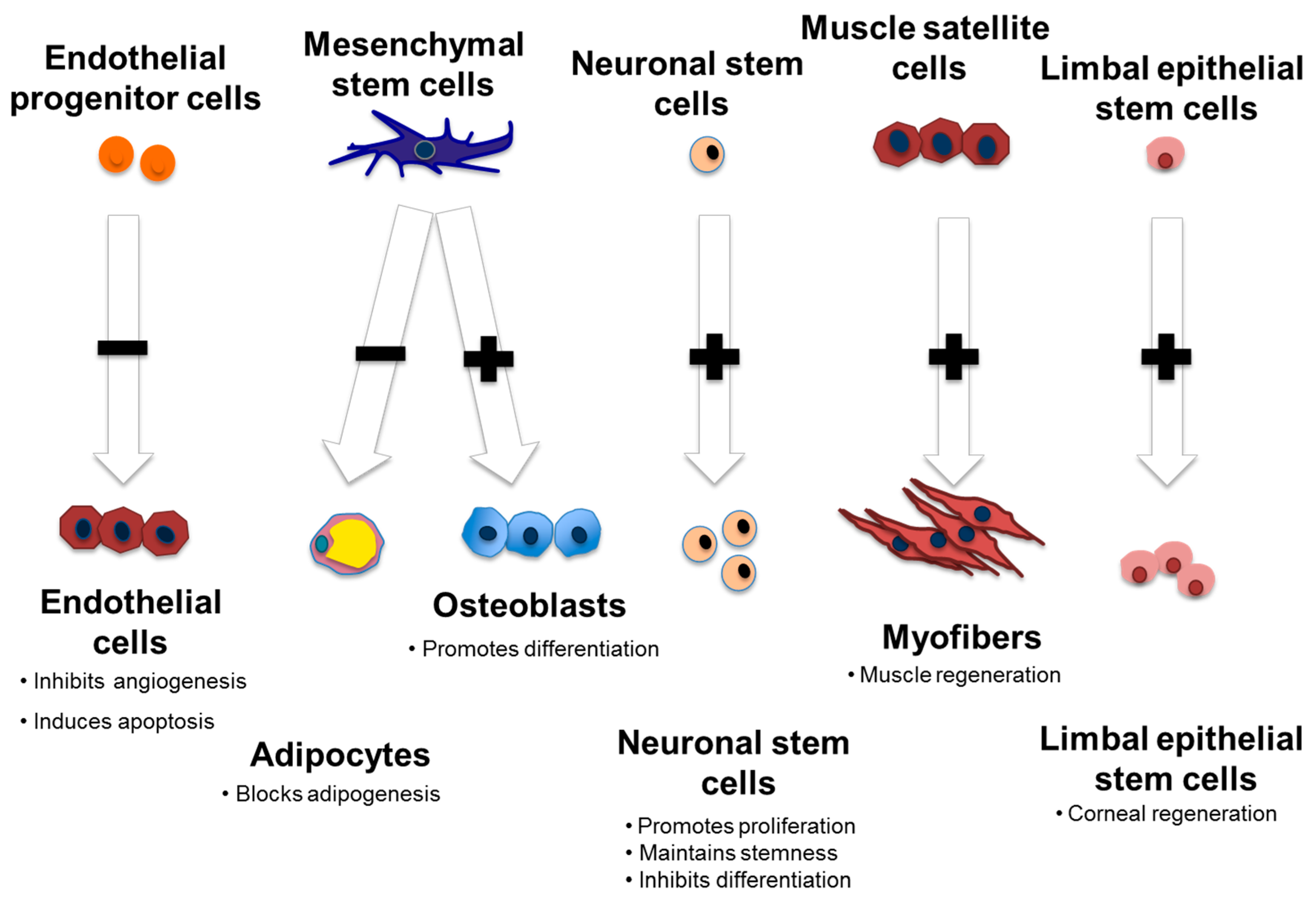
4. Stem Cells

4.1. Musculoskeletal System
4.2. Neurotrophic Effects
4.3. Ocular Development
5. Future Directions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tombran-Tink, J.; Johnson, L.V. Neuronal differentiation of retinoblastoma cells induced by medium conditioned by human rpe cells. Investig. Ophthalmol. Vis. Sci. 1989, 30, 1700–1707. [Google Scholar]
- Steele, F.R.; Chader, G.J.; Johnson, L.V.; Tombran-Tink, J. Pigment epithelium-derived factor: Neurotrophic activity and identification as a member of the serine protease inhibitor gene family. Proc. Natl. Acad. Sci. USA 1993, 90, 1526–1530. [Google Scholar] [CrossRef] [PubMed]
- Becerra, S.P.; Sagasti, A.; Spinella, P.; Notario, V. Pigment epithelium-derived factor behaves like a noninhibitory serpin. Neurotrophic activity does not require the serpin reactive loop. J. Biol. Chem. 1995, 270, 25992–25999. [Google Scholar] [CrossRef] [PubMed]
- Tombran-Tink, J.; Mazuruk, K.; Rodriguez, I.R.; Chung, D.; Linker, T.; Englander, E.; Chader, G.J. Organization, evolutionary conservation, expression and unusual Alu density of the human gene for pigment epithelium-derived factor, a unique neurotrophic serpin. Mol. Vis. 1996, 2, 11. [Google Scholar] [PubMed]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347. [Google Scholar] [CrossRef] [PubMed]
- Tombran-Tink, J.; Barnstable, C.J. Osteoblasts and osteoclasts express PEDF, VEGF-A isoforms, and VEGF receptors: Possible mediators of angiogenesis and matrix remodeling in the bone. Biochem. Biophys. Res. Commun. 2004, 316, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Petersen, S.V.; Valnickova, Z.; Enghild, J.J. Pigment-epithelium-derived factor (PEDF) occurs at a physiologically relevant concentration in human blood: Purification and characterization. Biochem. J. 2003, 374, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, S.; Adachi, H.; Abe, A.; Yashiro, T.; Enomoto, M.; Furuki, K.; Hino, A.; Jinnouchi, Y.; Takenaka, K.; Matsui, T.; et al. Elevated serum levels of pigment epithelium-derived factor in the metabolic syndrome. J. Clin. Endocrinol. Metab. 2006, 91, 2447–2450. [Google Scholar] [CrossRef] [PubMed]
- Rauch, F.; Husseini, A.; Roughley, P.; Glorieux, F.H.; Moffatt, P. Lack of circulating pigment epithelium-derived factor is a marker of osteogenesis imperfecta type VI. J. Clin. Endocrinol. Metab. 2012, 97, E1550–E1556. [Google Scholar] [CrossRef] [PubMed]
- Glorieux, F.H.; Ward, L.M.; Rauch, F.; Lalic, L.; Roughley, P.J.; Travers, R. Osteogenesis imperfecta type VI: A form of brittle bone disease with a mineralization defect. J. Bone Miner. Res. 2002, 17, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Roughley, P.J.; Rauch, F.; Glorieux, F.H. Osteogenesis imperfecta—Clinical and molecular diversity. Eur. Cells Mater. 2003, 5, 41–47; discussion 47. [Google Scholar]
