Mud Loss Restricts Yki-Dependent Hyperplasia in Drosophila Epithelia

Abstract
:1. Introduction
2. Materials and Methods
2.1. Drosophila Stocks and Maintenance
2.2. Antibody Staining
2.3. Edu Staining
2.4. Image Acquisition and Processing
2.5. RNA Sequencing
3. Results and Discussion
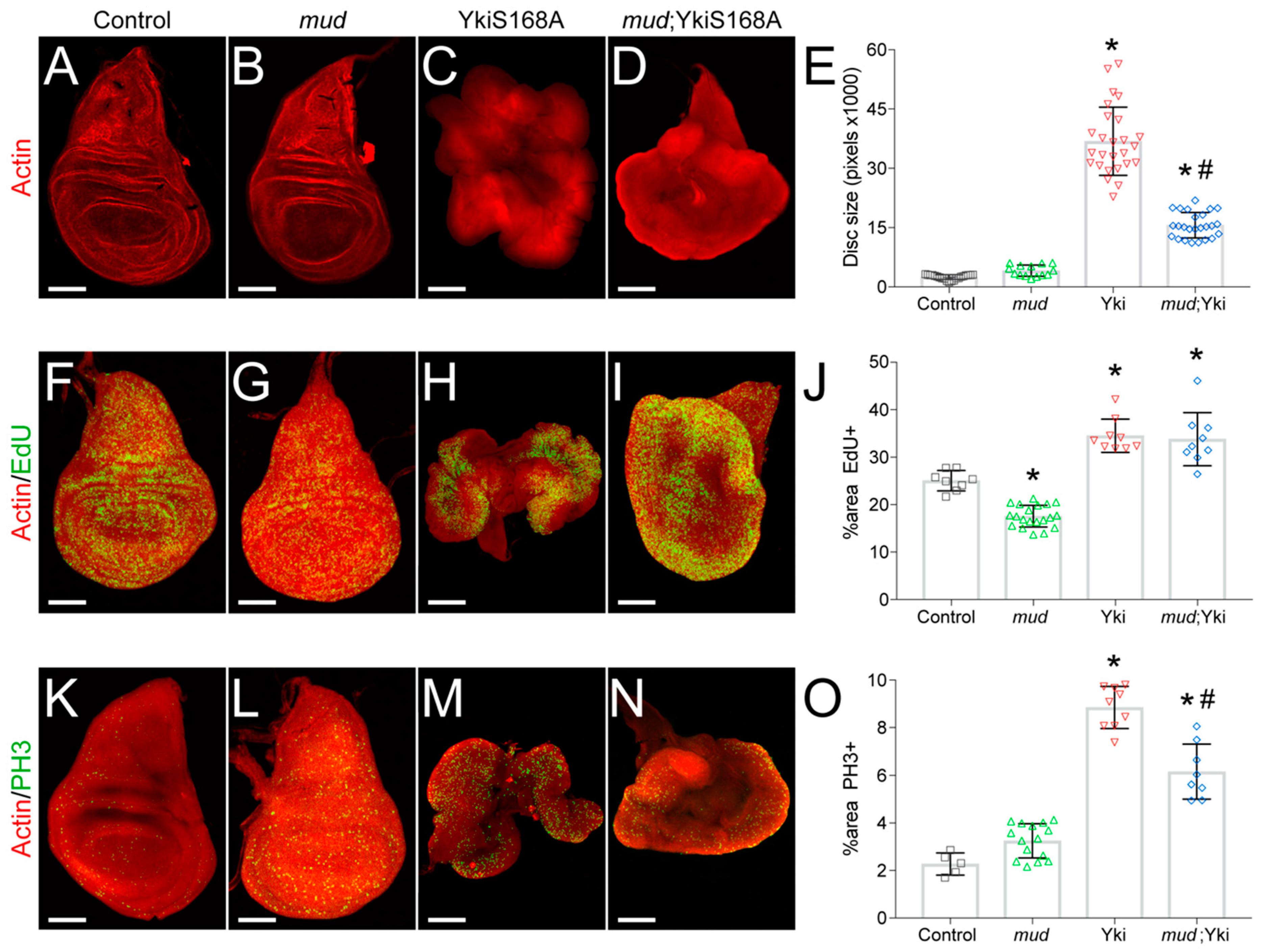
3.1. Mud Knockdown Attenuates YkiS168A-Driven Wing Disc Growth
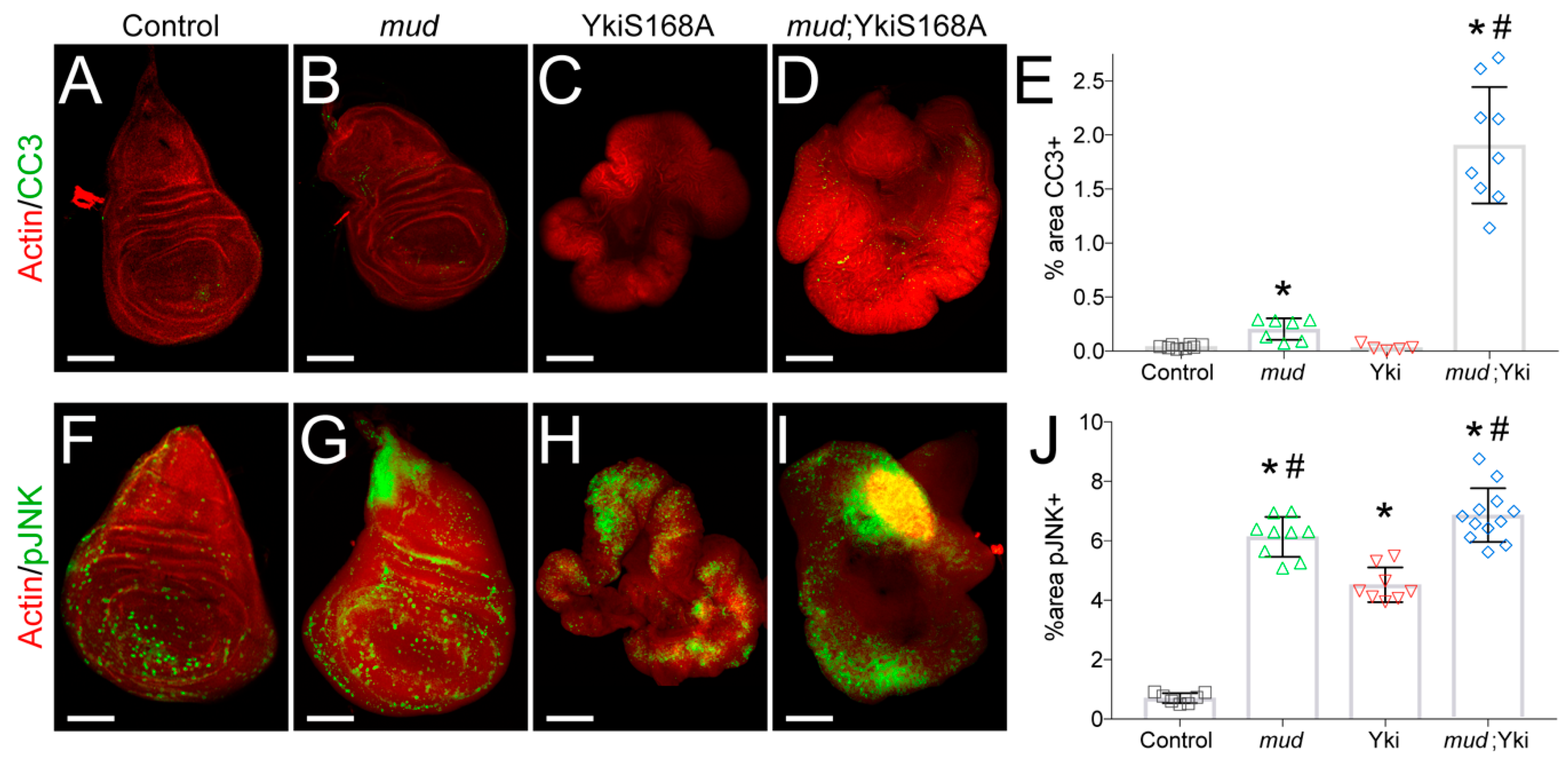
3.2. Mud Knockdown Induces JNK Activation and Apoptosis in YkiS168A-Expressing Wing Discs
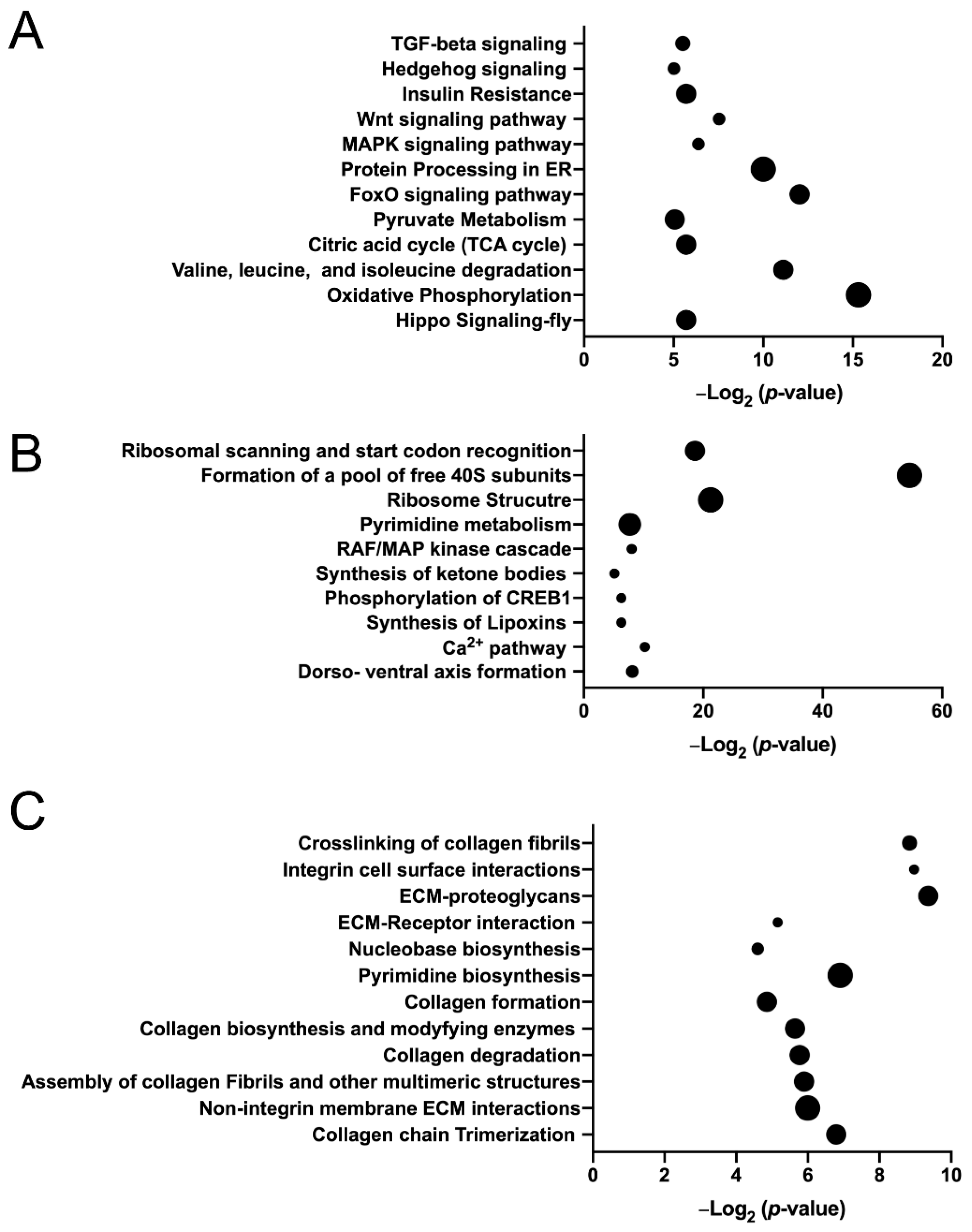
3.3. Mud Knockdown Alters the Transcriptional Landscape of Wing Disc Epithelial Cells
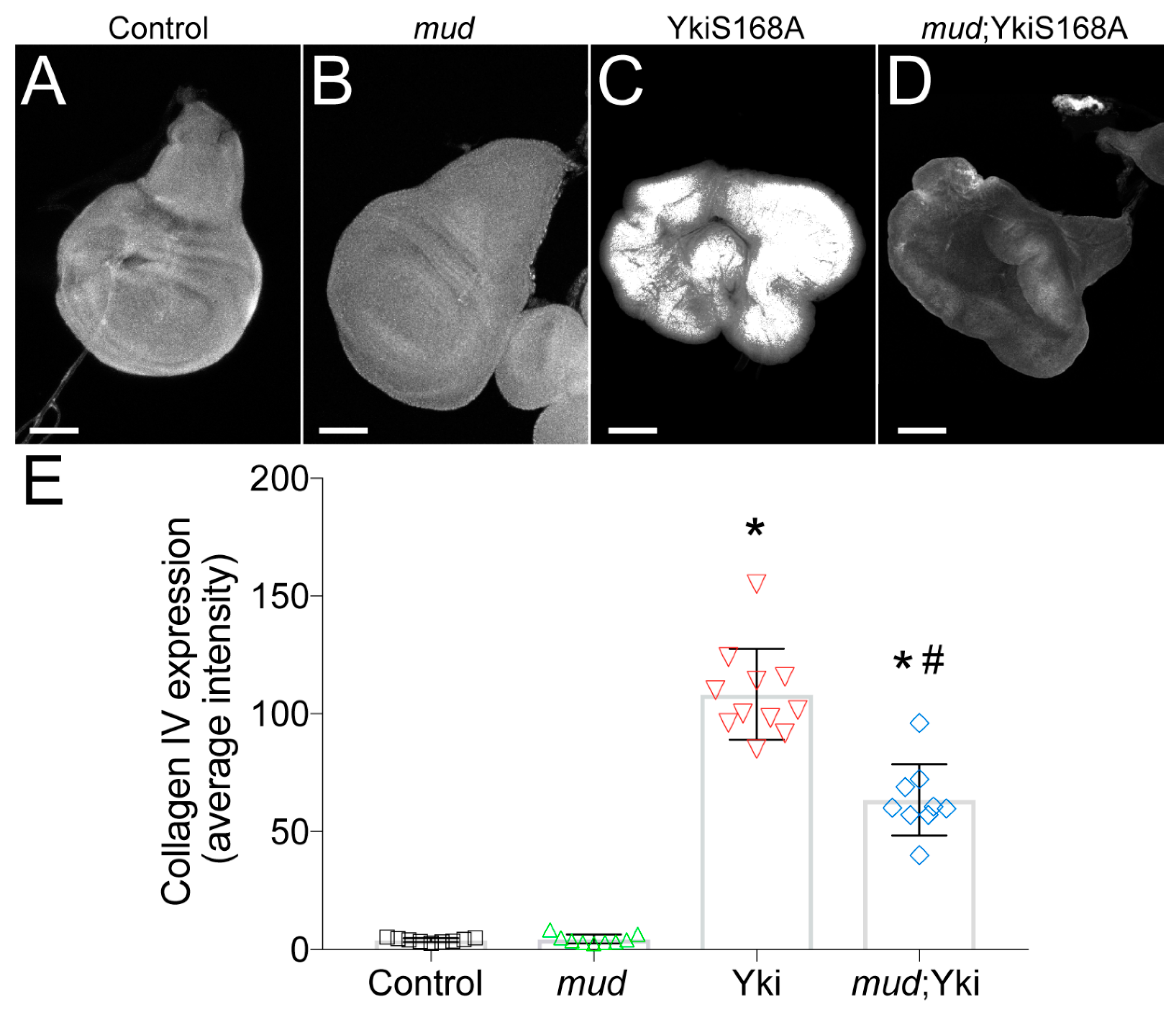
3.4. Restricted YkiS168A-Mediated Growth Is Associated with Reduced Collagen Expression
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ragkousi, K.; Gibson, M.C. Cell division and the maintenance of epithelial order. J. Cell Biol. 2014, 207, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial–mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Herranz, H.; Eichenlaub, T.; Cohen, S.M. Cancer in Drosophila. Curr. Top. Dev. Biol. 2016, 116, 181–199. [Google Scholar] [CrossRef] [PubMed]
- Gibson, M.C.; Schubiger, G. Peripodial Cells Regulate Proliferation and Patterning of Drosophila Imaginal Discs. Cell 2000, 103, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Aldaz, S.; Escudero, L.M. Imaginal discs. Curr. Biol. 2010, 20, R429–R431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Wu, S.; Barrera, J.; Matthews, K.; Pan, D. The Hippo Signaling Pathway Coordinately Regulates Cell Proliferation and Apoptosis by Inactivating Yorkie, the Drosophila Homolog of YAP. Cell 2005, 122, 421–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Huang, J.; Dong, J.; Pan, D. hippo Encodes a Ste-20 Family Protein Kinase that Restricts Cell Proliferation and Promotes Apoptosis in Conjunction with salvador and warts. Cell 2003, 114, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Harvey, K.F.; Zhang, X.; Thomas, D.M. The Hippo pathway and human cancer. Nat. Rev. Cancer 2013, 13, 246–257. [Google Scholar] [CrossRef]
- Snigdha, K.; Gangwani, K.S.; Lapalikar, G.V.; Singh, A.; Kango-Singh, M. Hippo Signaling in Cancer: Lessons From Drosophila Models. Front. Cell Dev. Biol. 2019, 7, 85. [Google Scholar] [CrossRef] [Green Version]
- Staley, B.K.; Irvine, K.D. Hippo signaling inDrosophila: Recent advances and insights. Dev. Dyn. 2012, 241, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, S.U.; Eichenlaub, T.; Herranz, H. Yorkie and JNK Control Tumorigenesis in Drosophila Cells with Cytokinesis Failure. Cell Rep. 2018, 23, 1491–1503. [Google Scholar] [CrossRef] [PubMed]
- Groth, C.; Vaid, P.; Khatpe, A.; Prashali, N.; Ahiya, A.; Andrejeva, D.; Chakladar, M.; Nagarkar, S.; Paul, R.; Kelkar, D.; et al. Genome-Wide Screen for Context-Dependent Tumor Suppressors Identified Using in Vivo Models for Neoplasia in Drosophila. G3 Genes Genomes Genet. 2020, 10, 2999–3008. [Google Scholar] [CrossRef] [PubMed]
