
In Utero Programming of Testicular Cancer


{kind=link}
Abstract
:1. Introduction
2. Testicular Germ Cell Cancer
3. Testicular Cancer in Young Adults
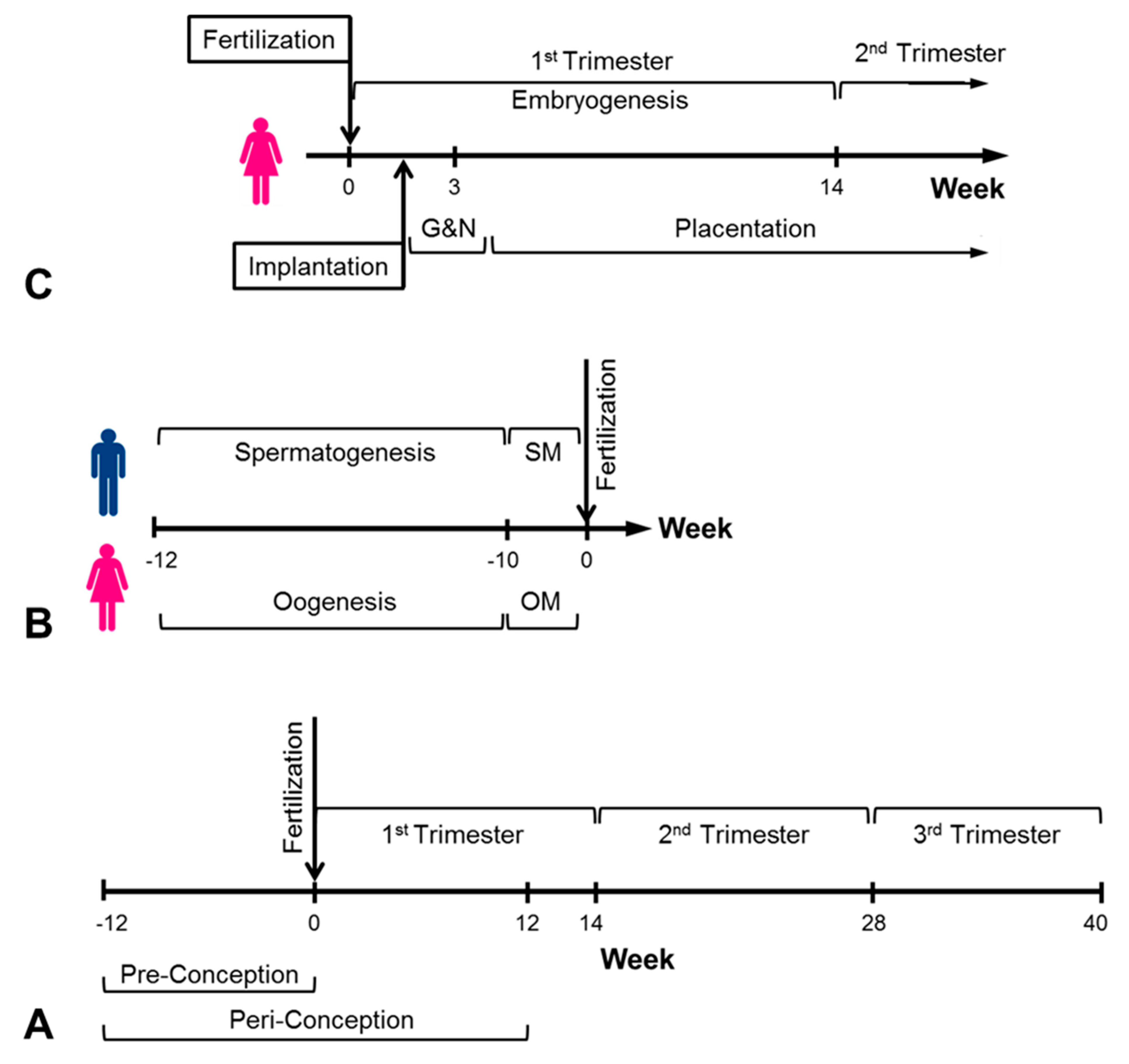
4. In Utero Conditions and Cancer
5. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mahabir, S.; Aagaard, K.; Anderson, L.M.; Herceg, Z.; Hiatt, R.A.; Hoover, R.N.; Linet, M.S.; Medina, D.; Potischman, N.; Tretli, S.; et al. Challenges and opportunities in research on early-life events/exposures and cancer development later in life. Cancer Causes Control 2012, 23, 983–990. [Google Scholar] [CrossRef]
- Stephansson, O.; Wahnström, C.; Pettersson, A.; Sørensen, H.T.; Tretli, S.; Gissler, M.; Troisi, R.; Akre, O.; Grotmol, T. Perinatal risk factors for childhood testicular germ-cell cancer: A Nordic population-based study. Cancer Epidemiol. 2012, 35, e100–e104. [Google Scholar] [CrossRef]
- Faruqui, N.; Kummrow, A.; Fu, B.; Divieto, C.; Rojas, F.; Kisulu, F.; Cavalcante, J.; Wang, J.; Campbell, J.; Martins, J.L.; et al. Cellular Metrology: Scoping for a Value Proposition in Extra- and Intracellular Measurements. Front. Bioeng. Biotechnol. 2020, 7, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, E.J.; Kim, Y.J. What is fetal programming? A lifetime health is under the control of in utero health. Obstet. Gynecol. Sci. 2017, 60, 506–519. [Google Scholar] [CrossRef] [PubMed]
- Duncan, J.M. On puerperal diabetes. Trans. Obstet. Soc. Lob. 1882, 24, 256–285. [Google Scholar]
- Lecture, F.N.B. Of pregnancy and progeny. Diabetes 1980, 29, 1023–1035. [Google Scholar]
- Strauss, R.S. Effects of the intrauterine environment on childhood growth. Br. Med Bull. 1997, 53, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Gluckman, P.D.; Hanson, M.; Cooper, C.; Thornburg, K. Effect of In Utero and Early-Life Conditions on Adult Health and Disease. N. Engl. J. Med. 2008, 359, 61–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzahor, E.; Poss, K.D. Cardiac regeneration strategies: Staying young at heart. Science 2017, 356, 1035–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.S.; Kim, J.; Elghiaty, A.; Ham, W.S. Recent global trends in testicular cancer incidence and mortality. Medicine 2018, 97, e12390. [Google Scholar] [CrossRef]
- Gurney, J.K.; Florio, A.A.; Znaor, A.; Ferlay, J.; Laversanne, M.; Sarfati, D.; Bray, F.; McGlynn, K.A. International Trends in the Incidence of Testicular Cancer: Lessons from 35 Years and 41 Countries. Eur. Urol. 2019, 76, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Harper, A.; Ruan, Y.; Barr, R.; Frazier, A.L.; Ferlay, J.; Steliarova-Foucher, E.; Fidler-Benaoudia, M.M. International Trends in the Incidence of Cancer Among Adolescents and Young Adults. J. Natl. Cancer Inst. 2020, 112, 1105–1117. [Google Scholar] [CrossRef]
