Use of Hyperspectral Reflectance Sensing for Assessing Growth and Chlorophyll Content of Spring Wheat Grown under Simulated Saline Field Conditions



 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Experimental Description
2.2. Salinity Treatments, Experimental Design, and Agronomic Practices
2.3. Hyperspectral Reflectance Measurements
2.4. Photosynthetic Pigments and Plant Dry Weight Measurements
2.5. Published and Newly Constructed Spectral Reflectance Indices (SRIs)
2.6. Data Analysis
3. Results
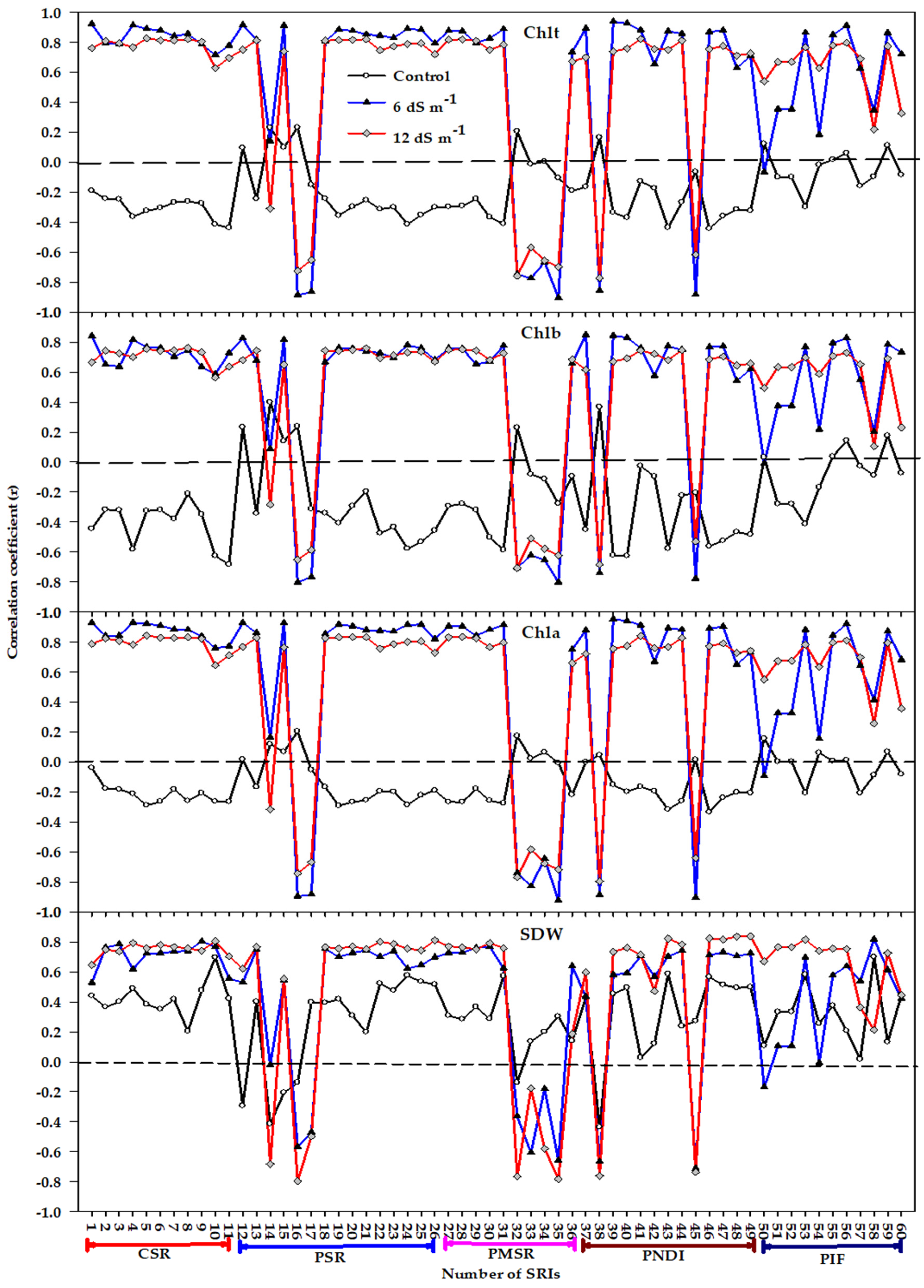
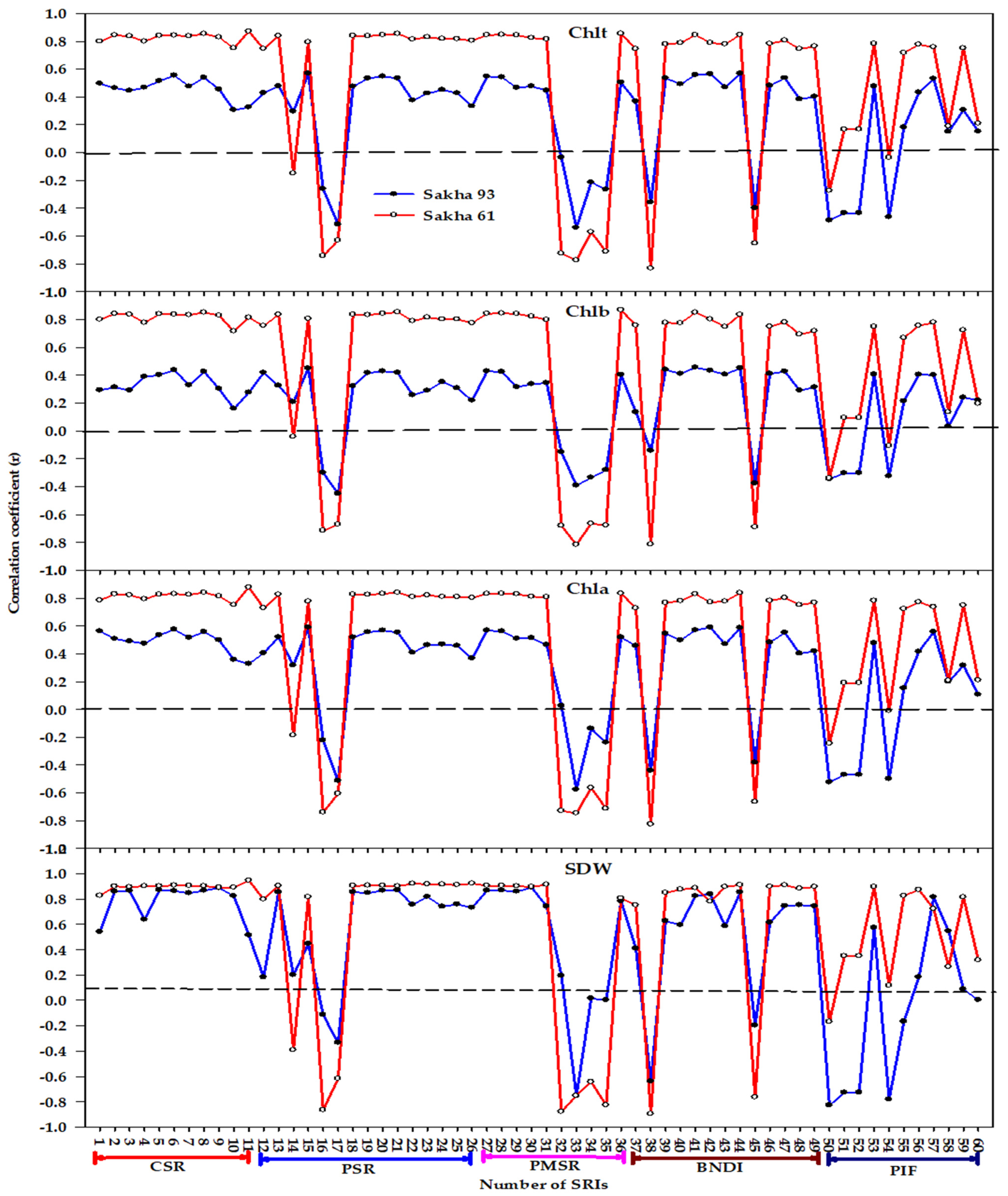
3.1. Influence of Salinity Level, Genotype, and Their Interaction on Biomass, Chlorophyll Content, and Spectral Reflectance Indices
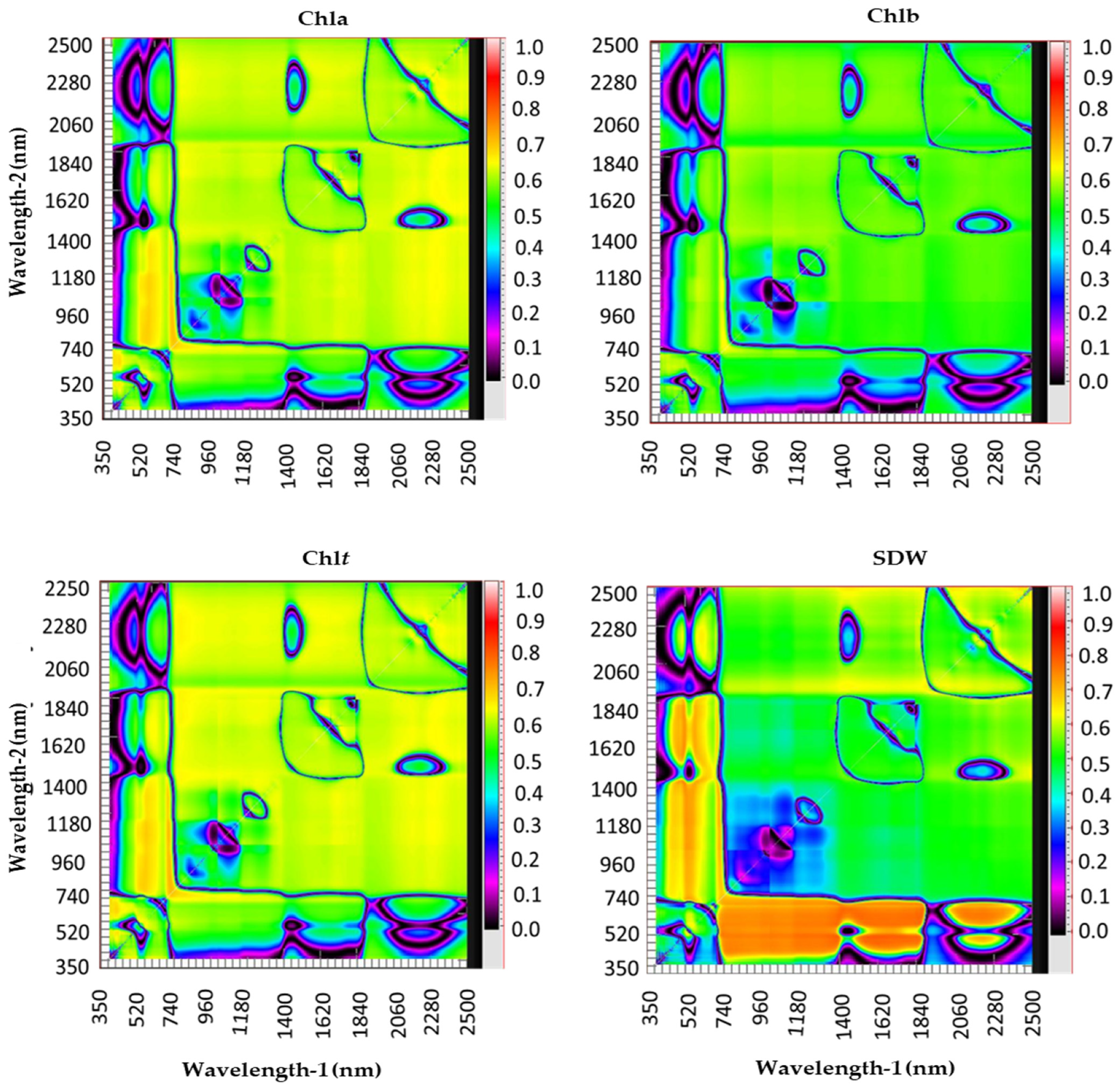
3.2. Relationship between Measured Variables and Different Types of Spectral Reflectance Index
3.3. Relative Importance of Spectral Reflectance Indices in Predicting Measured Variables
3.4. Validation of Predictive Models for Measured Variables Based on Influential SRIs in Each SRI Type
4. Discussion
4.1. Performance of Different Types of Spectral Reflectance Index under Different Growth Conditions
4.2. Validation of Predictive Models for Assessing Variables under Different Growth Conditions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- El-Hendawy, S.E.; Hassan, W.M.; Al-Suhaibani, N.A.; Schmidhalter, U. Spectral assessment of drought tolerance indices and grain yield in advanced spring wheat lines grown under full and limited water irrigation. Agric. Water Manag. 2017, 182, 1–12. [Google Scholar] [CrossRef]
- Al-Omran, A.M.; Falatah, A.M.; Al-Matrood, S.S. Evaluation of irrigation well water quality in Riyadh region, Saudi Arabia. J. King Abdulaziz Univ. 2005, 16, 23–40. [Google Scholar] [CrossRef]
- He, K.; Yang, Y.; Yang, Y.; Chen, S.; Hu, Q.; Liu, X.; Gao, F. Hydrus simulation of sustainable brackish water irrigation in a winter wheat-summer maize rotation system in the North China Plain. Water 2017, 9, 536. [Google Scholar]
- Wang, H.; Xu, Z.; Pang, G.; Zhang, L.; Wang, X. Effects of brackish water irrigation on water-salt distribution and winter wheat growth. J. Soil Water Conserv. 2017, 31, 291–297. [Google Scholar]
- Mansour, E.; Moustafa, E.S.A.; Desoky, E.M.; Ali, M.M.A.; Yasin, M.A.T.; Attia, A.; Alsuhaibani, N.; Tahir, M.U.; El-Hendawy, S.E. Multidimensional evaluation for detecting salt tolerance of bread wheat genotypes under actual saline field growing conditions. Plants 2020, 9, 1324. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salt tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- El-Hendawy, S.E.; Hassan, W.M.; Al-Suhaibani, N.A.; Refay, Y.; Abdella, K.A. Comparative performance of multivariable agro-physiological parameters for detecting salt tolerance of wheat cultivars under simulated saline field growing conditions. Front. Plant. Sci. 2017, 8, 435. [Google Scholar] [CrossRef] [Green Version]
- Oyiga, B.C.; Sharma, R.; Shen, J.; Baum, M.; Ogbonnaya, F.; Léon, J.; Ballvora, A. Identification and characterization of salt tolerance of wheat germplasm using a multivariable screening approach. J. Agron. Crop Sci. 2016, 202, 472–485. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Hoque, M.A.; Banu, M.N.A.; Okuma, E.; Amako, K.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Exogenous proline and glycinebetaine increase NaCl-induced ascorbate–glutathione cycle enzyme activities, and proline improves salt tolerance more than glycinebetaine in tobacco Bright Yellow-2 suspension-cultured cells. J. Plant Physiol. 2007, 164, 1457–1468. [Google Scholar]
- Taïbi, K.; Taïbi, F.; Abderrahim, L.A.; Ennajah, A.; Belkhodja, M.; Mulet, J.M. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defense systems in Phaseolus vulgaris L. S. Afr. J. Bot. 2016, 105, 306–312. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Garriga, M.; Retamales, J.B.; Romero, S.; Caligari, P.D.S.; Lobos, G.A. Chlorophyll, anthocyanin, and gas exchange changes assessed by spectroradiometry in Fragaria chiloensis under salt stress. J. Integr. Plant. Biol. 2014, 56, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Croft, H.; Chen, J.M.; Luo, X.; Bartlett, P.; Chen, B.; Staebler, R.M. Leaf chlorophyll content as a proxy for leaf photosynthetic capacity. Glob. Chang. Biol. 2017, 23, 3513–3524. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Huang, W.; Zhang, J.; Kong, W.; Casa, R.; Huang, Y. A novel combined spectral index for estimating the ratio of carotenoid to chlorophyll content to monitor crop physiological and phenological status. Int. J. Appl. Earth Obs. 2019, 76, 128–142. [Google Scholar] [CrossRef]
- Shah, S.H.; Houborg, M.; McCabe, M.F. Response of Chlorophyll, Carotenoid and SPAD-502 Measurement to Salinity and Nutrient Stress in Wheat (Triticum aestivum L.). Agronomy 2017, 7, 61. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Liu, L. Retrieving crop leaf chlorophyll content using an improved look-up-table approach by combining multiple canopy structures and soil backgrounds. Remote Sens. 2020, 12, 2139. [Google Scholar] [CrossRef]
