Integrins Have Cell-Type-Specific Roles in the Development of Motor Neuron Connectivity

 and
and 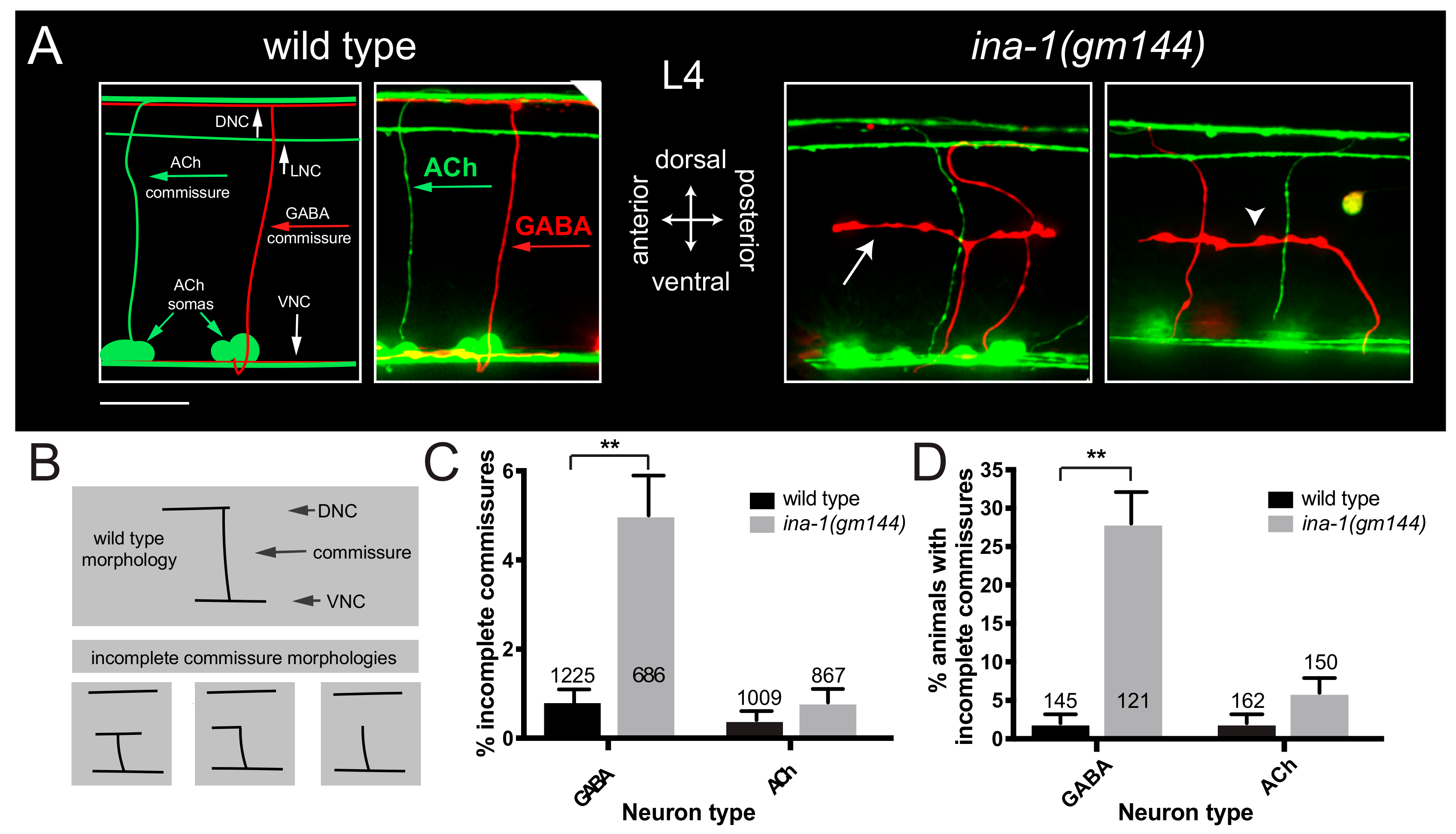{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. C. elegans Strains
2.2. Construction of Plasmids
2.3. ina-1 Reporter and Rescue Constructs
2.4. Microscopy
2.5. Commissure Analysis
2.6. Worm Staging
2.7. Fluorescence Intensity Analysis
2.8. Statistical Analysis
3. Results
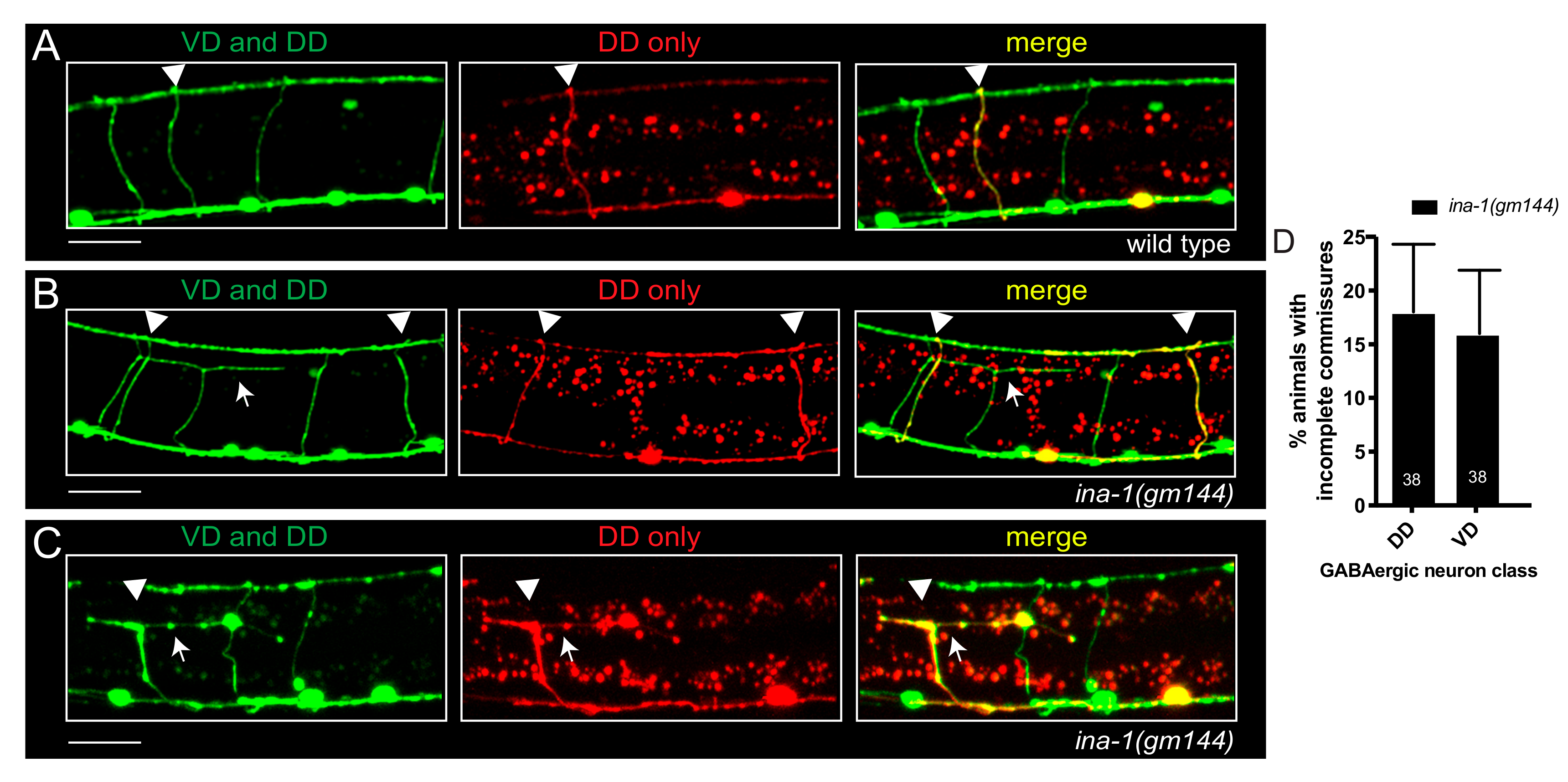
3.1. α Integrin/ina-1 Is Necessary for Commissural Patterning in a Neuron-Specific Manner
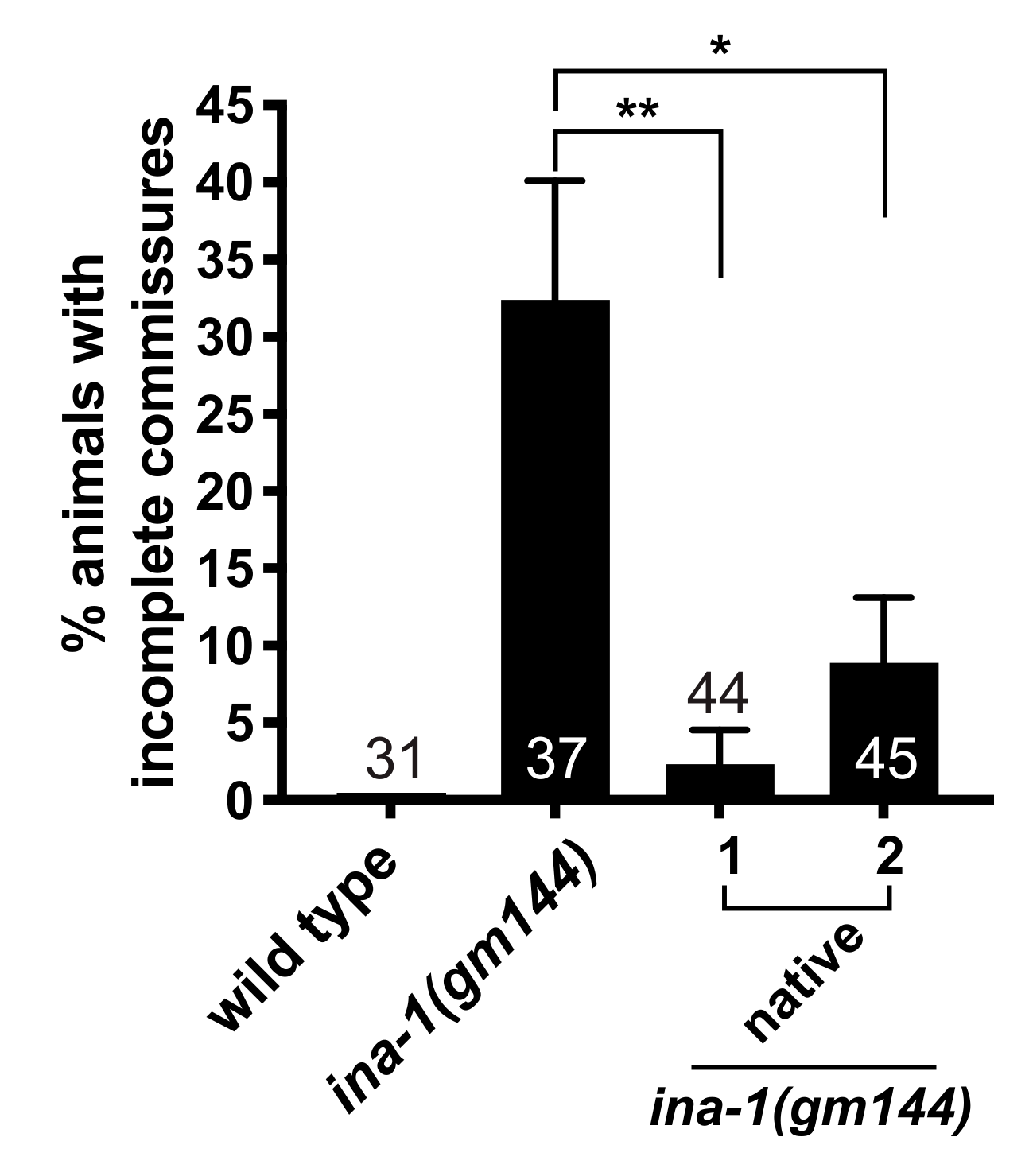
3.2. GABAergic Commissural Defects in ina-1 Mutants Are Rescued by Expression of Wild Type ina-1
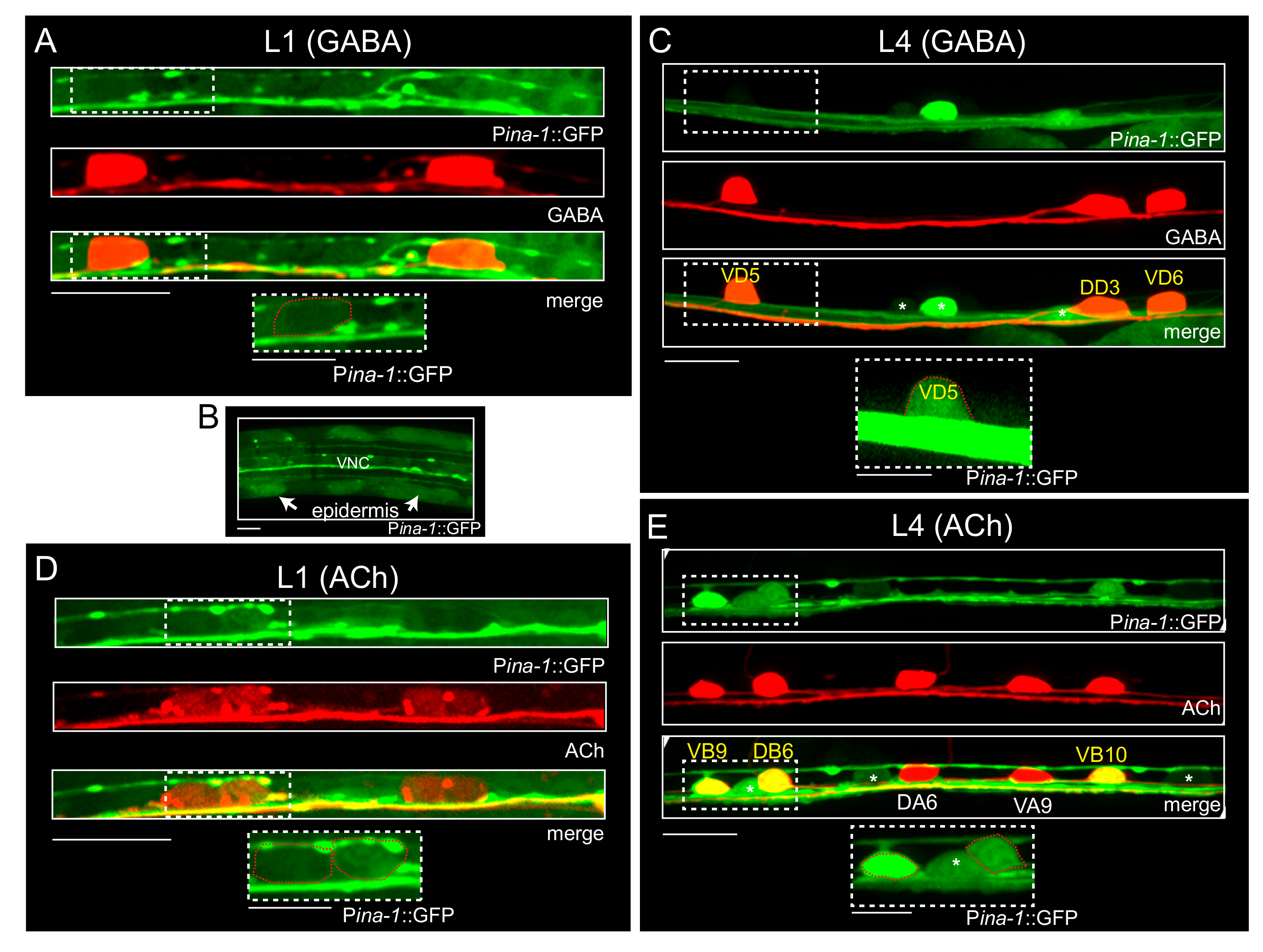
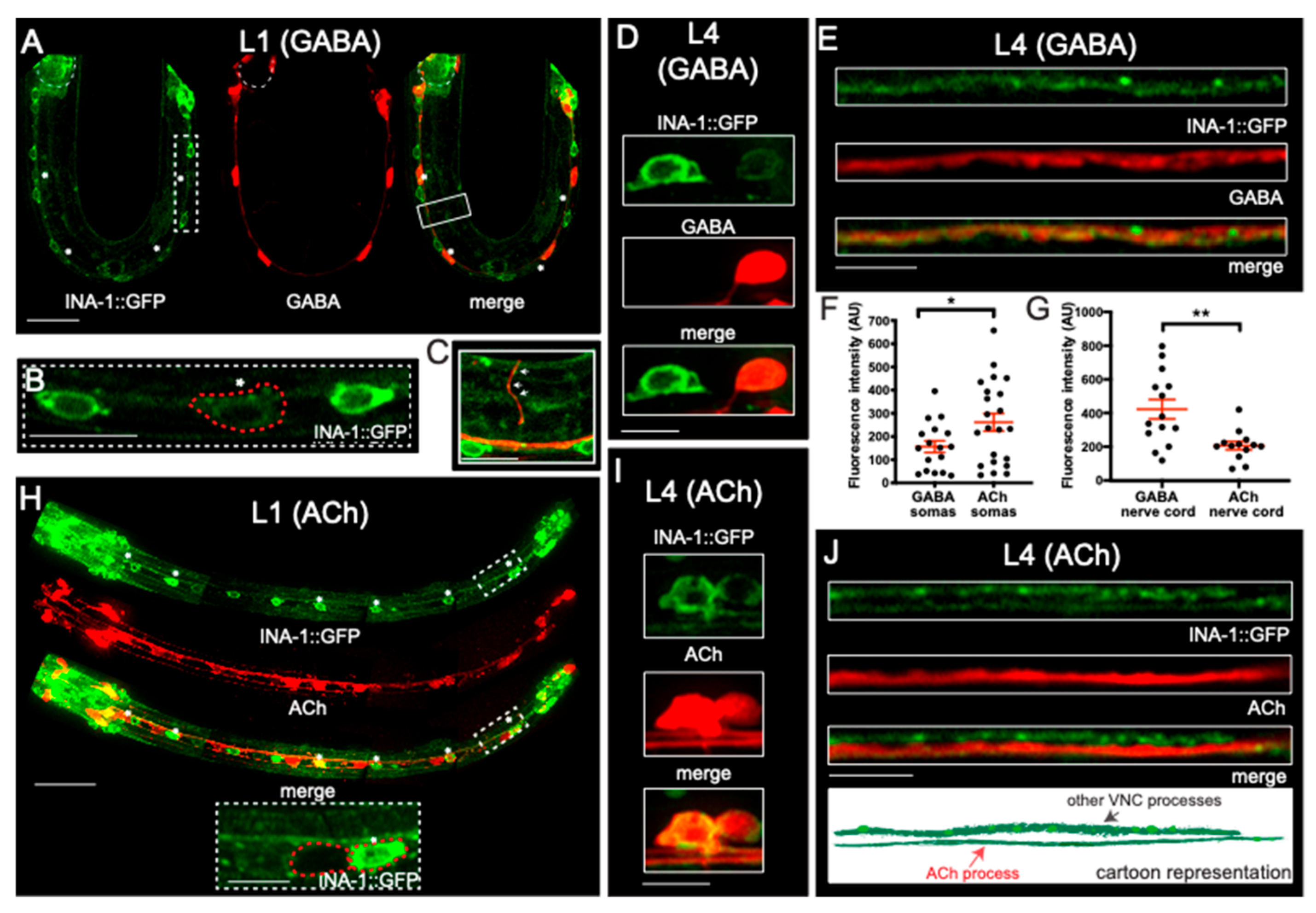
3.3. α Integrin/INA-1 Is Expressed in Both GABAergic and Cholinergic Motor Neurons
3.4. α Integrin/ina-1 Is Necessary for Embryonic and Post-Embryonic GABAergic Commissural Patterning
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- DeSimone, D.W.; Stepp, M.A.; Patel, R.S.; Hynes, R.O. The integrin family of cell surface receptors. Biochem. Soc. Trans. 1987, 15, 789–791. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. Integrins: versatility, modulation, and signaling in cell adhesion. Cell 1992, 69, 11–25. [Google Scholar] [CrossRef]
- Hynes, R.O. Integrins: bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef]