- Moch, H.; Cubilla, A.L.; Humphrey, P.A.; Reuter, V.E.; Ulbright, T.M. The 2016 WHO Classification of Tumours of the Urinary System and Male Genital Organs—Part A: Renal, Penile, and Testicular Tumours. Eur. Urol. 2016, 70, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Dorssers, L.C.J.; Gillis, A.J.M.; Stoop, H.; Van Marion, R.; Nieboer, M.M.; van Riet, J.; Van De Werken, H.J.; Oosterhuis, J.W.; De Ridder, J.; Looijenga, L.H.J. Molecular heterogeneity and early metastatic clone selection in testicular germ cell cancer development. Br. J. Cancer 2019, 120, 444–452. [Google Scholar] [CrossRef] [Green Version]
- Oosterhuis, J.W.; Looijenga, L. Testicular germ-cell tumours in a broader perspective. Nat. Rev. Cancer 2005, 5, 210–222. [Google Scholar] [CrossRef]
- Ulbright, T.M. Recently Described and Clinically Important Entities in Testis Tumors: A Selective Review of Changes Incorporated Into the 2016 Classification of the World Health Organization. Arch. Pathol. Lab. Med. 2019, 143, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Idrees, M.T.; Kao, C.S.; Epstein, J.I.; Ulbright, T.M. Nonchoriocarcinomatous Trophoblastic Tumors of the Testis: The Widening Spectrum of Trophoblastic Neoplasia. Am. J. Surg. Pathol. 2015, 39, 1468–1478. [Google Scholar] [CrossRef]
- Razafimahefa, J.; Gosset, C.; Culine, S.; Mongiat-Artus, P.; Verine, J. Placental Site Trophoblastic Tumor in Nonseminomatous Mixed Germ Cell Tumors of the Testis: A Case Report and Review of the Literature. Clin. Genitourin. Cancer 2018, 16, e349–e354. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zheng, W.; Liang, Q.; Yin, T. Diagnosis and treatment of placental site trophoblastic tumor. Int. J. Clin. Exp. Pathol. 2013, 6, 1448–1451. [Google Scholar]
- Baird, D.C.; Meyers, G.J.; Hu, J.S. Testicular Cancer: Diagnosis and Treatment. Am. Fam. Physician 2018, 97, 261–268. [Google Scholar] [PubMed]
- Saltzman, A.F.; Cost, N.G. Adolescent and Young Adult Testicular Germ Cell Tumors: Special Considerations. Adv. Urol. 2018, 31, 2375176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghazarian, A.A.; Kelly, S.P.; Altekruse, S.F.; Rosenberg, P.S.; McGlynn, K.A. Future of testicular germ cell tumor incidence in the United States: Forecast through 2026. Cancer 2017, 123, 2320–2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulytė, A.; Ulys, A.; Sužiedėlis, K.; Patašius, A.; Smailytė, G. Testicular cancer in two brothers of a quadruplet: A case report and a review of literature. Acta Med. Litu. 2017, 24, 12–17. [Google Scholar] [CrossRef]
- Grotmol, T.; Weiderpass, E.; Tretli, S. Conditions in utero and cancer risk. Eur. J. Epidemiol. 2006, 21, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Ekbom, A. Growing evidence that several human cancers may originate in utero. Semin. Cancer Biol. 1998, 8, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Timms, B.G.; Howdeshell, K.L.; Barton, L.; Bradley, S.; Richter, C.; Saal, F.S.V. Estrogenic chemicals in plastic and oral contraceptives disrupt development of the fetal mouse prostate and urethra. Proc. Natl. Acad. Sci. USA 2005, 102, 7014–7019. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, L.M.; Zava, D.; McGlynn, K.A.; Stanczyk, F.Z.; Kang, A.Y.; Ma, X.; Wiemels, J.L.; Metayer, C. Neonatal Hormone Concentrations and Risk of Testicular Germ Cell Tumors (TGCT). Cancer Epidemiol. Biomark. Prev. 2018, 27, 488–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, M.B.; Akre, O.; Forman, D.; Madigan, M.P.; Richiardi, L.; McGlynn, K.A. A systematic review and meta-analysis of perinatal variables in relation to the risk of testicular cancer—Experiences of the son. Int. J. Epidemiol. 2010, 39, 1605–1618. [Google Scholar] [CrossRef] [Green Version]
- Myatt, L. Placental adaptive responses and fetal programming. J. Physiol. 2006, 572, 25–30. [Google Scholar] [CrossRef]
- Desoye, G. The Human Placenta in Diabetes and Obesity: Friend or Foe? The 2017 Norbert Freinkel Award Lecture. Diabetes Care 2018, 41, 1362–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fall, C.H.D.; Kumaran, K. Metabolic programming in early life in humans. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.N.; Tain, Y.L. The good, the bad, and the ugly of pregnancy nutrients and developmental programming of adult disease. Nutrients 2019, 11, 894. [Google Scholar] [CrossRef] [Green Version]
- Oosterhuis, J.W.; Looijenga, L.H.J. Human germ cell tumours from a developmental perspective. Nat. Rev. Cancer 2019, 19, 522–537. [Google Scholar] [CrossRef]
- Christoforou, E.R.; Sferruzzi-Perri, A.N. Molecular mechanisms governing offspring metabolic programming in rodent models of in utero stress. Cell Mol. Life Sci. 2020, 77, 4861–4898. [Google Scholar] [CrossRef]
- Stanner, S.A.; Yudkin, J.S. Fetal programming and the Leningrad Siege study. Twin Res. 2001, 4, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Painter, R.C.; Roseboom, T.J.; Bleker, O.P. Prenatal exposure to the Dutch famine and disease in later life: An overview. Reprod. Toxicol. 2005, 20, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Roseboom, T.J.; Painter, R.C.; van Abeelen, A.F.; Veenendaal, M.V.; de Rooij, S.R. Hungry in the womb: What are the consequences? Lessons from the Dutch famine. Maturitas 2011, 70, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Bianco-Miotto, T.; Craig, J.M.; Gasser, Y.P.; van Dijk, S.J.; Ozanne, S.E. Epigenetics and DOHaD: From basics to birth and beyond. J. Dev. Orig. Health Dis. 2017, 8, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Heijmans, B.T.; Tobi, E.; Stein, A.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar] [CrossRef] [Green Version]
- Tobi, E.; Goeman, J.; Monajemi, R.; Gu, H.; Putter, H.; Zhang, Y.; Slieker, R.; Stok, A.P.; Thijssen, P.E.; Müller, F.; et al. DNA methylation signatures link prenatal famine exposure to growth and metabolism. Nat. Commun. 2014, 5, 5592. [Google Scholar] [CrossRef] [Green Version]