- Richardson, A.D.; Duigan, S.; Berlyn, G. An evaluation of noninvasive methods to estimate foliar chlorophyll content. New Phytol. 2002, 153, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Wang, P.; Zhao, K.; Yang, Y.; Yao, S.; Jiang, D. Characteristics of gas exchange and chlorophyll fluorescence in different position leaves at booting stage in rice plants. Rice Sci. 2004, 11, 283–289. [Google Scholar]
- Kong, W.; Huang, W.; Casa, R.; Zhou, X.; Ye, H.; Dong, Y. Off-nadir hyperspectral sensing for estimation of vertical profile of leaf chlorophyll content within wheat canopies. Sensors 2017, 17, 2711. [Google Scholar] [CrossRef] [Green Version]
- Marenco, R.A.; Antezana-Vera, S.A.; Nascimento, H.C.S. Relationship between specific leaf area, leaf thickness, leaf water content and SPAD—502 readings in six Amazonian tree species. Photosynthetica 2009, 47, 184–190. [Google Scholar] [CrossRef]
- Uddling, J.; Gelang-Alfredsson, J.; Piikki, K.; Pleijel, H. Evaluating the relationship between leaf chlorophyll concentration and spad-502 chlorophyll meter readings. Photosynth. Res. 2007, 91, 37–46. [Google Scholar] [CrossRef]
- Lu, S.; Lu, F.; You, W.; Wang, Z.; Liu, Y.; Omasa, K. A robust vegetation index for remotely assessing chlorophyll content of dorsiventral leaves across several species in different seasons. Plant Methods 2018, 14, 2–15. [Google Scholar] [CrossRef] [Green Version]
- El-Hendawy, S.E.; Al-Suhaibani, N.; Dewir, Y.H.; El-Sayed, S.; Alotaibi, M.; Hassan, W.M.; Refay, Y.; Tahir, M.U. Ability of modified spectral reflectance indices for estimating growth and photosynthetic efficiency of wheat under saline field conditions. Agronomy 2019, 9, 35. [Google Scholar] [CrossRef] [Green Version]
- Lobos, G.A.; Escobar-Opazo, A.; Estrada, F.; Romero-Bravo, S.; Garriga, M.; del Pozo, A.; Poblete-Echeverría, C.; Gonzalez-Talice, J.; González-Martinez, L.; Caligar, P. Spectral reflectance modeling by wavelength selection: Studying the scope for blueberry physiological breeding under contrasting water supply and heat conditions. Remote Sens. 2019, 11, 329. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Bu, Z.; Lu, S. Estimating chlorophyll content of leafy green vegetables from adaxial and abaxial reflectance. Sensors 2019, 19, 4059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Hendawy, S.E.; Al-Suhaibani, N.; Al-Ashkar, I.; Alotaibi, M.; Tahir, M.U.; Solieman, T.; Hassan, W.M. Combining genetic analysis and multivariate modeling to evaluate spectral reflectance indices as indirect selection tools in wheat breeding under water deficit stress conditions. Remote Sens. 2019, 9, 1480. [Google Scholar] [CrossRef]
- Elmetwalli, A.H.; El-Hendawy, S.E.; Al-Suhaibani, N.; Alotaibi, M.; Tahir, M.U.; Mubushar, M.; Hassan, W.M.; El-Sayed, S. Potential of hyperspectral and thermal proximal sensing for estimating growth performance and yield of soybean exposed to different drip irrigation regimes under arid conditions. Sensors 2020, 20, 6569. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Broge, N.H.; Leblanc, E. Comparing prediction power and stability of broadband and hyperspectral vegetation indices for estimation of green leaf area index and canopy chlorophyll density. Remote Sens Environ. 2001, 76, 156–172. [Google Scholar] [CrossRef]
- Wu, C.Y.; Niu, Z.; Tang, Q.; Huang, W.J. Estimating chlorophyll content from hyperspectral vegetation indices: Modeling and validation. Agric. For. Meteorol. 2008, 148, 1230–1241. [Google Scholar] [CrossRef]
- Pu, R.; Gong, P. Hyperspectral remote sensing of vegetation bioparameters. In Advances in Environmental Remote Sensing Sensors, Algorithms, and Applications; CRC Press: Boca Raton, FL, USA, 2011; ISBN 9780429143687. [Google Scholar] [CrossRef]
- Liang, S.; Li, X.; Wang, J. Advanced Remote Sensing; Academic Press: Cambridge, MA, USA, 2012; ISBN 978-0-12-815826-5. [Google Scholar] [CrossRef]
- Seelig, H.D.; Hoehn, A.; Stodieck, L.S.; Klaus, D.M.; Adams III, W.W.; Emery, W.J. Relations of remote sensing leaf water indices to leaf water thickness in cowpea, bean, and sugar beet plants. Remote Sens. Environ. 2008, 112, 445–455. [Google Scholar] [CrossRef]
- Wang, J.; Xu, R.; Yang, S. Estimation of plant water content by spectral absorption features centered at 1450 nm and 1940 nm regions. Environ. Monit. Assess. 2009, 157, 459–469. [Google Scholar] [CrossRef] [PubMed]
- El-Hendawy, S.E.; Al-Suhaibani, N.; Elsayed, S.; Hassan, W.M.; Dewir, Y.H.; Refay, Y.; Abdella, K.A. Potential of the existing and novel spectral reflectance indices for estimating the leaf water status and grain yield of spring wheat exposed to different irrigation rates. Agric. Water Manag. 2019, 217, 356–373. [Google Scholar] [CrossRef]
- Filella, I.; Peñuelas, J. The red edge position and shape as indicators of plant chlorophyll content, biomass and hydric status. Int. J. Remote Sens. 1994, 15, 1459–1470. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef]
- Xue, L.; Yang, L. Deriving leaf chlorophyll content of green-leafy vegetables from hyperspectral reflectance. ISPRS J. Photogramm. Remote Sens. 2009, 64, 97–106. [Google Scholar] [CrossRef]