- Fratzl-Zelman, N.; Schmidt, I.; Roschger, P.; Roschger, A.; Glorieux, F.H.; Klaushofer, K.; Wagermaier, W.; Rauch, F.; Fratzl, P. Unique micro- and nano-scale mineralization pattern of human osteogenesis imperfecta type VI bone. Bone 2015, 73, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Bogan, R.; Riddle, R.C.; Li, Z.; Kumar, S.; Nandal, A.; Faugere, M.C.; Boskey, A.; Crawford, S.E.; Clemens, T.L. A mouse model for human osteogenesis imperfecta type VI. J. Bone Miner. Res. 2013, 28, 1531–1536. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.; Semler, O.; Gilissen, C.; Li, Y.; Bolz, H.J.; Giunta, C.; Bergmann, C.; Rohrbach, M.; Koerber, F.; Zimmermann, K.; et al. Exome sequencing identifies truncating mutations in human serpinf1 in autosomal-recessive osteogenesis imperfecta. Am. J. Hum. Genet. 2011, 88, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Homan, E.P.; Rauch, F.; Grafe, I.; Lietman, C.; Doll, J.A.; Dawson, B.; Bertin, T.; Napierala, D.; Morello, R.; Gibbs, R.; et al. Mutations in serpinf1 cause osteogenesis imperfecta type VI. J. Bone Miner. Res. 2011, 26, 2798–2803. [Google Scholar] [CrossRef] [PubMed]
- Venturi, G.; Gandini, A.; Monti, E.; dalle Carbonare, L.; Corradi, M.; Vincenzi, M.; Valenti, M.T.; Valli, M.; Pelilli, E.; Boner, A.; et al. Lack of expression of serpinf1, the gene coding for pigment epithelium-derived factor, causes progressively deforming osteogenesis imperfecta with normal type I collagen. J. Bone Miner. Res. 2012, 27, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Al-Jallad, H.; Palomo, T.; Roughley, P.; Glorieux, F.H.; McKee, M.D.; Moffatt, P.; Rauch, F. The effect of SERPINF1 in-frame mutations in osteogenesis imperfecta type VI. Bone 2015, 76, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Bryant, H.U.; Macdougald, O.A. Regulation of bone mass by Wnt signaling. J. Clin. Invest. 2006, 116, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Gattu, A.K.; Swenson, E.S.; Iwakiri, Y.; Samuel, V.T.; Troiano, N.; Berry, R.; Church, C.D.; Rodeheffer, M.S.; Carpenter, T.O.; Chung, C. Determination of mesenchymal stem cell fate by pigment epithelium-derived factor (PEDF) results in increased adiposity and reduced bone mineral content. FASEB J. 2013, 27, 4384–4394. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Song, N.; Tombran-Tink, J.; Niyibizi, C. Pigment epithelium-derived factor enhances differentiation and mineral deposition of human mesenchymal stem cells. Stem Cells 2013, 31, 2714–2723. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Song, N.; Tombran-Tink, J.; Niyibizi, C. Pigment epithelium derived factor suppresses expression of Sost/Sclerostin by osteocytes: Implication for its role in bone matrix mineralization. J. Cell. Physiol. 2015, 230, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Lee, K.; Zhang, B.; Zhou, T.; He, X.; Gao, G.; Murray, A.R.; Ma, J.X. Identification of a novel inhibitor of the canonical Wnt pathway. Mol. Cell. Biol. 2011, 31, 3038–3051. [Google Scholar] [CrossRef] [PubMed]
- Protiva, P.; Gong, J.; Sreekumar, B.; Torres, R.; Zhang, X.; Belinsky, G.S.; Cornwell, M.; Crawford, S.E.; Iwakiri, Y.; Chung, C. Pigment epithelium-derived factor (PEDF) inhibits Wnt/B-catenin signaling in the liver. Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Yang, C.; Dai, Z.; Che, D.; Feng, J.; Mao, Y.; Cheng, R.; Wang, Z.; He, X.; Zhou, T.; et al. High levels of pigment epithelium-derived factor in diabetes impair wound healing through suppression of Wnt signaling. Diabetes 2015, 64, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.S.; Sorenson, C.M.; Sheibani, N. PEDF expression regulates the proangiogenic and proinflammatory phenotype of the lung endothelium. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, L620–L634. [Google Scholar] [CrossRef] [PubMed]