- Grzeschik, N.A.; Parsons, L.M.; Allott, M.L.; Harvey, K.F.; Richardson, H.E. Lgl, aPKC, and Crumbs Regulate the Salvador/Warts/Hippo Pathway through Two Distinct Mechanisms. Curr. Biol. 2010, 20, 573–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sander, M.; Eichenlaub, T.; Herranz, H. Oncogenic cooperation between Yorkie and the conserved microRNAmiR-8in the wing disc ofDrosophila. Development 2018, 145, dev153817. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Herranz, H.; Cohen, S.M. The chromatin remodeling BAP complex limits tumor-promoting activity of the Hippo pathway effector Yki to prevent neoplastic transformation in Drosophila epithelia. Dis. Model. Mech. 2017, 10, 1201–1209. [Google Scholar] [CrossRef] [Green Version]
- Sun, G.; Irvine, K.D. Regulation of Hippo signaling by Jun kinase signaling during compensatory cell proliferation and regeneration, and in neoplastic tumors. Dev. Biol. 2011, 350, 139–151. [Google Scholar] [CrossRef] [Green Version]
- Dewey, E.B.; Sanchez, D.; Johnston, C.A. Warts Phosphorylates Mud to Promote Pins-Mediated Mitotic Spindle Orientation in Drosophila, Independent of Yorkie. Curr. Biol. 2015, 25, 2751–2762. [Google Scholar] [CrossRef] [Green Version]
- Oh, H.; Irvine, K.D. In vivo regulation of Yorkie phosphorylation and localization. Development 2008, 135, 1081–1088. [Google Scholar] [CrossRef] [Green Version]
- Dewey, E.B.; Taylor, D.T.; Johnston, C.A. Rolling in the mud: Hippo controls oriented cell division. Cell Cycle 2016, 15, 607–608. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Leng, N.; Dawson, J.A.; Thomson, J.A.; Ruotti, V.; Rissman, A.I.; Smits, B.M.G.; Haag, J.D.; Gould, M.N.; Stewart, R.M.; Kendziorski, C. EBSeq: An empirical Bayes hierarchical model for inference in RNA-seq experiments. Bioinformatics 2013, 29, 1035–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, C.E.; Bailey, T.L.; Noble, W.S. FIMO: Scanning for occurrences of a given motif. Bioinformatics 2011, 27, 1017–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, B.A. From Cell Structure to Transcription: Hippo Forges a New Path. Cell 2006, 124, 267–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, D. Hippo signaling in organ size control. Genes Dev. 2007, 21, 886–897. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Feldmann, G.; Huang, J.; Wu, S.; Zhang, N.; Comerford, S.A.; Gayyed, M.F.; Anders, R.A.; Maitra, A.; Pan, D. Elucidation of a Universal Size-Control Mechanism in Drosophila and Mammals. Cell 2007, 130, 1120–1133. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.W.; Slattery, M.; Mann, R.S. Transcription factor choice in the Hippo signaling pathway: Homothorax and yorkie regulation of the microRNA bantam in the progenitor domain of the Drosophila eye imaginal disc. Genes Dev. 2009, 23, 2307–2319. [Google Scholar] [CrossRef] [Green Version]
- Ren, F.; Wang, B.; Yue, T.; Yun, E.-Y.; Ip, Y.T.; Jiang, J. Hippo signaling regulates Drosophila intestine stem cell proliferation through multiple pathways. Proc. Natl. Acad. Sci. USA 2010, 107, 21064–21069. [Google Scholar] [CrossRef] [Green Version]
- Mikhailov, A.; Rieder, C.L. Cell cycle: Stressed out of mitosis. Curr. Biol. 2002, 12, R331–R333. [Google Scholar] [CrossRef] [Green Version]
- Stark, G.R.; Taylor, W.R. Control of the G2/M Transition. Mol. Biotechnol. 2006, 32, 227–248. [Google Scholar] [CrossRef]