- Humphries, J.D.; Byron, A.; Humphries, M.J. Integrin ligands at a glance. J. Cell Sci. 2006, 119, 3901–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, N.H. Cell-cell adhesion via the ECM: integrin genetics in fly and worm. Matrix Biol. 2000, 19, 191–201. [Google Scholar] [CrossRef]
- De Arcangelis, A.; Georges-Labouesse, E. Integrin and ECM functions: roles in vertebrate development. Trends Genet. 2000, 16, 389–395. [Google Scholar] [CrossRef]
- Van der Flier, A.; Sonnenberg, A. Function and interactions of integrins. Cell Tissue Res. 2001, 305, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.; D’Amico, G.; Hodivala-Dilke, K.M.; Reynolds, L.E. Integrins: The keys to unlocking angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1703–1713. [Google Scholar] [CrossRef] [PubMed]
- Sixt, M.; Bauer, M.; Lammermann, T.; Fassler, R. Beta1 integrins: Zip codes and signaling relay for blood cells. Curr. Opin. Cell Biol. 2006, 18, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Rose, D.M.; Ginsberg, M.H. Integrins in the immune system. Adv. Immunol. 1999, 72, 325–380. [Google Scholar] [PubMed]
- Sonnenberg, A. Integrins and their ligands. Curr. Top. Microbiol. Immunol. 1993, 184, 7–35. [Google Scholar] [PubMed]
- Cirulli, V.; Yebra, M. Netrins: Beyond the brain. Nat. Rev. Mol. Cell Biol. 2007, 8, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Lemons, M.L.; Abanto, M.L.; Dambrouskas, N.; Clements, C.C.; Deloughery, Z.; Garozzo, J.; Condic, M.L. Integrins