- El-Hendawy, S.E.; Alotaibi, M.; Al-Suhaibani, N.; Al-Gaadi, K.; Hassan, W.; Dewir, Y.H.; Emam, M.A.E.-G.; Elsayed, S.; Schmidhalter, U. Comparative performance of spectral reflectance indices and multivariate modeling for assessing agronomic parameters in advanced spring wheat lines under two contrasting irrigation regimes. Front. Plant Sci. 2019, 10, 1537. [Google Scholar] [CrossRef]
- Li, W.; Sun, Z.; Lu, S.; Omasa, K. Estimation of the leaf chlorophyll content using multi-angular spectral reflectance factor. Plant Cell Environ. 2019, 42, 3152–3165. [Google Scholar] [CrossRef]
- Carter, G.A. Ratios of leaf reflectance in narrow wavebands as indicators of plant stress. Int. J. Remote Sens. 1994, 15, 697–703. [Google Scholar] [CrossRef]
- Peñuelas, J.; Baret, F.; Filella, I. Semiempirical indexes to assess carotenoids chlorophyll-a ratio from leaf spectral reflectance. Photosynthetica 1995, 31, 221–230. [Google Scholar]
- Lin, C.; Popescu, S.C.; Huang, S.C.; Chang, P.T.; Wen, H.L. A novel reflectance-based model for evaluating chlorophyll concentrations of fresh and water-stressed leaves. Biogeosciences 2015, 12, 49–66. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.; Lu, S.; Sun, Z. Estimation of leaf chlorophyll content with polarization measurements: Degree of linear polarization. J. Quant. Spectrosc. Radiat. Transf. 2020, 242, 106787. [Google Scholar] [CrossRef]
- Gitelson, A.; Merzlyak, M.N. Signature analysis of leaf reflectance spectra: Algorithm development for remote sensing of chlorophyll. J. Plant Physiol. 1996, 148, 494–500. [Google Scholar] [CrossRef]
- Çullu, M.A.; AydemIR, S.; Qadir, M.; Almaca, A.; Öztürkmen, A.R.; Biligç, A.; Agca, N. Implication of groundwater fluctuation on the seasonal salt dynamic in the Harran Plain, south-eastern Turkey. Irrig. Drain. 2009, 59, 465–476. [Google Scholar] [CrossRef]
- Bazihizina, N.; Barrett-Lennard, E.G.; Colmer, T.D. Plant growth and physiology under heterogeneous salinity. Plant Soil 2012, 354, 1–19. [Google Scholar] [CrossRef]
- Hackl, H.; Mistele, B.; Hu, Y.; Schmidhalter, U. Spectral assessments of wheat plants grown in pots and containers under saline conditions. Funct. Plant Biol. 2013, 40, 409–424. [Google Scholar] [CrossRef] [PubMed]
- El-Hendawy, S.E.; Hassan, W.M.; Refay, Y.; Schmidhalter, U. On the use of spectral reflectance indices to assess agro-morphological traits of wheat plants grown under simulated saline field conditions. J. Agron. Crop. Sci. 2017, 203, 406–428. [Google Scholar] [CrossRef]
- El-Hendawy, S.E.; Al-Suhaibani, N.; Hassan, W.; Dewir, Y.H.; El-Sayed, S.; Al-Ashkar, I.; Abdella, K.A.; Schmidhalter, U. Evaluation of wavelengths and spectral reflectance indices for high throughput assessment of growth, water relations and ion contents of wheat irrigated with saline water. Agric. Water Manag. 2019, 212, 358–377. [Google Scholar] [CrossRef]
- El-Hendawy, S.E.; Hu, Y.C.; Yakout, G.M.; Awad, A.M.; Hafiz, S.E.; Schmidhalter, U. Evaluating salt tolerance of wheat genotypes using multiple parameters. Euro. J. Agron. 2005, 22, 243–253. [Google Scholar] [CrossRef]
- El-Hendawy, S.E.; Ruan, Y.; Hu, Y.; Schmidhalter, U. A comparison of screening criteria for salt tolerance in wheat under field and environment controlled conditions. J. Agron. Crop Sci. 2009, 49, 1–9. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 603, 591–592. [Google Scholar]
- Zhang, L.; Ma, H.; Chen, T.; Pen, J.; Yu, S.; Zhao, X. Morphological and physiological responses of cotton (Gossypium hirsutum L.) plants to salinity. PLoS ONE 2014, 9, e112807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rud, R.; Shoshany, M.; Alchanatis, V. Spectral indicators for salinity effects in crops: A comparison of a new green indigo ratio with existing indices. Remote Sens. Lett. 2011, 2, 289–298. [Google Scholar] [CrossRef]
- Lara, M.A.; Diezma, B.; Lleo, L.; Roger, J.M.; Garrido, Y.; Gil, M.I.; Ruiz-Altisent, M. Hyperspectral imaging to evaluate the effect of irrigation water salinity in lettuce. Appl. Sci. 2016, 6, 412. [Google Scholar] [CrossRef]
- Yue, J.; Feng, H.; Tian, Q.; Zhou, C.A. Robust spectral angle index for remotely assessing soybean canopy chlorophyll content in different growing stages. Plant Methods 2020, 16, 104. [Google Scholar] [CrossRef]
- Lu, S.; Lu, X.; Zhao, W.; Liu, Y.; Wang, Z.; Omasa, K. Comparing vegetation indices for remote chlorophyll measurement of white poplar and Chinese elm leaves with different adaxial and abaxial surfaces. J. Exp. Bot. 2015, 66, 5625–5637. [Google Scholar] [CrossRef] [Green Version]
- Babar, M.A.; Reynolds, M.P.; van Ginkel, M.; Klatt, A.R.; Raun, W.R.; Stone, M.L. Spectral Reflectance to Estimate Genetic Variation for In-Season Biomass, Leaf Chlorophyll, and Canopy Temperature in Wheat. Crop Sci. 2006, 46, 1046–1057. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Lauchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [Green Version]
- Hamzeh, S.; Naseri, A.A.; AlaviPanah, S.K.; Mojaradi, B.; Bartholomeus, H.M.; Clevers, J.G.P.W.; Behzad, M. Estimating salinity stress in sugarcane fields with space borne hyperspectral vegetation indices. Int. J. Appl. Earth Obs. Geoinf. 2013, 21, 282–290. [Google Scholar] [CrossRef]