- Dewey, E.B.; Johnston, C.A. Diverse mitotic functions of the cytoskeletal cross-linking protein Shortstop suggest a role in Dynein/Dynactin activity. Mol. Biol. Cell 2017, 28, 2555–2568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, Y.-I.; Meyer, E.J.; Kroesen, A.; McKinney, S.A.; Gibson, M.C. Epithelial junctions maintain tissue architecture by directing planar spindle orientation. Nat. Cell Biol. 2013, 500, 359–362. [Google Scholar] [CrossRef]
- Poulton, J.S.; Cuningham, J.C.; Peifer, M. Acentrosomal Drosophila epithelial cells exhibit abnormal cell division, leading to cell death and compensatory proliferation. Dev. Cell 2014, 30, 731–745. [Google Scholar] [CrossRef] [Green Version]
- Poulton, J.; McKay, D.J.; Peifer, M. Centrosome Loss Triggers a Transcriptional Program To Counter Apoptosis-Induced Oxidative Stress. Genetics 2019, 212, 187–211. [Google Scholar] [CrossRef]
- Sabino, D.; Gogendeau, D.; Gambarotto, D.; Nano, M.; Pennetier, C.; Dingli, F.; Arras, G.; Loew, D.; Basto, R. Moesin Is a Major Regulator of Centrosome Behavior in Epithelial Cells with Extra Centrosomes. Curr. Biol. 2015, 25, 879–889. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Sun, J.; Wang, D.; Pflugfelder, G.O.; Shen, J. Fold formation at the compartment boundary of Drosophila wing requires Yki signaling to suppress JNK dependent apoptosis. Sci. Rep. 2016, 6, 38003. [Google Scholar] [CrossRef] [Green Version]
- Pinal, N.; Calleja, M.; Morata, G. Pro-apoptotic and pro-proliferation functions of the JNK pathway of Drosophila: Roles in cell competition, tumorigenesis and regeneration. Open Biol. 2019, 9, 180256. [Google Scholar] [CrossRef] [Green Version]
- La Marca, J.E.; Richardson, H.E. Two-Faced: Roles of JNK Signalling During Tumourigenesis in the Drosophila Model. Front. Cell Dev. Biol. 2020, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, G.J.; Tsuji, T.; Cross, J.V.; Davis, R.J.; Templeton, D.J.; Jiang, W.; Ronai, Z.A. JNK-mediated Phosphorylation of Cdc25C Regulates Cell Cycle Entry and G2/M DNA Damage Checkpoint. J. Biol. Chem. 2010, 285, 14217–14228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, S.; Yoshioka, K.; Kizu, R.; Nakagama, H.; Matsunaga, T.; Ishizaka, Y.; Poon, R.Y.; Yamashita, K. Stress-Activated Mitogen-Activated Protein Kinases c-Jun NH2-Terminal Kinase and p38 Target Cdc25B for Degradation. Cancer Res. 2009, 69, 6438–6444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosolo, A.; Jaiswal, J.; Csordás, G.; Grass, I.; Uhlirova, M.; Classen, A.-K. JNK-dependent cell cycle stalling in G2 promotes survival and senescence-like phenotypes in tissue stress. eLife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Neto-Silva, R.M.; De Beco, S.; Johnston, L.A. Evidence for a Growth-Stabilizing Regulatory Feedback Mechanism between Myc and Yorkie, the Drosophila Homolog of Yap. Dev. Cell 2010, 19, 507–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziosi, M.; Baena-López, L.A.; Grifoni, D.; Froldi, F.; Pession, A.; Garoia, F.; Trotta, V.; Bellosta, P.; Cavicchi, S.; Pession, A. dMyc Functions Downstream of Yorkie to Promote the Supercompetitive Behavior of Hippo Pathway Mutant Cells. PLoS Genet. 2010, 6, e1001140. [Google Scholar] [CrossRef] [Green Version]