and cAMP mediate netrin-induced growth cone collapse. Brain Res. 2013, 1537, 46–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagedorn, E.J.; Yashiro, H.; Ziel, J.W.; Ihara, S.; Wang, Z.; Sherwood, D.R. Integrin acts upstream of netrin signaling to regulate formation of the anchor cell’s invasive membrane in C. elegans. Dev. Cell 2009, 17, 187–198. [Google Scholar] [CrossRef]
- Serini, G.; Valdembri, D.; Zanivan, S.; Morterra, G.; Burkhardt, C.; Caccavari, F.; Zammataro, L.; Primo, L.; Tamagnone, L.; Logan, M.; et al. Class 3 semaphorins control vascular morphogenesis by inhibiting integrin function. Nature 2003, 424, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Pasterkamp, R.J.; Peschon, J.J.; Spriggs, M.K.; Kolodkin, A.L. Semaphorin 7A promotes axon outgrowth through integrins and MAPKs. Nature 2003, 424, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Okuno, T.; Yamamoto, M.; Pasterkamp, R.J.; Takegahara, N.; Takamatsu, H.; Kitao, T.; Takagi, J.; Rennert, P.D.; Kolodkin, A.L.; et al. Semaphorin 7A initiates T-cell-mediated inflammatory responses through alpha1beta1 integrin. Nature 2007, 446, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Stevens, A.; Jacobs, J.R. Integrins regulate responsiveness to slit repellent signals. J. Neurosci. 2002, 22, 4448–4455. [Google Scholar] [CrossRef]