- Hernàndez, E.I.; Melendez-Pastor, N.; Navarro-Pedreno, J.; Gómez, I. Spectral indices for the detection of salinity effects in melon plants. Sci. Agric. 2014, 71, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.D.; Lin, K.H.; Hsu, M.H.; Huang, M.Y.; Yang, Z.W.; Chao, P.Y.; Yang, C.M. Eliminating interference by anthocyanin in chlorophyll estimation of sweet potato (Ipomoea batatas L.) leaves. Bot. Stud. 2014, 55, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, B.; Carver, B.F.; Stone, M.L.; Babar, M.A.; Raun, W.R.; Klatt, A.R. Potential use of spectral reflectance indices as a selection tool for grain yield in winter wheat under Great Plains conditions. Crop Sci. 2007, 47, 1426–1440. [Google Scholar] [CrossRef] [Green Version]
- Dobrota, C.; Lazar, L.; Baciu, C. Assessment of physiological state of Betula pendula and Carpinus betulus through leaf reflectance measurements. Flora 2015, 216, 26–34. [Google Scholar] [CrossRef]
- Zhang, T.T.; Zeng, S.L.; Gao, Y.; Ouyang, Z.T.; Li, B.; Fang, C.M.; Zhao, B. Using hyperspectral vegetation indices as a proxy to monitor soil salinity. Ecol. Indic. 2011, 11, 1552–1562. [Google Scholar] [CrossRef]
- Hu, Y.; Hackl, H.; Schmidhalter, U. Comparative performance of spectral and thermographic properties of plants and physiological traits for phenotyping salinity tolerance of wheat cultivars under simulated field conditions. Funct. Plant. Biol. 2016, 44, 134–142. [Google Scholar] [CrossRef]
- Fu, Y.; Yang, G.; Wang, J.; Song, X.; Feng, H. Winter wheat biomass estimation based on spectral indices, band depth analysis and partial least squares regression using hyperspectral measurements. Comput. Electron. Agric. 2014, 100, 51–59. [Google Scholar] [CrossRef]
- Yue, J.; Yang, G.; Li, C.; Li, Z.; Wang, Y.; Feng, H.; Xu, B. Estimation of Winter Wheat Above-Ground Biomass Using Unmanned Aerial Vehicle-Based Snapshot Hyperspectral Sensor and Crop Height Improved Models. Remote Sens. 2017, 9, 708. [Google Scholar] [CrossRef] [Green Version]
- Main, R.; Cho, M.A.; Mathieu, R.; O’Kennedy, R.M.; Ramoelo, A. An investigation into robust spectral indices for leaf chlorophyll estimation. ISPRS J. Photogramm. Remote Sens. 2011, 66, 751–761. [Google Scholar] [CrossRef]
- Christenson, B.S.; Schapaugh, W.T.; Nan, J.R.; Price, K.P.; Prasad, V.; Fritz, A.K. Predicting soybean relative maturity and seed yield using canopy reflectance. Crop Sci. 2016, 56, 625–643. [Google Scholar] [CrossRef] [Green Version]
- Schlemmer, M.; Gitelson, A.; Schepers, J.; Ferguson, R.; Peng, Y. Remote estimation of nitrogen and chlorophyll contents in maize at leaf and canopy levels. Int. J. Appl. Earth Obs. 2013, 25, 47–54. [Google Scholar]
- Gitelson, A.A.; Keydan, G.P.; Merzlyak, M.N. Three-band model for noninvasive estimation of chlorophyll, carotenoids and anthocyanin contents in higher plant leaves. Geophys. Res. Lett. 2006, 33, 33. [Google Scholar] [CrossRef] [Green Version]
- Mutanga, O.; Skidmore, A.K. Narrow band vegetation indices overcome the saturation problem in biomass estimation. Int. J. Remote Sens. 2004, 25, 3999–4014. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | SRIs | Formula |
|---|---|---|
| Constructed simple ratio type (CSR) | ||
| 1 | Simple ratio (400 and 478 nm) | R400/R478 |
| 2 | Simple ratio (810 and 550 nm) | R810/R550 |
| 3 | Simple ratio (970 and 564 nm) | R970/R564 |
| 4 | Simple ratio (554 and 572 nm) | R554/R572 |
| 5 | Simple ratio (1100 and 700 nm) | R1100/R700 |
| 6 | Simple ratio (740 and 710 nm) | R740/R710 |
| 7 | Simple ratio (754 and 568 nm) | R754/R568 |
| 8 | Simple ratio (762 and 722 nm) | R762/R722 |
| 9 | Simple ratio (1250 and 560 nm) | R1250/R560 |
| 10 | Simple ratio (1650 and 482 nm) | R1650/R482 |
| 11 | Simple ratio (2250 and 2296 nm) | R2250/R2296 |
| Published simple ratio type (PSR) | ||
| 12 | Simple ratio pigment index-1 (SRPI-1) | R430/R680 |
| 13 | Simple ratio pigment index-2 (SRPI-1) | R750/R556 |
| 14 | Blue/Green pigment Index-1 (BGI-1) | R450/R550 |
| 15 | Blue/Red pigment Index-1 (BRI-1) | R400/R690 |
| 16 | Red/green pigment Index-1 (RGI-1) | R690/R550 |
| 17 | Red/blue pigment Index (RBI) | R695/R445 |
| 18 | Gitelson and merzlyak index 1 (GM-1) | R750/R550 |
| 19 | Gitelson and merzlyak index-2 (GM-2) | R750/R700 |
| 20 | Ratio analysis of reflectance spectra-a (PARS-a) | R750/R710 |
| 21 | Ratio analysis of reflectance spectra-a developed (PARS-a-D) | R780/R720 |
| 22 | Ratio analysis of reflectance spectra-c (PARS-c) | R760/R500 |
| 23 | Ratio analysis of reflectance spectra-c developed (PARS-c-D) | R760/R515 |
| 24 | Pigment Specific Simple ratio-a (PSSR-a) | R800/R675 |
| 25 | Pigment-specific simple ratio-b (PSSR-b) | R800/R650 |
| 26 | Pigment-specific simple ratio-c (PSSR-c) | R800/R470 |
| Published modified simple ratio type (PMSR) | ||
| 27 | Red edge chlorophyll index (ClRed-edge) | (R750/R710) − 1 |
| 28 | Red edge chlorophyll index –developed (Cl-DRed-edge) | (R760/R710) − 1 |
| 29 | Green chlorophyll index (ClGreen) | (R800/R550) − 1 |