- Hamaratoglu, F.; Atkins, M. Rounding up the Usual Suspects: Assessing Yorkie, AP-1, and Stat Coactivation in Tumorigenesis. Int. J. Mol. Sci. 2020, 21, 4580. [Google Scholar] [CrossRef]
- Djiane, A.; Krejci, A.; Bernard, F.; Fexova, S.; Millen, K.; Bray, S.J. Dissecting the mechanisms of Notch induced hyperplasia. EMBO J. 2012, 32, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Doggett, K.; Turkel, N.; Willoughby, L.F.; Ellul, J.; Murray, M.J.; Richardson, H.; Brumby, A.M. BTB-Zinc Finger Oncogenes Are Required for Ras and Notch-Driven Tumorigenesis in Drosophila. PLoS ONE 2015, 10, e0132987. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Andrejeva, D.; Freitas, F.C.P.; Cohen, S.M.; Herranz, H. dTcf/Pangolinsuppresses growth and tumor formation inDrosophila. Proc. Natl. Acad. Sci. USA 2019, 116, 14055–14064. [Google Scholar] [CrossRef] [Green Version]
- Broadie, K.; Baumgartner, S.; Prokop, A. Extracellular matrix and its receptors in drosophila neural development. Dev. Neurobiol. 2011, 71, 1102–1130. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.N.; Horne-Badovinac, S.; Naba, A. In-silico definition of the Drosophila melanogaster matrisome. Matrix Biol. Plus 2019, 4. [Google Scholar] [CrossRef]
- Ikmi, A.; Gaertner, B.; Seidel, C.; Srivastava, M.; Zeitlinger, J.; Gibson, M.C. Molecular Evolution of the Yap/Yorkie Proto-Oncogene and Elucidation of Its Core Transcriptional Program. Mol. Biol. Evol. 2014, 31, 1375–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittkorn, E.; Sarkar, A.; Garcia, K.; Kango-Singh, M.; Singh, A. The Hippo pathway effector Yki downregulates Wg signaling to promote retinal differentiation in the Drosophila eye. Development 2015, 142, 2002–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunker, B.D.; Nellimoottil, T.T.; Boileau, R.M.; Classen, A.K.; Bilder, D. The transcriptional response to tumorigenic polarity loss in Drosophila. eLife 2015, 4, e03189. [Google Scholar] [CrossRef]
- Pascual, J.; Jacobs, J.; Sansores-Garcia, L.; Natarajan, M.; Zeitlinger, J.; Aerts, S.; Halder, G.; Hamaratoglu, F. Hippo Reprograms the Transcriptional Response to Ras Signaling. Dev. Cell 2017, 42, 667–680.e4. [Google Scholar] [CrossRef] [Green Version]
- Conkright, M.D.; Guzmán, E.; Flechner, L.; Su, A.I.; HogenEsch, J.B.; Montminy, M. Genome-Wide Analysis of CREB Target Genes Reveals A Core Promoter Requirement for cAMP Responsiveness. Mol. Cell 2003, 11, 1101–1108. [Google Scholar] [CrossRef]
- Sobala, Ł.F.; Adler, P.N. The Gene Expression Program for the Formation of Wing Cuticle in Drosophila. PLoS Genet. 2016, 12, e1006100. [Google Scholar] [CrossRef] [Green Version]
- Yurchenco, P.D. Basement Membranes: Cell Scaffoldings and Signaling Platforms. Cold Spring Harb. Perspect. Biol. 2010, 3, a004911. [Google Scholar] [CrossRef] [Green Version]
- Velez, A.M.A.; Howard, M.S. Collagen IV in Normal Skin and in Pathological Processes. N. Am. J. Med. Sci. 2012, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Tanjore, H.; Kalluri, R. The Role of Type IV Collagen and Basement Membranes in Cancer Progression and Metastasis. Am. J. Pathol. 2006, 168, 715–717. [Google Scholar] [CrossRef] [Green Version]