- Becerra, S.P.; Palmer, I.; Kumar, A.; Steele, F.; Shiloach, J.; Notario, V.; Chader, G.J. Overexpression of fetal human pigment epithelium-derived factor in escherichia coli. A functionally active neurotrophic factor. J. Biol. Chem. 1993, 268, 23148–23156. [Google Scholar] [PubMed]
- Tombran-Tink, J.; Chader, G.G.; Johnson, L.V. PEDF: A pigment epithelium-derived factor with potent neuronal differentiative activity. Exp. Eye Res. 1991, 53, 411–414. [Google Scholar] [CrossRef]
- Seigel, G.M.; Tombran-Tink, J.; Becerra, S.P.; Chader, G.J.; Diloreto, D.A., Jr.; del Cerro, C.; Lazar, E.S.; del Cerro, M. Differentiation of Y79 retinoblastoma cells with pigment epithelial-derived factor and interphotoreceptor matrix wash: Effects on tumorigenicity. Growth Factors 1994, 10, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Castillejo, C.; Sanchez-Sanchez, F.; Andreu-Agullo, C.; Ferron, S.R.; Aroca-Aguilar, J.D.; Sanchez, P.; Mira, H.; Escribano, J.; Farinas, I. Pigment epithelium-derived factor is a niche signal for neural stem cell renewal. Nat. Neurosci. 2006, 9, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Bilak, M.M.; Becerra, S.P.; Vincent, A.M.; Moss, B.H.; Aymerich, M.S.; Kuncl, R.W. Identification of the neuroprotective molecular region of pigment epithelium-derived factor and its binding sites on motor neurons. J. Neurosci. 2002, 22, 9378–9386. [Google Scholar] [PubMed]
- DeCoster, M.A.; Schabelman, E.; Tombran-Tink, J.; Bazan, N.G. Neuroprotection by pigment epithelial-derived factor against glutamate toxicity in developing primary hippocampal neurons. J. Neurosci. Res. 1999, 56, 604–610. [Google Scholar] [CrossRef]
- Taniwaki, T.; Becerra, S.P.; Chader, G.J.; Schwartz, J.P. Pigment epithelium-derived factor is a survival factor for cerebellar granule cells in culture. J. Neurochem. 1995, 64, 2509–2517. [Google Scholar] [CrossRef] [PubMed]
- Taniwaki, T.; Hirashima, N.; Becerra, S.P.; Chader, G.J.; Etcheberrigaray, R.; Schwartz, J.P. Pigment epithelium-derived factor protects cultured cerebellar granule cells against glutamate-induced neurotoxicity. J. Neurochem. 1997, 68, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Filleur, S.; Volz, K.; Nelius, T.; Mirochnik, Y.; Huang, H.; Zaichuk, T.A.; Aymerich, M.S.; Becerra, S.P.; Yap, R.; Veliceasa, D.; et al. Two functional epitopes of pigment epithelial-derived factor block angiogenesis and induce differentiation in prostate cancer. Cancer Res. 2005, 65, 5144–5152. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.I.; Ho, T.C.; Chen, S.L.; Chen, C.P.; Cheng, H.C.; Lan, Y.W.; Hsieh, J.W.; Wang, C.T.; Tsao, Y.P. Pigment epithelial-derived factor peptide facilitates the regeneration of a functional limbus in rabbit partial limbal deficiency. Investig. Ophthalmol. Visual Sci. 2015, 56, 2126–2134. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.C.; Chiang, Y.P.; Chuang, C.K.; Chen, S.L.; Hsieh, J.W.; Lan, Y.W.; Tsao, Y.P. PEDF-derived peptide promotes skeletal muscle regeneration through its mitogenic effect on muscle progenitor cells. Am. J. Physiol. Cell Physiol. 2015, 309, C159–C168. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.H.; Shih, S.C.; Ho, T.C.; Ma, H.I.; Liu, M.Y.; Chen, S.L.; Tsao, Y.P. Pigment epithelium-derived factor 34-mer peptide prevents liver fibrosis and hepatic stellate cell activation through down-regulation of the PDGF receptor. PLoS ONE 2014, 9, e95443. [Google Scholar] [CrossRef] [PubMed]