- Davy, A.; Robbins, S.M. Ephrin-A5 modulates cell adhesion and morphology in an integrin-dependent manner. EMBO J. 2000, 19, 5396–5405. [Google Scholar] [CrossRef] [Green Version]
- Huai, J.; Drescher, U. An ephrin-A-dependent signaling pathway controls integrin function and is linked to the tyrosine phosphorylation of a 120-kDa protein. J. Biol. Chem. 2001, 276, 6689–6694. [Google Scholar] [CrossRef]
- Huynh-Do, U.; Vindis, C.; Liu, H.; Cerretti, D.P.; McGrew, J.T.; Enriquez, M.; Chen, J.; Daniel, T.O. Ephrin-B1 transduces signals to activate integrin-mediated migration, attachment and angiogenesis. J. Cell Sci. 2002, 115, 3073–3081. [Google Scholar] [PubMed]
- Nakamoto, T.; Kain, K.H.; Ginsberg, M.H. Neurobiology: New connections between integrins and axon guidance. Curr. Biol. 2004, 14, R121–R123. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, M.H. Integrins: Dynamic regulation of ligand binding. Biochem. Soc. Trans. 1995, 23, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Humphries, M.J. The molecular basis and specificity of integrin-ligand interactions. J. Cell Sci. 1990, 97, 585–592. [Google Scholar] [PubMed]
- Plow, E.F.; Meller, J.; Byzova, T.V. Integrin function in vascular biology: A view from 2013. Curr. Opin. Hematol. 2014, 21, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Gettner, S.N.; Kenyon, C.; Reichardt, L.F. Characterization of beta pat-3 heterodimers, a family of essential integrin receptors in C. elegans. J. Cell Biol. 1995, 129, 1127–1141. [Google Scholar] [CrossRef]