- Amatruda, J.F.; Ross, J.A.; Christensen, B.; Fustino, N.J.; Chen, K.S.; Hooten, A.J.; Nelson, H.; Kuriger, J.K.; Rakheja, D.; Frazier, A.L.; et al. DNA methylation analysis reveals distinct methylation signatures in pediatric germ cell tumors. BMC Cancer 2013, 13, 313. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Fernandez, J.; Ochoa, J.J.; Lopez-Frias, M.; Diaz-Castro, J. Impact of Early Nutrition, Physical Activity and Sleep on the Fetal Programming of Disease in the Pregnancy: A Narrative Review. Nutrients 2020, 12, 3900. [Google Scholar] [CrossRef] [PubMed]
- Elad, D.; Jaffa, A.J.; Grisaru, D. Biomechanics of Early Life in the Female Reproductive Tract. Physiology 2020, 35, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Takakuwa, T. 3D Analysis of Human Embryos and Fetuses Using Digitized Datasets From the Kyoto Collection. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2018, 301, 960–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padhee, M.; Zhang, S.; Lie, S.; Wang, K.C.; Botting, K.J.; McMillen, I.C.; MacLaughlin, S.M.; Morrison, J.L. The Periconceptional Environment and Cardiovascular Disease: Does In Vitro Embryo Culture and Transfer Influence Cardiovascular Development and Health? Nutrients 2015, 7, 1378–1425. [Google Scholar] [CrossRef] [PubMed]
- van Uitert, E.M.; Van Der Elst-Otte, N.; Wilbers, J.J.; Exalto, N.; Willemsen, S.P.; Eilers, P.H.C.; Koning, A.H.J.; Steegers, E.A.P.; Steegers-Theunissen, R.P.M. Periconception maternal characteristics and embryonic growth trajectories: The Rotterdam Predict study. Hum. Reprod. 2013, 28, 3188–3196. [Google Scholar] [CrossRef] [Green Version]
- Louis, G.M.; Cooney, M.A.; Lynch, C.D.; Handal, A. Periconception window: Advising the pregnancy-planning couple. Fertil. Steril. 2008, 89, e119–e121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafizi, L.; Asgarieh, E.R.; Taheri, N.; Ghomian, N. Successful Management of Spontaneous Quadruplet Pregnancy: A Case Report. J. Fam. Reprod. Health 2018, 12, 173–176. [Google Scholar]
- Schlueter, R.; Arnett, C.; Huang, C.; Burlingame, J. Successful quintuplet pregnancy of monochorionic male quadruplets and single female after double embryo transfer: Case report and review of the literature. Fertil. Steril. 2018, 109, 284–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baroni, T.; Arato, I.; Mancuso, F.; Calafiore, R.; Luca, G. On the Origin of Testicular Germ Cell Tumors: From Gonocytes to Testicular Cancer. Front. Endocrinol. 2019, 10, 343. [Google Scholar] [CrossRef]
- Das, M.K.; Kleppa, L.; Haugen, T.B. Functions of genes related to testicular germ cell tumour development. Andrology 2019, 7, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Bo, H.; Cao, K.; Tang, R.; Zhang, H.; Gong, Z.; Liu, Z.; Liu, J.; Li, J.; Fan, L. A network-based approach to identify DNA methylation and its involved molecular pathways in testicular germ cell tumors. J. Cancer 2019, 10, 893–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, K.M.; Elad, D. Biomechanics of the human uterus. Wiley Interdiscip. Rev. Syst. Biol. Med. 2017, 9, e1388. [Google Scholar] [CrossRef]