- Alberdi, E.; Aymerich, M.S.; Becerra, S.P. Binding of pigment epithelium-derived factor (PEDF) to retinoblastoma cells and cerebellar granule neurons. Evidence for a PEDF receptor. J. Biol. Chem. 1999, 274, 31605–31612. [Google Scholar] [CrossRef] [PubMed]
- Notari, L.; Baladron, V.; Aroca-Aguilar, J.D.; Balko, N.; Heredia, R.; Meyer, C.; Notario, P.M.; Saravanamuthu, S.; Nueda, M.L.; Sanchez-Sanchez, F.; et al. Identification of a lipase-linked cell membrane receptor for pigment epithelium-derived factor. J. Biol. Chem. 2006, 281, 38022–38037. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Gao-Li, J.; Franco, C.A.; Bouceba, T.; Huet, A.; Li, Z. Laminin receptor involvement in the anti-angiogenic activity of pigment epithelium-derived factor. J. Biol. Chem. 2009, 284, 10480–10490. [Google Scholar] [CrossRef] [PubMed]
- Notari, L.; Arakaki, N.; Mueller, D.; Meier, S.; Amaral, J.; Becerra, S.P. Pigment epithelium-derived factor binds to cell-surface F(1)-ATP synthase. FEBS J. 2010, 277, 2192–2205. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.; Notari, L.; Becerra, S.P. Mapping the type I collagen-binding site on pigment epithelium-derived factor. Implications for its antiangiogenic activity. J. Biol. Chem. 2002, 277, 45400–45407. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R. Wnt signaling. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Wietecha, M.S.; Krol, M.J.; Michalczyk, E.R.; Chen, L.; Gettins, P.G.; DiPietro, L.A. Pigment epithelium-derived factor as a multifunctional regulator of wound healing. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H812–H826. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, J.C.; Protiva, P.; Gattu, A.K.; Utsumi, T.; Iwakiri, Y.; Neto, A.G.; Quinn, M.; Cornwell, M.L.; Fitchev, P.; Lugea, A.; et al. Pigment epithelium-derived factor regulates early pancreatic fibrotic responses and suppresses the profibrotic cytokine thrombospondin-1. Am. J. Pathol. 2011, 179, 2990–2999. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.C.; Chen, S.L.; Shih, S.C.; Wu, J.Y.; Han, W.H.; Cheng, H.C.; Yang, S.L.; Tsao, Y.P. Pigment epithelium-derived factor is an intrinsic antifibrosis factor targeting hepatic stellate cells. Am. J. Pathol. 2010, 177, 1798–1811. [Google Scholar] [CrossRef] [PubMed]
- Mejias, M.; Coch, L.; Berzigotti, A.; Garcia-Pras, E.; Gallego, J.; Bosch, J.; Fernandez, M. Antiangiogenic and antifibrogenic activity of pigment epithelium-derived factor (PEDF) in bile duct-ligated portal hypertensive rats. Gut 2015, 64, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Gattu, A.K.; Birkenfeld, A.L.; Iwakiri, Y.; Jay, S.; Saltzman, M.; Doll, J.; Protiva, P.; Samuel, V.T.; Crawford, S.E.; Chung, C. Pigment epithelium-derived factor (PEDF) suppresses IL-1β-mediated c-jun N-terminal kinase (JNK) activation to improve hepatocyte insulin signaling. Endocrinology 2014, 155, 1373–1385. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fergusson, M.M.; Wu, J.J.; Rovira, I.I.; Liu, J.; Gavrilova, O.; Lu, T.; Bao, J.; Han, D.; Sack, M.N.; et al. Wnt signaling regulates hepatic metabolism. Sci. Signal. 2011, 4, ra6. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Singh, R.; Choi, C.S.; Lee, H.Y.; Keramati, A.R.; Samuel, V.T.; Lifton, R.P.; Shulman, G.I.; Mani, A. Low density lipoprotein (LDL) receptor-related protein 6 (LRP6) regulates body fat and glucose homeostasis by modulating nutrient sensing pathways and mitochondrial energy expenditure. J. Biol. Chem. 2012, 287, 7213–7223. [Google Scholar] [CrossRef] [PubMed]