- Williams, B.D.; Waterston, R.H. Genes critical for muscle development and function in Caenorhabditis elegans identified through lethal mutations. J. Cell Biol. 1994, 124, 475–490. [Google Scholar] [CrossRef] [Green Version]
- Baum, P.D.; Garriga, G. Neuronal migrations and axon fasciculation are disrupted in ina-1 integrin mutants. Neuron 1997, 19, 51–62. [Google Scholar] [CrossRef]
- Forrester, W.C.; Garriga, G. Genes necessary for C. elegans cell and growth cone migrations. Development 1997, 124, 1831–1843. [Google Scholar]
- Poinat, P.; De Arcangelis, A.; Sookhareea, S.; Zhu, X.; Hedgecock, E.M.; Labouesse, M.; Georges-Labouesse, E. A conserved interaction between beta1 integrin/PAT-3 and Nck-interacting kinase/MIG-15 that mediates commissural axon navigation in C. elegans. Curr. Biol. 2002, 12, 622–631. [Google Scholar] [CrossRef]
- Chisholm, A.D.; Hutter, H.; Jin, Y.; Wadsworth, W.G. The Genetics of Axon Guidance and Axon Regeneration in Caenorhabditis elegans. Genetics 2016, 204, 849–882. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [PubMed]
- Mello, C.C.; Kramer, J.M.; Stinchcomb, D.; Ambros, V. Efficient gene transfer in C. elegans: Extrachromosomal maintenance and integration of transforming sequences. EMBO J. 1991, 10, 3959–3970. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, B.; Philbrook, A.; Touroutine, D.; Banerjee, N.; Oliver, D.; Lambert, C.M.; Francis, M.M. Excitatory neurons sculpt GABAergic neuronal connectivity in the C. elegans motor circuit. Development 2017, 144, 1807–1819. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, R.; Touroutine, D.; Barbagallo, B.; Climer, J.; Lambert, C.M.; Clark, C.M.; Alkema, M.J.; Francis, M.M. A conserved dopamine-cholecystokinin signaling pathway shapes context-dependent Caenorhabditis elegans behavior. PLoS Genet. 2014, 10, e1004584. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, B.; Prescott, H.A.; Boyle, P.; Climer, J.; Francis, M.M. A dominant mutation in a neuronal acetylcholine receptor subunit leads to motor neuron degeneration in Caenorhabditis elegans. J. Neurosci. 2010, 30, 13932–13942. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.H. Handbook of Biological Statistics, 3rd ed.; Sparky House Publishing: Baltimore, MD, USA, 2014; p. 299. [Google Scholar]
- Meighan, C.M.; Schwarzbauer, J.E. alpha Integrin cytoplasmic tails have tissue-specific roles during C. elegans development. Int. J. Dev. Biol. 2014, 58, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Wehrli, M.; DiAntonio, A.; Fearnley, I.M.; Smith, R.J.; Wilcox, M. Cloning and characterization of alpha PS1, a novel Drosophila melanogaster integrin. Mech. Dev. 1993, 43, 21–36. [Google Scholar] [CrossRef]