| 30 | Ratio analysis of reflectance spectra-a (PARS-b) | R675/(R650*R700) |
| 31 | Chlorophyll reflectance index-a (Chl-a) | R776 (1/R673 − 1) |
| 32 | Chlorophyll reflectance index-b (Chl-b) | R776 (1/R625 − 1/R673) |
| 33 | Carotenoid reflectance index (CRI) | (1/R507 − 1/R603 − 0.65*(1/R530))*R776 |
| 34 | Anthocyanin (Gitelson) (AntGitelson) | R780 (1/R550 − 1/R700) |
| 35 | Plant senescence reflectance index (PSRI) | (R680 − R500)/R750 |
| 36 | Red-edge vegetation stress index (RVSI) | 0.5 (R722 + R763) − R733 |
| Published normalized difference type (PND) | ||
| 37 | Normalized phaeophytinization index (NPQ) | (R415 − R435)/(R415+R435) |
| 38 | Normalized phaeophytinization index developed (NPQ-D) | (R482 − R350)/(R482+R350) |
| 39 | Photochemical reflectance index (PRI) | (R531 − R570)/(R531+R570) |
| 40 | Photochemical reflectance index developed (PRI-D) | (R531 − R580)/(R531+R580) |
| 41 | Modified simple ratio of reflectance-1 (MSR-1) | (R750 − R445)/(R705-R445) |
| 42 | Modified simple ratio of reflectance-2 (MSR-2) | (R780 − R710)/(R780-R680) |
| 43 | Normalized difference vegetation index (NDVI) | (R750 − R680)/(R750+R680) |
| 44 | Normalized difference vegetation index developed (NDVI-D) | (R780 − R715)/(R780+R715) |
| 45 | Structure insensitive pigment index (SIPI) | (R800 − R445)/(R800-R680) |
| 46 | Pigment specific normalized difference-a (PSND-a) | (R800 − R680)/(R800+R680) |
| 47 | Pigment specific normalized difference-b (PSND-b) | (R800 − R635)/(R800+R635) |
| 48 | Pigment specific normalized difference-c (PSND-c) | (R800 − R460)/(R800+R460) |
| 49 | Pigment specific normalized difference-c developed (PSND-c-D) | (R800 − R482)/(R800+R482) |
| Published integrated form type (PIF) | ||
| 50 | Chlorophyll absorption ratio index (CARI) | [(R700 − R670) − 0.2 × (R700 − R550)] |
| 51 | Modified chlorophyll absorption ratio index (MCARI) | [(R700 − R670) − 0.2 × (R700 − R550)] × (R700/R670) |
| 52 | Transformed chlorophyll absorption reflectance index (TCARI) | 3 × ((R700 − R670) − 0.2 × (R700 − R550)] × (R700/R670) |
| 53 | Optimized soil-adjusted vegetation index (OSAVI) | 1.16(R800 − R670)/(R800 + R670 + 0.16) |
| 54 | Triangular vegetation index (TVI) | 1.2 × (R700 − R550) − 1.5 × (R670 − R550) × (R700/R670)1/2] |
| 55 | Modified triangular vegetation index (MTVI) | 1.2 [1.2(R800 − R550) − 2.5(R670 − R550)] |
| 56 | Enhanced vegetation index (EVI) | 2.5 (R782 − R675)/(R782 + 6 × R675 − 7.5 × R445 + 1)) |
| 57 | Red edge inflection point (REIP) | REIP = 700 + 40 × {[(R670 + R780)/2 − R700]/(R740 − R700)} |
| 58 | Salinity and water stress index-1 (SWSI-1) | (R803 − R681)/root(R905 − R972) |
| 59 | Salinity and water stress index-2 (SWSI-2) | (R803 − R681)/root(R1326 − R1507) |
| 60 | Salinity and water stress index-3 (SWSI-3) | (R803 − R681)/root(R972 − R1174) |
| F-Values | ||||||
|---|---|---|---|---|---|---|
| Source of Variance | df | SDW (g plant−1) | Chla (mg g−1 Fresh Weight) | Chlb (mg g−1 Fresh Weight) | Chlt (mg g−1 Fresh Weight) | |
| Year (Y) | 1 | 119.8 ** | 32.8 * | 905.3 ** | 88.7 * | |
| Salinity (S) | 2 | 574.9 *** | 315.2 *** | 73.3 *** | 323.1 *** | |
| S*Y | 2 | 0.38 ns | 2.65 ns | 2.16 ns | 0.26 ns | |
| Genotypes (G) | 1 | 119.6 *** | 30.3 *** | 54.1 *** | 37.9 *** | |
| G*Y | 1 | 1.28 ns | 0.029 ns | 0.91 ns | 0.15 ns | |
| G*S | 2 | 19.8 *** | 12.3 ** | 9.59 ** | 12.1 ** | |
| G*S*Y | 2 | 1.04 ns | 0.64 ns | 0.29 ns | 0.57 ns | |
| Mean values of the main factor ± standard deviation | ||||||
| Salinity | Control | 7.70a ± 0.50 | 2.65a ± 0.22 | 0.844a ± 0.11 | 3.49a ± 0.31 | |
| 6 dS m−1 | 5.34b ± 0.82 | 2.34b ± 0.25 | 0.692b ± 0.14 | 3.03b ± 0.38 | ||
| 12 dS m−1 | 3.67c ± 0.91 | 1.93c ± 0.37 | 0.580c ± 0.13 | 2.51c ± 0.50 | ||
| Genotypes | Sakha 93 | 6.04a ± 1.55 | 2.48a ± 0.21 | 0.784a ± 0.12 | 3.26a ± 0.34 | |
| Sakha 61 | 5.10b ± 2.02 | 2.14b ± 0.47 | 0.626b ± 0.17 | 2.76b ± 0.64 | ||
| SRIs | Y | S | S × Y | G | G × Y | G × S | G × S × Y | SRIs | Y | S | S × Y | G | G × Y | G × S | G × S × Y | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Constructed simple ratio type (CSR) | 1 | ns | *** | ns | *** | ns | *** | ns | Published modified simple ratio type (PMSR) | 31 | ns | *** | ns | *** | ns | *** | ns |
| 2 | ns | *** | ns | *** | ns | * | ns | 32 | ns | * | ns | *** | * | ** | ns | ||
| 3 | ns | *** | ns | *** | ns | * | ns | 33 | ns | *** | ns | ** | ns | ns | ns | ||
| 4 | ns | *** | ns | *** | ns | *** | ns | 34 | ns | ns | ns | * | ns | ns | ns | ||
| 5 | ns | *** | ns | *** | ns | ** | ns | 35 | ns | *** | ns | *** | * | *** | ns | ||
| 6 | ** | *** | ns | *** | ns | *** | ns | 36 | ns | *** | ns | *** | ns | ns | ns | ||
| 7 | ns | *** | ns | *** | ns | ** | ns | Published normalized difference type (PND) | 37 | ns | *** | * | *** | ns | ** | * | |
| 8 | ns | *** | ns | *** | ns | ** | ns | 38 | ns | *** | ns | *** | ns | *** | * | ||
| 9 | * | *** | ns | ** | ns | ns | ns | 39 | ns | *** | ns | *** | ns | *** | ns | ||
| 10 | ns | *** | *** | *** | ns | ns | ns | 40 | ns | *** | ns | *** | ns | *** | ns | ||
| 11 | ns | *** | ns | *** | ns | ** | ns | 41 | ns | *** | ns | *** | ns | ** | ns | ||