- Grippo, P.J.; Fitchev, P.S.; Bentrem, D.J.; Melstrom, L.G.; Dangi-Garimella, S.; Krantz, S.B.; Heiferman, M.J.; Chung, C.; Adrian, K.; Cornwell, M.L.; et al. Concurrent PEDF deficiency and kras mutation induce invasive pancreatic cancer and adipose-rich stroma in mice. Gut 2012, 61, 1454–1464. [Google Scholar] [CrossRef] [PubMed]
- Jaenisch, R.; Young, R. Stem cells, the molecular circuitry of pluripotency and nuclear reprogramming. Cell 2008, 132, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.J.; Kaufman, M.H. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981, 292, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef] [PubMed]
- Miki, T.; Ring, A.; Gerlach, J. Hepatic differentiation of human embryonic stem cells is promoted by three-dimensional dynamic perfusion culture conditions. Tissue Eng. Part C Methods 2011, 17, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Anisimov, S.V.; Christophersen, N.S.; Correia, A.S.; Hall, V.J.; Sandelin, I.; Li, J.Y.; Brundin, P. Identification of molecules derived from human fibroblast feeder cells that support the proliferation of human embryonic stem cells. Cell. Mol. Biol. Lett. 2011, 16, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Jennings, L.L.; Knuth, M.; Orth, A.P.; Klock, H.E.; Ou, W.; Feuerhelm, J.; Hull, M.V.; Koesema, E.; Wang, Y.; et al. Screening the mammalian extracellular proteome for regulators of embryonic human stem cell pluripotency. Proc. Natl. Acad. Sci. USA 2010, 107, 3552–3557. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.; Pang, C.P.; Yam, H.F.; Cheung, K.F.; Liu, W.W.; Lu, Y. Inhibition of glioma invasion by overexpression of pigment epithelium-derived factor. Cancer Gene Ther. 2004, 11, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Prowse, A.B.; McQuade, L.R.; Bryant, K.J.; Marcal, H.; Gray, P.P. Identification of potential pluripotency determinants for human embryonic stem cells following proteomic analysis of human and mouse fibroblast conditioned media. J. Proteome Res. 2007, 6, 3796–3807. [Google Scholar] [CrossRef] [PubMed]
- Kanemura, H.; Go, M.J.; Nishishita, N.; Sakai, N.; Kamao, H.; Sato, Y.; Takahashi, M.; Kawamata, S. Pigment epithelium-derived factor secreted from retinal pigment epithelium facilitates apoptotic cell death of iPSC. Sci. Rep. 2013, 3, 2334. [Google Scholar] [CrossRef] [PubMed]
- Prockop, D.J.; Olson, S.D. Clinical trials with adult stem/progenitor cells for tissue repair: Let’s not overlook some essential precautions. Blood 2007, 109, 3147–3151. [Google Scholar] [CrossRef] [PubMed]
- Dor, Y.; Melton, D.A. How important are adult stem cells for tissue maintenance? Cell Cycle 2004, 3, 1104–1106. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Guan, J.; Zhang, C. Mesenchymal stem cells: Mechanisms and role in bone regeneration. Postgrad. Med. J. 2014, 90, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Sarojini, H.; Estrada, R.; Lu, H.; Dekova, S.; Lee, M.J.; Gray, R.D.; Wang, E. PEDF from mouse mesenchymal stem cell secretome attracts fibroblasts. J. Cell. Biochem. 2008, 104, 1793–1802. [Google Scholar] [CrossRef] [PubMed]