- Wood, W.; Jacinto, A. Drosophila melanogaster embryonic haemocytes: Masters of multitasking. Nat. Rev. Mol. Cell Biol. 2007, 8, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Diaz-De-La-Loza, M.-D.-C.; Ray, R.P.; Ganguly, P.S.; Alt, S.; Davis, J.R.; Hoppe, A.; Tapon, N.; Salbreux, G.; Thompson, B. Apical and Basal Matrix Remodeling Control Epithelial Morphogenesis. Dev. Cell 2018, 46, 23–39.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor-Pareja, J.C.; Xu, T. Shaping Cells and Organs in Drosophila by Opposing Roles of Fat Body-Secreted Collagen IV and Perlecan. Dev. Cell 2011, 21, 245–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, M.; Cao, X.; Dai, J.; Pastor-Pareja, J.C. Basement Membrane Manipulation in Drosophila Wing Discs Affects Dpp Retention but Not Growth Mechanoregulation. Dev. Cell 2017, 42, 97–106.e4. [Google Scholar] [CrossRef] [Green Version]
- Chioran, A.; Duncan, S.; Catalano, A.; Brown, T.J.; Ringuette, M. Collagen IV trafficking: The inside-out and beyond story. Dev. Biol. 2017, 431, 124–133. [Google Scholar] [CrossRef]
- Shahab, J.; Baratta, C.; Scuric, B.; Godt, D.; Venken, K.J.; Ringuette, M. Loss of SPARC dysregulates basal lamina assembly to disrupt larval fat body homeostasis inDrosophila melanogaster. Dev. Dyn. 2015, 244, 540–552. [Google Scholar] [CrossRef]
- Isabella, A.J.; Horne-Badovinac, S. Dynamic regulation of basement membrane protein levels promotes egg chamber elongation in Drosophila. Dev. Biol. 2015, 406, 212–221. [Google Scholar] [CrossRef] [Green Version]
- Calvo, F.; Ege, N.; Grande-Garcia, A.; Hooper, S.; Jenkins, R.P.; Chaudhry, S.I.; Harrington, K.; Williamson, P.; Moeendarbary, E.; Charras, G.; et al. Mechanotransduction and YAP-dependent matrix remodelling is required for the generation and maintenance of cancer-associated fibroblasts. Nat. Cell Biol. 2013, 15, 637–646. [Google Scholar] [CrossRef]
- Nardone, G.; La Cruz, J.O.-D.; Vrbsky, J.; Martini, C.; Pribyl, J.; Skládal, P.; Pešl, M.; Caluori, G.; Pagliari, S.; Martino, F.; et al. YAP regulates cell mechanics by controlling focal adhesion assembly. Nat. Commun. 2017, 8, 15321. [Google Scholar] [CrossRef]
- Totaro, A.; Zhuang, Q.; Panciera, T.; Battilana, G.; Azzolin, L.; Brumana, G.; Gandin, A.; Brusatin, G.; Cordenonsi, M.; Piccolo, S. Cell phenotypic plasticity requires autophagic flux driven by YAP/TAZ mechanotransduction. Proc. Natl. Acad. Sci. USA 2019, 116, 17848–17857. [Google Scholar] [CrossRef] [Green Version]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in mechanotransduction. Nat. Cell Biol. 2011, 474, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Panciera, T.; Azzolin, L.; Cordenonsi, M.; Piccolo, S. Mechanobiology of YAP and TAZ in physiology and disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Zanconato, F.; Forcato, M.; Battilana, G.; Azzolin, L.; Quaranta, E.; Bodega, B.; Rosato, A.; Bicciato, S.; Cordenonsi, M.; Piccolo, S. Genome-wide association between YAP/TAZ/TEAD and AP-1 at enhancers drives oncogenic growth. Nat. Cell Biol. 2015, 17, 1218–1227. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Goel, H.L.; Gao, H.; Pursell, B.; Shultz, L.D.; Greiner, D.L.; Ingerpuu, S.; Patarroyo, M.; Cao, S.; Lim, E.; et al. A laminin 511 matrix is regulated by TAZ and functions as the ligand for the α6Bβ1 integrin to sustain breast cancer stem cells. Genes Dev. 2015, 29, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Stein, C.; Bardet, A.F.; Roma, G.; Bergling, S.; Clay, I.; Ruchti, A.; Agarinis, C.; Schmelzle, T.; Bouwmeester, T.; Schuebeler, D.; et al. YAP1 Exerts Its Transcriptional Control via TEAD-Mediated Activation of Enhancers. PLoS Genet. 2015, 11, e1005465. [Google Scholar] [CrossRef] [Green Version]
- Bunt, S.; Hooley, C.; Hu, N.; Scahill, C.; Weavers, H.; Skaer, H. Hemocyte-Secreted Type IV Collagen Enhances BMP Signaling to Guide Renal Tubule Morphogenesis in Drosophila. Dev. Cell 2010, 19, 296–306. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Harris, R.E.; Bayston, L.J.; Ashe, H.L. Type IV collagens regulate BMP signalling in Drosophila. Nat. Cell Biol. 2008, 455, 72–77. [Google Scholar] [CrossRef]
- Kagey, J.D.; Brown, J.A.; Moberg, K.H. Regulation of Yorkie activity in Drosophila imaginal discs by the Hedgehog receptor gene patched. Mech. Dev. 2012, 129, 339–349. [Google Scholar] [CrossRef]
- Oh, H.; Irvine, K.D. Cooperative Regulation of Growth by Yorkie and Mad through bantam. Dev. Cell 2011, 20, 109–122. [Google Scholar] [CrossRef] [Green Version]
- Hariharan, I.K.; Bilder, D. Regulation of Imaginal Disc Growth by Tumor-Suppressor Genes inDrosophila. Annu. Rev. Genet. 2006, 40, 335–361. [Google Scholar] [CrossRef]
- Herranz, H.; Weng, R.; Cohen, S.M. Crosstalk between Epithelial and Mesenchymal Tissues in Tumorigenesis and Imaginal Disc Development. Curr. Biol. 2014, 24, 1476–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brumby, A.M. scribble mutants cooperate with oncogenic Ras or Notch to cause neoplastic overgrowth in Drosophila. EMBO J. 2003, 22, 5769–5779. [Google Scholar] [CrossRef] [PubMed]
- Pocaterra, A.; Romani, P.; Dupont, S. YAP/TAZ functions and their regulation at a glance. J. Cell Sci. 2020, 133, jcs230425. [Google Scholar] [CrossRef] [PubMed]
- Borreguero-Muñoz, N.; Fletcher, G.C.; Aguilar-Aragon, M.; Elbediwy, A.; Vincent-Mistiaen, Z.I.; Thompson, B. The Hippo pathway integrates PI3K–Akt signals with mechanical and polarity cues to control tissue growth. PLoS Biol. 2019, 17, e3000509. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Reilein, A.; Kalderon, D. Yorkie and Hedgehog independently restrict BMP production in escort cells to permit germline differentiation in the Drosophila ovary. Development 2017, 144, 2584–2594. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO Terms Enriched in YkiS168A-Expressing Discs (Relative to Control) | |
| Upregulated Genes (Biological Process) | −Log2 p-Value |
| Cytoplasmic translation | 130.2 |
| Cellular process | 80.1 |
| Cellular metabolic process | 64.9 |
| Single organism cellular process | 59.9 |
| Organonitrogen compound metabolic process | 54.8 |
| Upregulated Genes (Molecular Function) | −Log2 p-Value |
| Structural constituent of ribosome | 56.3 |
| Structural molecule activity | 25.8 |
| Cofactor binding | 23.8 |
| Translation factor activity; RNA binding | 15.7 |
| Translation initiation factor activity | 13.6 |