| Published simple ratio type (PSR) | 12 | ns | ** | ns | *** | ns | ** | ns | 42 | ns | *** | ns | ** | ns | ns | ns | |
| 13 | ns | *** | ns | *** | ns | ** | ns | 43 | ns | *** | ns | *** | * | *** | ns | ||
| 14 | ns | ns | ns | ns | ns | ns | ns | 44 | * | *** | ns | *** | ns | *** | ns | ||
| 15 | ns | *** | ns | *** | ns | ** | ns | 45 | ns | ** | ns | *** | ns | ** | ns | ||
| 16 | ns | *** | ns | *** | * | *** | ns | 46 | ns | *** | ns | *** | * | *** | ns | ||
| 17 | ns | *** | ns | *** | ns | * | ns | 47 | ns | *** | ns | *** | * | *** | ns | ||
| 18 | ns | *** | ns | *** | ns | * | ns | 48 | ns | *** | ns | *** | ns | ** | ns | ||
| 19 | * | *** | ns | *** | ns | *** | ns | 49 | ns | *** | ns | *** | ns | ** | ns | ||
| 20 | ** | *** | ns | *** | ns | *** | ns | Published integrated form type (PIF) | 50 | ns | * | ns | ns | ns | ns | ns | |
| 21 | ns | *** | ns | *** | ns | ** | ns | 51 | ns | ns | ns | ** | ns | * | ns | ||
| 22 | ns | *** | * | *** | ns | ** | ns | 52 | ns | ns | ns | ** | ns | * | ns | ||
| 23 | ns | *** | ns | *** | ns | ** | ns | 53 | ns | *** | ns | *** | * | *** | ns | ||
| 24 | ns | *** | ns | *** | ns | ** | ns | 54 | ns | ns | ns | * | ns | * | ns | ||
| 25 | ns | *** | * | *** | ns | *** | ns | 55 | ns | ** | ns | *** | * | *** | ns | ||
| 26 | ns | *** | ** | *** | ns | * | ns | 56 | ns | *** | ns | *** | * | *** | * | ||
| 27 | ** | *** | ns | *** | ns | *** | ns | 57 | ns | *** | ns | ** | ns | ns | ns | ||
| 28 | * | *** | ns | *** | ns | *** | ns | 58 | ns | ns | ns | ns | ns | ns | ns | ||
| 29 | ns | *** | ns | *** | ns | * | ns | 59 | ns | *** | ns | *** | ns | ** | ns | ||
| 30 | * | *** | ns | *** | ns | *** | ns | 60 | ns | ns | ns | ** | ns | ns | ns | ||
| SRIs | Genotypes | Salinity Levels | SRIs | Genotypes | Salinity Levels | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S93 | S61 | C | 6 dS m−1 | 12 dS m−1 | S93 | S61 | C | 6 dS m−1 | 12 dS m−1 | ||||||
| Constructed simple ratio type (CSR) | 1 | 0.79a | 0.64b | 0.84a | 0.67b | 0.62b | Published modified simple ratio type (PMSR) | 31 | 10.56a | 6.18b | 14.32a | 5.95b | 4.84b | ||
| 2 | 5.62a | 4.25b | 7.01a | 4.25b | 3.55c | 32 | −1.82b | −0.77a | −1.90b | −0.95a | −1.03a | ||||
| 3 | 5.81a | 4.33b | 7.39a | 4.25b | 3.57c | 33 | −2.04b | −1.24a | −2.90b | −1.17a | −0.86a | ||||
| 4 | 1.19a | 1.08b | 1.25a | 1.09b | 1.07c | 34 | 0.40b | 0.70a | 0.31a | 0.69a | 0.65a | ||||
| 5 | 5.11a | 3.49b | 6.49a | 3.51b | 2.90c | 35 | 0.009b | 0.11a | 0.004b | 0.084a | 0.091a | ||||
| 6 | 2.50a | 1.94b | 2.92a | 1.99b | 1.75c | 36 | 0.032a | 0.021b | 0.039a | 0.024b | 0.016c | ||||
| 7 | 5.95a | 4.28b | 7.64a | 4.23b | 3.48c | Published normalized difference type (PND) | 37 | −0.057a | −0.088b | −0.046a | −0.081b | −0.090b | |||
| 8 | 1.92a | 1.60b | 2.14a | 1.66b | 1.49c | 38 | −0.014b | 0.14a | −0.091b | 0.11a | 0.17a | ||||
| 9 | 4.87a | 3.76b | 6.09a | 3.68b | 3.17c | 39 | 0.023a | −0.028b | 0.045a | −0.024b | −0.029b | ||||
| 10 | 6.23a | 4.98b | 7.84a | 4.69b | 4.28c | 40 | 0.083a | −0.001b | 0.119a | 0.007b | −0.002b | ||||
| 11 | 1.17a | 1.13b | 1.18a | 1.14b | 1.12b | 41 | 4.87a | 3.28b | 6.08a | 3.40b | 2.75c | ||||
| Published simple ratio type (PSR) | 12 | 0.83a | 0.56b | 0.89a | 0.61b | 0.60b | 42 | 0.74a | 0.68b | 0.79a | 0.70b | 0.65c | |||
| 13 | 5.13a | 3.85b | 6.38a | 3.84b | 3.24c | 43 | 0.80a | 0.57b | 0.85a | 0.62b | 0.59b | ||||
| 14 | 0.48a | 0.47a | 0.47a | 0.47a | 0.47a | 44 | 0.43a | 0.30b | 0.49a | 0.33b | 0.27c | ||||
| 15 | 0.57a | 0.38b | 0.64a | 0.41b | 0.38b | 45 | 1.02b | 1.24a | 1.01b | 1.17a | 1.22a | ||||
| 16 | 1.00a | 0.65b | 0.94a | 0.94a | 0.60b | 46 | 0.81a | 0.60b | 0.86a | 0.65b | 0.61b | ||||
| 17 | 2.47a | 1.79b | 2.40a | 2.36a | 1.64b | 47 | 0.79a | 0.59b | 0.86a | 0.63b | 0.59b | ||||
| 18 | 5.10a | 3.87b | 6.32a | 3.87b | 3.27c | 48 | 0.84a | 0.76b | 0.87a | 0.78b | 0.74c | ||||
| 19 | 4.73a | 3.22b | 6.03a | 3.22b | 2.66c | 49 | 0.84a | 0.74b | 0.87a | 0.76b | 0.73c | ||||
| 20 | 2.85a | 2.14b | 3.38a | 2.21b | 1.90c | Published integrated form type (PIF) | 50 | 0.083a | 0.074a | 0.094a | 0.080ab | 0.062b | |||
| 21 | 2.11a | 1.73b | 2.37a | 1.79b | 1.59c | 51 | 0.186a | 0.132b | 0.145a | 0.149a | 0.182a | ||||
| 22 | 10.91a | 7.67b | 14.14a | 7.48b | 6.25b | 52 | 0.558a | 0.395b | 0.436a | 0.448a | 0.547a | ||||
| 23 | 8.56a | 6.16b | 11.1a | 6.02b | 4.96c | 53 | 0.803a | 0.599b | 0.845a | 0.646b | 0.613b | ||||
| 24 | 11.23a | 6.82b | 14.81a | 6.67b | 5.60b | 54 | 0.186a | 0.147b | 0.142b | 0.163ab | 0.195a | ||||
| 25 | 11.20a | 7.02b | 15.55a | 6.49b | 5.28b | 55 | 1.103a | 0.778b | 1.079a | 0.880b | 0.862b | ||||
| 26 | 12.08a | 8.80b | 15.38a | 8.66b | 7.29c | 56 | 0.999a | 0.654b | 1.023a | 0.736b | 0.720b | ||||
| 27 | 1.85a | 1.14b | 2.38a | 1.21b | 0.99c | 57 | 722.31a | 720.01b | 723.98a | 720.61b | 718.88c | ||||
| 28 | 2.01a | 1.25b | 2.60a | 1.32b | 0.97c | 58 | 3.06a | 2.84a | 3.13a | 2.92a | 2.80a | ||||
| 29 | 4.60a | 3.23b | 5.99a | 3.23b | 2.53c | 59 | 1.17a | 0.99b | 1.17a | 1.05b | 1.03b | ||||
| 30 | 6.81a | 5.09b | 9.53a | 4.66b | 3.65c | 60 | 2.11a | 1.86b | 2.06a | 1.98b | 1.91b | ||||