- Morrissey, M.A.; Keeley, D.P.; Hagedorn, E.J.; McClatchey, S.T.H.; Chi, Q.; Hall, D.H.; Sherwood, D.R. B-LINK: A hemicentin, plakin, and integrin-dependent adhesion system that links tissues by connecting adjacent basement membranes. Deve. Cell 2014, 31, 319–331. [Google Scholar] [CrossRef]
- Jayadev, R.; Chi, Q.; Keeley, D.P.; Hastie, E.L.; Kelley, L.C.; Sherwood, D.R. α-Integrins dictate distinct modes of type IV collagen recruitment to basement membranes. J. Cell Biol. 2019. [Google Scholar] [CrossRef]
- Bulow, H.E.; Hobert, O. Differential sulfations and epimerization define heparan sulfate specificity in nervous system development. Neuron 2004, 41, 723–736. [Google Scholar] [CrossRef]
- Leptin, M.; Bogaert, T.; Lehmann, R.; Wilcox, M. The function of PS integrins during Drosophila embryogenesis. Cell 1989, 56, 401–408. [Google Scholar] [CrossRef]
- O’Toole, T.E.; Loftus, J.C.; Plow, E.F.; Glass, A.A.; Harper, J.R.; Ginsberg, M.H. Efficient surface expression of platelet GPIIb-IIIa requires both subunits. Blood 1989, 74, 14–18. [Google Scholar] [PubMed]
- Meighan, C.M.; Schwarzbauer, J.E. Control of C. elegans hermaphrodite gonad size and shape by vab-3/Pax6-mediated regulation of integrin receptors. Genes Dev. 2007, 21, 1615–1620. [Google Scholar] [CrossRef]
- Hunt-Newbury, R.; Viveiros, R.; Johnsen, R.; Mah, A.; Anastas, D.; Fang, L.; Halfnight, E.; Lee, D.; Lin, J.; Lorch, A.; et al. High-throughput in vivo analysis of gene expression in Caenorhabditis elegans. PLoS Biol. 2007, 5, e237. [Google Scholar] [CrossRef]