| Downregulated Genes (Biological Process) | −Log2 p-Value |
| System development | 179.7 |
| Regulation of biological process | 175.9 |
| Regulation of cellular process | 172.9 |
| Biological regulation | 172.6 |
| Animal organ development | 169.8 |
| Downregulated Genes (Molecular Function) | −Log2 p-Value |
| Sequence-specific DNA binding | 36.3 |
| Nucleic acid binding transcription factor activity | 35.8 |
| Transcription factor activity, sequence-specific DNA binding | 35.8 |
| mRNA binding | 33.4 |
| RNA polymerase II transcription factor activity, sequence-specific DNA binding | 32.6 |
| ECM-Related Genes Altered in YkiS168A-Expressing Discs (Relative to Control) | |
| Gene | Annotated Function (FlyBase.org) |
| Upregulated | |
| dpy | ECM protein involved in cuticle attachment |
| mmy | Protein glycosylase involved in ECM and cuticle construction |
| wb | Laminin-α chain ECM protein; BM integrity and integrin signaling |
| serp | Secreted apical ECM protein involved in cuticle construction |
| trol | Perlecan ECM secreted heparan sulfate proteoglycan |
| verm | Chitin deacetylase involved in cuticle development |
| Timp | Inhibitor of matrix metalloproteinases, regulates ECM remodeling |
| by | Tensin, binds integrin/actin to regulate wing surface adhesion |
| mgl | Apical membrane protein, cuticle development |
| Ndg | Structural ECM protein, organizes BM assembly |
| Mmp1 | Matrix metalloproteinase, regulates ECM remodeling |
| mfas | Regulator of cell adhesion and hemocyte proliferation |
| sona | ADAM protease in ECM, positive regulator of tissue growth |
| mspo | Spondin family ECM glycoprotein |
| Crag | Rab exchange factor involved in BM protein secretion |
| scaf | Inactive serine protease, regulates Laminin localization in BM |
| Gasp | Regulator of cuticle biosynthesis |
| Cpr47Eb | Structural component of cuticle |
| if | Integrin α-subunit of receptor for ECM |
| Itgbn | Integrin β-subunit of receptor for ECM |
| Vinc | Vinculin; mechanotransducer in cell-matrix interactions |
| stck | Integrin adaptor protein, wing disc apposition |
| plx | RabGAP, regulates integrin-mediated adhesion |
| Pax | Paxillin; integrin adaptor |
| Itgbn | |
| Downregulated | |
| Dg | Non-integrin ECM-actin adapting receptor |
| dlp | Glypican ECM membrane heparan sulfate proteoglycan |
| Glt | Secreted component of BM |
| LanB2 | Laminin-B2; BM component interacts with ECM and integrins |
| Mmp2 | Metalloproteinase, regulates ECM remodeling |
| Mp | Collagen XV/XVIII member |
| Ppn | ADAM-like protein, regulates ECM construction |
| Pxn | Peroxidase, regulates BM assembly |
| scb | α-PS3 Integrin, ECM ligand function |
| SPARC | Collagen and ECM-binding protein; BM maturation |
| Tig | Integrin ligand component of ECM |
| Tsp | Integrin ligand component of ECM |
| GO Terms Enriched in mudRNAi-Expressing Discs (Relative to Control) | |
| Upregulated Genes (Biological Process) | −Log2 p-Value |
| Cytoplasmic translation | 69.6 |
| Organic substance biosynthetic process | 23.4 |
| Regulation of cellular protein localization | 22.5 |
| Biosynthetic process | 21.4 |
| Cellular biosynthetic process | 21.3 |
| Upregulated Genes (Molecular Function) | −Log2 p-Value |