| SRIs Groups | Measured Variables | Influential SRI | Best Fitted Equation | Model R2 | Model RMSE |
|---|---|---|---|---|---|
| Constructed simple ratio type (CSR) | SDW | SRI(1250,560) | SDW = 0.909 +1.08 (SRI(1250,560)) | 0.80 | 0.836 |
| Chla | SRI(740,710) | Chla = 1.139 + 0.526 (SRI(740,710)) | 0.66 | 0.242 | |
| Chlb | SRI(700,1100) | Chlb = 0.974 − 0.928 (SRI(700,1100)) | 0.62 | 0.105 | |
| Chlt | SRI(740,710) | Chlt = 1.358 + 0.732 SRI(740,710) | 0.67 | 0.331 | |
| Published simple ration type (PSR) | SDW | GMI | SDW = 1.006 + 1.017 (GMI) | 0.79 | 0.846 |
| Chla | PARS-a | Chla = 1.268 + 0.416 (PARS-a) | 0.65 | 0.244 | |
| Chlb | PARS-D-a | Chlb = 0.114 + 0.308 (PARSD-a) | 0.60 | 0.107 | |
| Chlt | PARS-a | Chlt = 1.566 + 0.579 (PARS-a) | 0.66 | 0.334 | |
| Published modified simple ratio type (PMSR) | SDW | PARS-b | SDW = 2.258 + 0.557 (PARS-b) | 0.81 | 0.806 |
| Chla | CIRed-edge | Chla = 1.684 + 0.416 (CIRed-edge) | 0.65 | 0.244 | |
| Chlb | CIRed-edge, PARS-b | Chlb = 0.50 + 0.43 (CIRed-edge) − 0.007 (PARS-b) | 0.68 | 0.098 | |
| Chlt | CIRed-edge, PARS-b | Chlt = 2.23 + 1.22 (CIRed-edge) − 0.18 (PARS-b) | 0.70 | 0.318 | |
| Published normalized difference type (PND) | SDW | NDVI-D | SDW = 0.950+12.718 (NDV3-D) | 0.75 | 0.928 |
| Chla | NDVI-D | Chla = 1.326 + 2.701 (NDVI-D) | 0.68 | 0.234 | |
| Chlb | NDVI-D | Chlb = 0.320 + 1.06 (NDVI-D) | 0.63 | 0.103 | |
| Chlt | NDVI-D | Chlt = 1.646 + 3.76 (NDVI-D) | 0.69 | 0.318 | |
| Published integrated form type (PIF) | SDW | OSAVI, REIP | SDW = −211.45 + 4.48 (OSAVI) + 0.30 (REIP) | 0.69 | 1.051 |
| Chla | TCARI, OSAVI | Chla = 1.076 − 0.602 (TCARI) + 2.165 (OSAVI) | 0.68 | 0.238 | |
| Chlb | EVI-1, REIP | Chlb = −15.074 + 0.316 (EVI-1) + 0.002 (REIP) | 0.65 | 0.102 | |
| Chlt | OSAVI, REIP | Chlt = −49.277 + 1.72 (OSAVI) +0.007 (REIP) | 0.69 | 0.325 |
| SRI Type | Measured Variables | Control | Moderate Salinity Level (6 dS m−1) | High Salinity Level (12 dS m−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Equation | R2 | RMSE | Equation | R2 | RMSE | Equation | R2 | RMSE | ||
| Constructed simple ratio type (CSR) | SDW | y = 0.7885x + 1.411 | 0.23 | 0.732 | y = 1.1424x − 1.208 | 0.65 | 0.809 | y = 0.476x + 2.587 | 0.55 | 0.889 |
| Chla | y = −0.0549x + 2.789 | 0.04 | 0.230 | y = 1.0082x − 0.151 | 0.67 | 0.212 | y = 0.5552x + 0.995 | 0.68 | 0.250 | |
| Chlb | y = −0.0432x + 0.866 | 0.11 | 0.113 | y = 0.6707x + 0.199 | 0.58 | 0.095 | y = 0.6103x + 0.269 | 0.53 | 0.098 | |
| Chlt | y = −0.0564x + 3.649 | 0.11 | 0.317 | y = 1.0317x − 0.267 | 0.66 | 0.272 | y = 0.628x + 1.144 | 0.64 | 0.357 | |
| Published simple ration type (PSR) | SDW | y = 0.5635x + 3.0961 | 0.16 | 0.708 | y = 1.1356x − 1.121 | 0.57 | 0.880 | y = 0.6262x + 2.034 | 0.59 | 0.868 |
| Chla | y = −0.1091x + 2.963 | 0.07 | 0.248 | y = 1.0194x − 0.199 | 0.82 | 0.190 | y = 0.398x + 1.290 | 0.69 | 0.268 | |
| Chlb | y = −0.0754x + 0.911 | 0.04 | 0.122 | y = 0.5676x + 0.273 | 0.55 | 0.097 | y = 0.3971x + 0.374 | 0.58 | 0.091 | |
| Chlt | y = −0.1208x + 3.946 | 0.09 | 0.350 | y = 0.8905x + 0.145 | 0.77 | 0.261 | y = 0.4049x + 1.649 | 0.67 | 0.353 | |
| Published modified simple ratio type (PMSR) | SDW | y = 0.3545x + 4.833 | 0.08 | 0.655 | y = 1.0647x − 0.831 | 0.59 | 0.852 | y = 0.5072x + 2.430 | 0.63 | 0.829 |
| Chla | y = −0.1091x + 2.963 | 0.07 | 0.248 | y = 1.0194x − 0.199 | 0.82 | 0.190 | y = 0.398x + 1.290 | 0.69 | 0.268 | |
| Chlb | y = 0.1155x + 0.727 | 0.06 | 0.109 | y = 0.8061x + 0.118 | 0.68 | 0.083 | y = 0.6485x + 0.239 | 0.61 | 0.087 | |
| Chlt | y = −0.0231x + 3.543 | 0.00 | 0.338 | y = 1.0921x − 0.424 | 0.80 | 0.248 | y = 0.5797x + 1.230 | 0.71 | 0.321 | |
| Published normalized difference type (PND) | SDW | y = 0.1511x + 6.037 | 0.06 | 0.705 | y = 1.3114x − 1.892 | 0.55 | 0.992 | y = 0.9458x + 0.929 | 0.62 | 0.979 |
| Chla | y = −0.0796x + 2.864 | 0.07 | 0.235 | y = 1.109x − 0.388 | 0.88 | 0.194 | y = 0.5149x + 1.064 | 0.69 | 0.249 | |
| Chlb | y = −0.0529x + 0.886 | 0.05 | 0.115 | y = 0.6297x + 0.231 | 0.66 | 0.096 | y = 0.5149x + 0.309 | 0.56 | 0.089 | |
| Chlt | y = −0.0811x + 3.778 | 0.07 | 0.329 | y = 0.9726x − 0.075 | 0.73 | 0.264 | y = 0.524x + 1.350 | 0.66 | 0.331 | |
| Published integrated form type (PIF) | SDW | y = 0.1053x + 6.239 | 0.03 | 0.830 | y = 1.2462x − 1.491 | 0.47 | 1.076 | y = 0.9116x + 1.153 | 0.56 | 1.091 |
| Chla | y = −0.038x + 2.743 | 0.03 | 0.222 | y = 1.1193x − 0.415 | 0.78 | 0.198 | y = 0.5564x + 0.999 | 0.64 | 1.504 | |
| Chlb | y = 0.0103x + 0.819 | 0.00 | 0.110 | y = 0.7442x + 0.150 | 0.62 | 0.091 | y = 0.6731x + 0.233 | 0.61 | 0.091 | |
| Chlt | y = −0.0648x + 3.685 | 0.06 | 0.324 | y = 1.0211x − 0.218 | 0.73 | 0.274 | y = 0.6243x + 1.131 | 0.68 | 0.331 | |
| SRI Type | Measured Variables | Salt-Tolerant Genotype Sakha 93 | Salt-Sensitive Genotype Sakha 61 | ||||
|---|---|---|---|---|---|---|---|
| Equation | R2 | RMSE | Equation | R2 | RMSE | ||
| Constructed simple ratio type (CSR) | SDW | y = 0.6537x + 2.836 | 0.84 | 1.388 | y = 0.725x + 1.271 | 0.80 | 0.907 |
| Chla | y = 0.4303x + 1.403 | 0.30 | 0.203 | y = 0.6552x + 0.745 | 0.69 | 0.257 | |