- Mammoto, T.; Mammoto, A.; Ingber, D.E. Mechanobiology and Developmental Control. Annu. Rev. Cell Dev. Biol. 2013, 29, 27–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Noble, F.; Moyon, D.; Pardanaud, L.; Yuan, L.; Djonov, V.; Matthijsen, R.; Bréant, C.; Fleury, V.; Eichmann, A. Flow regulates arterial-venous differentiation in the chick embryo yolk sac. Development 2004, 131, 361–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hove, J.R.; Köster, R.W.; Forouhar, A.S.; Acevedo-Bolton, G.; Fraser, S.E.; Gharib, M. Intracardiac fluid forces are an essential epigenetic factor for embryonic cardiogenesis. Nature 2003, 421, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Hove, J.R. Quantifying cardiovascular flow dynamics during early development. Ped. Res. 2006, 60, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Critchley, H.O.D.; Maybin, J.A.; Armstrong, G.M.; Williams, A.R.W. Physiology of the Endometrium and Regulation of Menstruation. Physiol. Rev. 2020, 100, 1149–1179. [Google Scholar] [CrossRef]
- Kamm, R.D.; Bashir, R.; Arora, N.; Dar, R.D.; Gillette, M.U.; Griffith, L.G.; Kemp, M.L.; Kinlaw, K.; Levin, M.; Martin, A.C.; et al. Perspective: The promise of multicellular engineered living systems. APL Bioeng. 2018, 2, 040901. [Google Scholar] [CrossRef] [Green Version]
- Biechonski, S.; Gourevich, D.; Rall, M.; Aqaqe, N.; Yassin, M.; Zipin-Roitman, A.; Trakhtenbrot, L.; Olender, L.; Raz, Y.; Jaffa, A.J.; et al. Quercetin alters the DNA damage response in human hematopoietic cells via TopoII- and PI3K- dependent mechanisms synergizing in leukemogenic rearrangements. Int. J. Cancer 2017, 140, 864–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makoolati, Z.; Movahedin, M.; Forouzandeh-Moghadam, M. Proliferation in culture of primordial germ cells derived from embryonic stem cell: Induction by retinoic acid. Biosci. Rep. 2016, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gell, J.; Liu, W.; Sosa, E.; Chialastri, A.; Hancock, G.; Tao, Y.; Wamaitha, S.; Bower, G.; Dey, S.S.; Clark, A.T. An Extended Culture System that Supports Human Primordial Germ Cell-like Cell Survival and Initiation of DNA Methylation Erasure. Stem Cell Rep. 2020, 14, 433–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elad, D.; Jaffa, A.J.; Grisaru, D.; Leibovitch, I. In Utero Programming of Testicular Cancer. J. Dev. Biol. 2021, 9, 35. https://doi.org/10.3390/jdb9030035
Elad D, Jaffa AJ, Grisaru D, Leibovitch I. In Utero Programming of Testicular Cancer. Journal of Developmental Biology. 2021; 9(3):35. https://doi.org/10.3390/jdb9030035
Chicago/Turabian StyleElad, David, Ariel J. Jaffa, Dan Grisaru, and Ilan Leibovitch. 2021. "In Utero Programming of Testicular Cancer" Journal of Developmental Biology 9, no. 3: 35. https://doi.org/10.3390/jdb9030035
APA StyleElad, D., Jaffa, A. J., Grisaru, D., & Leibovitch, I. (2021). In Utero Programming of Testicular Cancer. Journal of Developmental Biology, 9(3), 35. https://doi.org/10.3390/jdb9030035