| Structural constituent of ribosome | 27.2 |
| Structural molecule activity | 21.5 |
| rRNA binding | 16.3 |
| Unfolded protein binding | 10.1 |
| ATP binding | 10.0 |
| Downregulated Genes (Biological Process) | −Log2 p-Value |
| Tissue development | 26.5 |
| Negative regulation of translation | 25.0 |
| Negative regulation of cellular amide metabolic process | 23.9 |
| Response to stimulus | 23.8 |
| Single-multicellular organism process | 21.8 |
| Downregulated Genes (Molecular Function) | −Log2 p-Value |
| Nutrient reservoir activity | 18.6 |
| mRNA 3′-UTR binding | 13.0 |
| Receptor binding | 11.4 |
| Translation repressor activity | 11.3 |
| Nucleic acid binding transcription factor activity | 11.1 |
| GO Terms Enriched in mudRNAi;YkiS168A-Expressing Discs (Relative to YkiS168A) | |
| Upregulated Genes (Biological Process) | −Log2 p-Value |
| Apoptotic process | 10.5 |
| Programmed cell death | 7.9 |
| Regulation of apoptotic process | 7.7 |
| Cell death | 7.5 |
| Regulation of programmed cell death | 7.2 |
| Upregulated Genes (Molecular Function) | −Log2 p-Value |
| Double-stranded RNA-specific ribonuclease activity | 6.5 |
| Nuclease activity | 5.5 |
| Catalytic activity | 4.4 |
| Downregulated Genes (Biological Process) | −Log2 p-Value |
| Post-embryonic development | 12.3 |
| Chitin-based cuticle development | 10.4 |
| Striated muscle cell differentiation | 9.3 |
| Multicellular organism development | 9.2 |
| Anatomical structure development | 8.9 |
| Downregulated Genes (Molecular Function) | −Log2 p-Value |
| Structural constituent of chitin-based larval cuticle | 9.1 |
| Extracellular matrix structural constituent | 9.0 |
| Structural molecule activity | 8.4 |
| Structural constituent of chitin-based cuticle | 8.1 |
| Structural constituent of cuticle | 7.7 |
| ECM-Related Genes Downregulated in mudRNAi;YkiS168A-Expressing Discs (Relative to YkiS168A Alone) | |
| Gene | Annotated Function (FlyBase.org) |
| vkg | Type IV collagen; core BM component |
| Col4a1 | Type IV collagen; core BM component |
| tnc | Structural component of ECM/collagen |
| pot | Transmembrane protein component of apical ECM; links epithelia to cuticle and organizes microtubules |
| Glt | Secreted glycoprotein component of BM |
| kirre | Transmembrane adhesion protein and ECM interactor |
| Cpr49Ac | Cuticle component |
| Lcp1 | Structural component of cuticle |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parra, A.S.; Johnston, C.A. Mud Loss Restricts Yki-Dependent Hyperplasia in Drosophila Epithelia. J. Dev. Biol. 2020, 8, 34. https://doi.org/10.3390/jdb8040034
Parra AS, Johnston CA. Mud Loss Restricts Yki-Dependent Hyperplasia in Drosophila Epithelia. Journal of Developmental Biology. 2020; 8(4):34. https://doi.org/10.3390/jdb8040034
Chicago/Turabian StyleParra, Amalia S., and Christopher A. Johnston. 2020. "Mud Loss Restricts Yki-Dependent Hyperplasia in Drosophila Epithelia" Journal of Developmental Biology 8, no. 4: 34. https://doi.org/10.3390/jdb8040034
APA StyleParra, A. S., & Johnston, C. A. (2020). Mud Loss Restricts Yki-Dependent Hyperplasia in Drosophila Epithelia. Journal of Developmental Biology, 8(4), 34. https://doi.org/10.3390/jdb8040034