- Baron, R.; Kneissel, M. Wnt signaling in bone homeostasis and disease: From human mutations to treatments. Nat. Med. 2013, 19, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Farmer, S.R. Regulating the balance between peroxisome proliferator-activated receptor gamma and beta-catenin signaling during adipogenesis. A glycogen synthase kinase 3beta phosphorylation-defective mutant of beta-catenin inhibits expression of a subset of adipogenic genes. J. Biol. Chem. 2004, 279, 45020–45027. [Google Scholar] [PubMed]
- Ross, S.E.; Hemati, N.; Longo, K.A.; Bennett, C.N.; Lucas, P.C.; Erickson, R.L.; MacDougald, O.A. Inhibition of adipogenesis by Wnt signaling. Science 2000, 289, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Cruciat, C.M.; Niehrs, C. Secreted and transmembrane wnt inhibitors and activators. Cold Spring Harb. Perspect. Biol. 2013, 5, a015081. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Niehrs, C. Kremen2 modulates Dickkopf2 activity during Wnt/LRP6 signaling. Gene 2003, 302, 179–183. [Google Scholar] [CrossRef]
- Quan, G.M.; Ojaimi, J.; Li, Y.; Kartsogiannis, V.; Zhou, H.; Choong, P.F. Localization of pigment epithelium-derived factor in growing mouse bone. Calcif. Tissue Int. 2005, 76, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Garzon-Muvdi, T.; Quinones-Hinojosa, A. Neural stem cell niches and homing: Recruitment and integration into functional tissues. ILAR J. Natl. Res. Counc. Inst. Lab. Anim. Resour. 2009, 51, 3–23. [Google Scholar] [CrossRef]
- Massouh, M.; Saghatelyan, A. De-routing neuronal precursors in the adult brain to sites of injury: Role of the vasculature. Neuropharmacology 2010, 58, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Buylla, A.; Lim, D.A. For the long run: Maintaining germinal niches in the adult brain. Neuron 2004, 41, 683–686. [Google Scholar] [CrossRef]
- Doetsch, F.; Caille, I.; Lim, D.A.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell 1999, 97, 703–716. [Google Scholar] [CrossRef]
- Doetsch, F.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Cellular composition and three-dimensional organization of the subventricular germinal zone in the adult mammalian brain. J. Neurosci. 1997, 17, 5046–5061. [Google Scholar] [PubMed]
- Tombran-Tink, J.; Barnstable, C.J. PEDF: A multifaceted neurotrophic factor. Nat. Rev. Neurosci. 2003, 4, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Namba, T.; Yabe, T.; Gonda, Y.; Ichikawa, N.; Sanagi, T.; Arikawa-Hirasawa, E.; Mochizuki, H.; Kohsaka, S.; Uchino, S. Pigment epithelium-derived factor up-regulation induced by memantine, an N-methyl-d-aspartate receptor antagonist, is involved in increased proliferation of hippocampal progenitor cells. Neuroscience 2010, 167, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Castro-Garcia, P.; Diaz-Moreno, M.; Gil-Gas, C.; Fernandez-Gomez, F.J.; Honrubia-Gomez, P.; Alvarez-Simon, C.B.; Sanchez-Sanchez, F.; Cano, J.C.; Almeida, F.; Blanco, V.; et al. Defects in subventricular zone pigmented epithelium-derived factor niche signaling in the senescence-accelerated mouse prone-8. FASEB J. 2015, 29, 1480–1492. [Google Scholar] [CrossRef] [PubMed]
- Pignolo, R.J.; Cristofalo, V.J.; Rotenberg, M.O. Senescent WI-38 cells fail to express EPC-1, a gene induced in young cells upon entry into the G0 state. J. Biol. Chem. 1993, 268, 8949–8957. [Google Scholar] [PubMed]