| Chlb | y = 0.2178x + 0.607 | 0.22 | 0.103 | y = 0.6688x + 0.214 | 0.64 | 0.102 | |
| Chlt | y = 0.3355x + 2.175 | 0.34 | 0.271 | y = 0.66x + 0.931 | 0.67 | 0.357 | |
| Published simple ration type (PSR) | SDW | y = 0.7019x + 0.858 | 0.73 | 1.223 | y = 0.7786x + 0.976 | 0.82 | 0.850 |
| Chla | y = 0.537x + 1.122 | 0.32 | 0.214 | y = 0.6259x + 0.824 | 0.69 | 0.259 | |
| Chlb | y = 0.328x + 0.507 | 0.18 | 0.115 | y = 0.6721x + 0.226 | 0.73 | 0.091 | |
| Chlt | y = 0.5062x + 1.564 | 0.30 | 0.310 | y = 0.6569x + 0.994 | 0.71 | 0.338 | |
| Published modified simple ratio type (PMSR) | SDW | y = 0.7391x + 1.585 | 0.79 | 0.688 | y = 0.8311x + 0.851 | 0.81 | 0.868 |
| Chla | y = 0.537x + 1.122 | 0.32 | 0.214 | y = 0.6259x + 0.824 | 0.69 | 0.259 | |
| Chlb | y = 0.374x + 0.500 | 0.32 | 0.096 | y = 0.6289x + 0.224 | 0.72 | 0.091 | |
| Chlt | y = 0.5043x + 1.645 | 0.34 | 0.276 | y = 0.6235x + 1.012 | 0.73 | 0.331 | |
| Published normalized difference type (PND) | SDW | y = 0.5115x + 3.273 | 0.73 | 0.930 | y = 0.7846x + 0.775 | 0.83 | 0.873 |
| Chla | y = 0.479x + 1.288 | 0.34 | 0.197 | y = 0.6514x + 0.747 | 0.70 | 0.259 | |
| Chlb | y = 0.2923x + 0.542 | 0.20 | 0.107 | y = 0.6961x + 0.203 | 0.70 | 0.094 | |
| Chlt | y = 0.4536x + 1.767 | 0.32 | 0.286 | y = 0.6805x + 0.898 | 0.72 | 0.332 | |
| Published integrated form type (PIF) | SDW | y = 0.4501x + 3.650 | 0.68 | 1.007 | y = 0.7153x + 1.121 | 0.77 | 1.006 |
| Chla | y = 0.453x + 1.356 | 0.37 | 0.189 | y = 0.6467x + 0.753 | 0.68 | 0.262 | |
| Chlb | y = 0.2622x + 0.579 | 0.25 | 0.101 | y = 0.6766x + 0.202 | 0.69 | 0.094 | |
| Chlt | y = 0.3802x + 2.029 | 0.32 | 0.276 | y = 0.6676x + 0.911 | 0.70 | 0.342 | |
| SRI Type | Measured Variables | First Season | Second Season | ||||
|---|---|---|---|---|---|---|---|
| Equation | R2 | RMSE | Equation | R2 | RMSE | ||
| Constructed simple ratio type (CSR) | SDW | y = 0.9072x + 0.326 | 0.82 | 0.777 | y = 0.7325x + 1.609 | 0.80 | 0.847 |
| Chla | y = 0.6009x + 0.996 | 0.69 | 0.256 | y = 0.8293x + 0.302 | 0.79 | 0.203 | |
| Chlb | y = 0.7281x + 0.250 | 0.74 | 0.102 | y = 0.7802x + 0.093 | 0.84 | 0.102 | |
| Chlt | y = 0.6186x + 1.270 | 0.68 | 0.351 | y = 0.8533x + 0.288 | 0.84 | 0.279 | |
| Published simple ration type (PSR) | SDW | y = 0.8832x + 0.437 | 0.84 | 0.754 | y = 0.7534x + 1.533 | 0.78 | 0.886 |
| Chla | y = 0.6209x + 0.932 | 0.64 | 0.262 | y = 0.7465x + 0.518 | 0.74 | 0.211 | |
| Chlb | y = 0.7786x + 0.212 | 0.70 | 0.103 | y = 0.6996x + 0.161 | 0.80 | 0.105 | |
| Chlt | y = 0.6751x + 1.090 | 0.67 | 0.348 | y = 0.3955x + 1.421 | 0.76 | 0.595 | |
| Published modified simple ratio type (PMSR) | SDW | y = 0.8981x + 0.372 | 0.85 | 0.712 | y = 0.7721x + 1.412 | 0.79 | 0.849 |
| Chla | y = 0.6209x + 0.932 | 0.64 | 0.262 | y = 0.7465x + 0.518 | 0.74 | 0.211 | |
| Chlb | y = 0.73x + 0.230 | 0.68 | 0.096 | y = 0.8121x + 0.088 | 0.82 | 0.092 | |
| Chlt | y = 0.6396x + 1.158 | 0.66 | 0.377 | y = 0.8358x + 0.395 | 0.81 | 0.269 | |
| Published normalized difference type (PND) | SDW | y = 0.7573x + 1.126 | 0.81 | 0.836 | y = 0.8031x + 1.341 | 0.75 | 0.963 |
| Chla | y = 0.6154x + 0.949 | 0.66 | 0.256 | y = 0.8245x + 0.329 | 0.80 | 0.194 | |
| Chlb | y = 0.7773x + 0.215 | 0.73 | 0.101 | y = 0.7583x + 0.114 | 0.84 | 0.100 | |
| Chlt | y = 0.671x + 1.107 | 0.70 | 0.378 | y = 0.8217x + 0.400 | 0.83 | 0.278 | |
| Published integrated form type (PIF) | SDW | y = 0.7105x + 1.3684 | 0.72 | 0.985 | y = 0.7319x + 1.745 | 0.71 | 1.027 |
| Chla | y = 0.5726x + 1.047 | 0.65 | 0.261 | y = 0.8698x + 0.218 | 0.81 | 0.190 | |
| Chlb | y = 0.7721x + 0.209 | 0.69 | 0.099 | y = 0.7603x + 0.121 | 0.82 | 0.096 | |
| Chlt | y = 0.6558x + 1.147 | 0.67 | 0.346 | y = 0.8282x + 0.383 | 0.84 | 0.271 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Hendawy, S.; Elsayed, S.; Al-Suhaibani, N.; Alotaibi, M.; Tahir, M.U.; Mubushar, M.; Attia, A.; Hassan, W.M. Use of Hyperspectral Reflectance Sensing for Assessing Growth and Chlorophyll Content of Spring Wheat Grown under Simulated Saline Field Conditions. Plants 2021, 10, 101. https://doi.org/10.3390/plants10010101
El-Hendawy S, Elsayed S, Al-Suhaibani N, Alotaibi M, Tahir MU, Mubushar M, Attia A, Hassan WM. Use of Hyperspectral Reflectance Sensing for Assessing Growth and Chlorophyll Content of Spring Wheat Grown under Simulated Saline Field Conditions. Plants. 2021; 10(1):101. https://doi.org/10.3390/plants10010101
Chicago/Turabian StyleEl-Hendawy, Salah, Salah Elsayed, Nasser Al-Suhaibani, Majed Alotaibi, Muhammad Usman Tahir, Muhammad Mubushar, Ahmed Attia, and Wael M. Hassan. 2021. "Use of Hyperspectral Reflectance Sensing for Assessing Growth and Chlorophyll Content of Spring Wheat Grown under Simulated Saline Field Conditions" Plants 10, no. 1: 101. https://doi.org/10.3390/plants10010101
APA StyleEl-Hendawy, S., Elsayed, S., Al-Suhaibani, N., Alotaibi, M., Tahir, M. U., Mubushar, M., Attia, A., & Hassan, W. M. (2021). Use of Hyperspectral Reflectance Sensing for Assessing Growth and Chlorophyll Content of Spring Wheat Grown under Simulated Saline Field Conditions. Plants, 10(1), 101. https://doi.org/10.3390/plants10010101