- Locke, C.J.; Kautu, B.B.; Berry, K.P.; Lee, S.K.; Caldwell, K.A.; Caldwell, G.A. Pharmacogenetic analysis reveals a post-developmental role for Rac GTPases in Caenorhabditis elegans GABAergic neurotransmission. Genetics 2009, 183, 1357–1372. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliver, D.; Norman, E.; Bates, H.; Avard, R.; Rettler, M.; Bénard, C.Y.; Francis, M.M.; Lemons, M.L. Integrins Have Cell-Type-Specific Roles in the Development of Motor Neuron Connectivity. J. Dev. Biol. 2019, 7, 17. https://doi.org/10.3390/jdb7030017
Oliver D, Norman E, Bates H, Avard R, Rettler M, Bénard CY, Francis MM, Lemons ML. Integrins Have Cell-Type-Specific Roles in the Development of Motor Neuron Connectivity. Journal of Developmental Biology. 2019; 7(3):17. https://doi.org/10.3390/jdb7030017
Chicago/Turabian StyleOliver, Devyn, Emily Norman, Heather Bates, Rachel Avard, Monika Rettler, Claire Y. Bénard, Michael M. Francis, and Michele L. Lemons. 2019. "Integrins Have Cell-Type-Specific Roles in the Development of Motor Neuron Connectivity" Journal of Developmental Biology 7, no. 3: 17. https://doi.org/10.3390/jdb7030017
APA StyleOliver, D., Norman, E., Bates, H., Avard, R., Rettler, M., Bénard, C. Y., Francis, M. M., & Lemons, M. L. (2019). Integrins Have Cell-Type-Specific Roles in the Development of Motor Neuron Connectivity. Journal of Developmental Biology, 7(3), 17. https://doi.org/10.3390/jdb7030017