- Gaiano, N.; Fishell, G. The role of notch in promoting glial and neural stem cell fates. Ann. Rev. Neurosci. 2002, 25, 471–490. [Google Scholar] [CrossRef] [PubMed]
- Andreu-Agullo, C.; Morante-Redolat, J.M.; Delgado, A.C.; Farinas, I. Vascular niche factor PEDF modulates notch-dependent stemness in the adult subependymal zone. Nat. Neurosci. 2009, 12, 1514–1523. [Google Scholar] [CrossRef] [PubMed]
- Doyon, G.; St-Jean, S.; Darsigny, M.; Asselin, C.; Boudreau, F. Nuclear receptor co-repressor is required to maintain proliferation of normal intestinal epithelial cells in culture and down-modulates the expression of pigment epithelium-derived factor. J. Biol. Chem. 2009, 284, 25220–25229. [Google Scholar] [CrossRef] [PubMed]
- Brack, A.S.; Conboy, I.M.; Conboy, M.J.; Shen, J.; Rando, T.A. A temporal switch from notch to wnt signaling in muscle stem cells is necessary for normal adult myogenesis. Cell Stem Cell 2008, 2, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Hurlbut, G.D.; Kankel, M.W.; Lake, R.J.; Artavanis-Tsakonas, S. Crossing paths with notch in the hyper-network. Curr. Opin. Cell Biol. 2007, 19, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Schira, J.; Falkenberg, H.; Hendricks, M.; Waldera-Lupa, D.M.; Kogler, G.; Meyer, H.E.; Muller, H.W.; Stuhler, K. Characterization of regenerative phenotype of unrestricted somatic stem cells (USSC) from human umbilical cord blood (hUCB) by functional secretome analysis. Mol. Cell. Proteom. 2015, 14, 2630–2643. [Google Scholar] [CrossRef] [PubMed]
- Holland, E.C. Gliomagenesis: Genetic alterations and mouse models. Nat. Rev. Genet. 2001, 2, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Holland, E.C.; Celestino, J.; Dai, C.; Schaefer, L.; Sawaya, R.E.; Fuller, G.N. Combined activation of Ras and Akt in neural progenitors induces glioblastoma formation in mice. Nat. Genet. 2000, 25, 55–57. [Google Scholar] [CrossRef] [PubMed]
- Alcantara Llaguno, S.; Chen, J.; Kwon, C.-H.; Jackson, E.L.; Li, Y.; Burns, D.K.; Alvarez-Buylla, A.; Parada, L.F. Malignant astrocytomas originate from neural stem/progenitor cells in a somatic tumor suppressor mouse model. Cancer Cell 2009, 15, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.K.; Wang, Y.; Belz, T.; Bock, D.; Takacs, A.; Radlwimmer, B.; Barbus, S.; Reifenberger, G.; Lichter, P.; Schutz, G. The nuclear receptor tailless induces long-term neural stem cell expansion and brain tumor initiation. Genes Dev. 2010, 24, 683–695. [Google Scholar] [PubMed]
- Wang, Y.; Yang, J.; Zheng, H.; Tomasek, G.J.; Zhang, P.; McKeever, P.E.; Lee, E.Y.H.P.; Zhu, Y. Expression of mutant p53 proteins implicates a lineage relationship between neural stem cells and malignant astrocytic glioma in a murine model. Cancer Cell 2009, 15, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Park, G.; Kim, T.H.; Hong, J.H.; Kim, Y.J.; Jin, X.; Kang, S.; Jung, J.E.; Kim, J.Y.; Yun, H.; et al. Pigment epithelium-derived factor (PEDF) expression induced by EGFRvIII promotes self-renewal and tumor progression of glioma stem cells. PLoS Biol. 2015, 13, e1002152. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.; Yam, H.F.; Su, B.; Chan, K.P.; Pang, C.P.; Liu, W.W.; Zhang, W.Z.; Lu, Y. Loss of pigment epithelium derived factor expression in glioma progression. J. Clin. Pathol. 2003, 56, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, Z.; Ding, T.; Chen, Z.; Zhang, T. Mesenchymal stem cells overexpressing PEDF decrease the angiogenesis of gliomas. Biosci. Rep. 2013, 33, e00019. [Google Scholar] [CrossRef] [PubMed]
- Gehrs, K.M.; Anderson, D.H.; Johnson, L.V.; Hageman, G.S. Age-related macular degeneration—Emerging pathogenetic and therapeutic concepts. Ann. Med. 2006, 38, 450–471. [Google Scholar] [CrossRef] [PubMed]
- Arnhold, S.; Heiduschka, P.; Klein, H.; Absenger, Y.; Basnaoglu, S.; Kreppel, F.; Henke-Fahle, S.; Kochanek, S.; Bartz-Schmidt, K.U.; Addicks, K.; et al. Adenovirally transduced bone marrow stromal cells differentiate into pigment epithelial cells and induce rescue effects in RCS rats. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4121–4129. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Deng, X.; Spee, C.; Sonoda, S.; Hsieh, C.L.; Barron, E.; Pera, M.; Hinton, D.R. Polarized secretion of PEDF from human embryonic stem cell-derived RPE promotes retinal progenitor cell survival. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1573–1585. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Malcuit, C.; Wang, S.; Girman, S.; Francis, P.; Lemieux, L.; Lanza, R.; Lund, R. Long-term safety and function of RPE from human embryonic stem cells in preclinical models of macular degeneration. Stem Cells 2009, 27, 2126–2135. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Mandai, M.; Kamao, H.; Hashiguchi, T.; Shikamura, M.; Kawamata, S.; Sugita, S.; Takahashi, M. Protective effects of human IPS-derived retinal pigmented epithelial cells in comparison with human mesenchymal stromal cells and human neural stem cells on the degenerating retina in rd1 mice. Stem Cells 2015, 33, 1543–1553. [Google Scholar] [CrossRef] [PubMed]
- Cotsarelis, G.; Cheng, S.Z.; Dong, G.; Sun, T.T.; Lavker, R.M. Existence of slow-cycling limbal epithelial basal cells that can be preferentially stimulated to proliferate: Implications on epithelial stem cells. Cell 1989, 57, 201–209. [Google Scholar] [CrossRef]
- Ho, T.C.; Chen, S.L.; Wu, J.Y.; Ho, M.Y.; Chen, L.J.; Hsieh, J.W.; Cheng, H.C.; Tsao, Y.P. PEDF promotes self-renewal of limbal stem cell and accelerates corneal epithelial wound healing. Stem Cells 2013, 31, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sagheer, U.; Gong, J.; Chung, C. Pigment Epithelium-Derived Factor (PEDF) is a Determinant of Stem Cell Fate: Lessons from an Ultra-Rare Disease. J. Dev. Biol. 2015, 3, 112-128. https://doi.org/10.3390/jdb3040112
Sagheer U, Gong J, Chung C. Pigment Epithelium-Derived Factor (PEDF) is a Determinant of Stem Cell Fate: Lessons from an Ultra-Rare Disease. Journal of Developmental Biology. 2015; 3(4):112-128. https://doi.org/10.3390/jdb3040112
Chicago/Turabian StyleSagheer, Usman, Jingjing Gong, and Chuhan Chung. 2015. "Pigment Epithelium-Derived Factor (PEDF) is a Determinant of Stem Cell Fate: Lessons from an Ultra-Rare Disease" Journal of Developmental Biology 3, no. 4: 112-128. https://doi.org/10.3390/jdb3040112
APA StyleSagheer, U., Gong, J., & Chung, C. (2015). Pigment Epithelium-Derived Factor (PEDF) is a Determinant of Stem Cell Fate: Lessons from an Ultra-Rare Disease. Journal of Developmental Biology, 3(4), 112-128. https://doi.org/10.3390/jdb3040112