Strategy of Salt Tolerance and Interactive Impact of Azotobacter chroococcum and/or Alcaligenes faecalis Inoculation on Canola (Brassica napus L.) Plants Grown in Saline Soil

 ,
, 
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Microbiological Characteristics in the Canola Rhizosphere
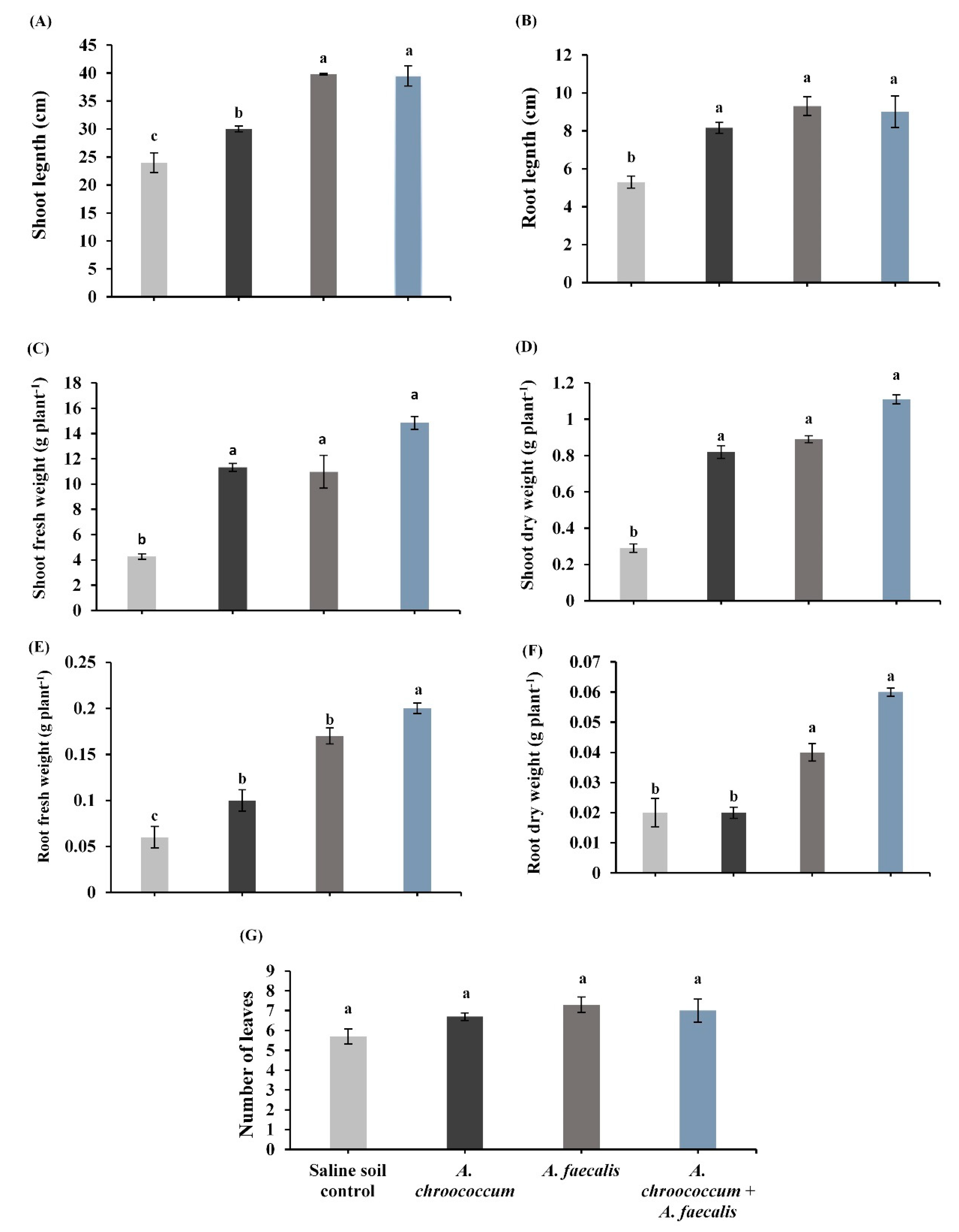
2.2. PGPR Enhance Canola Plant Growth under Salinity Stress
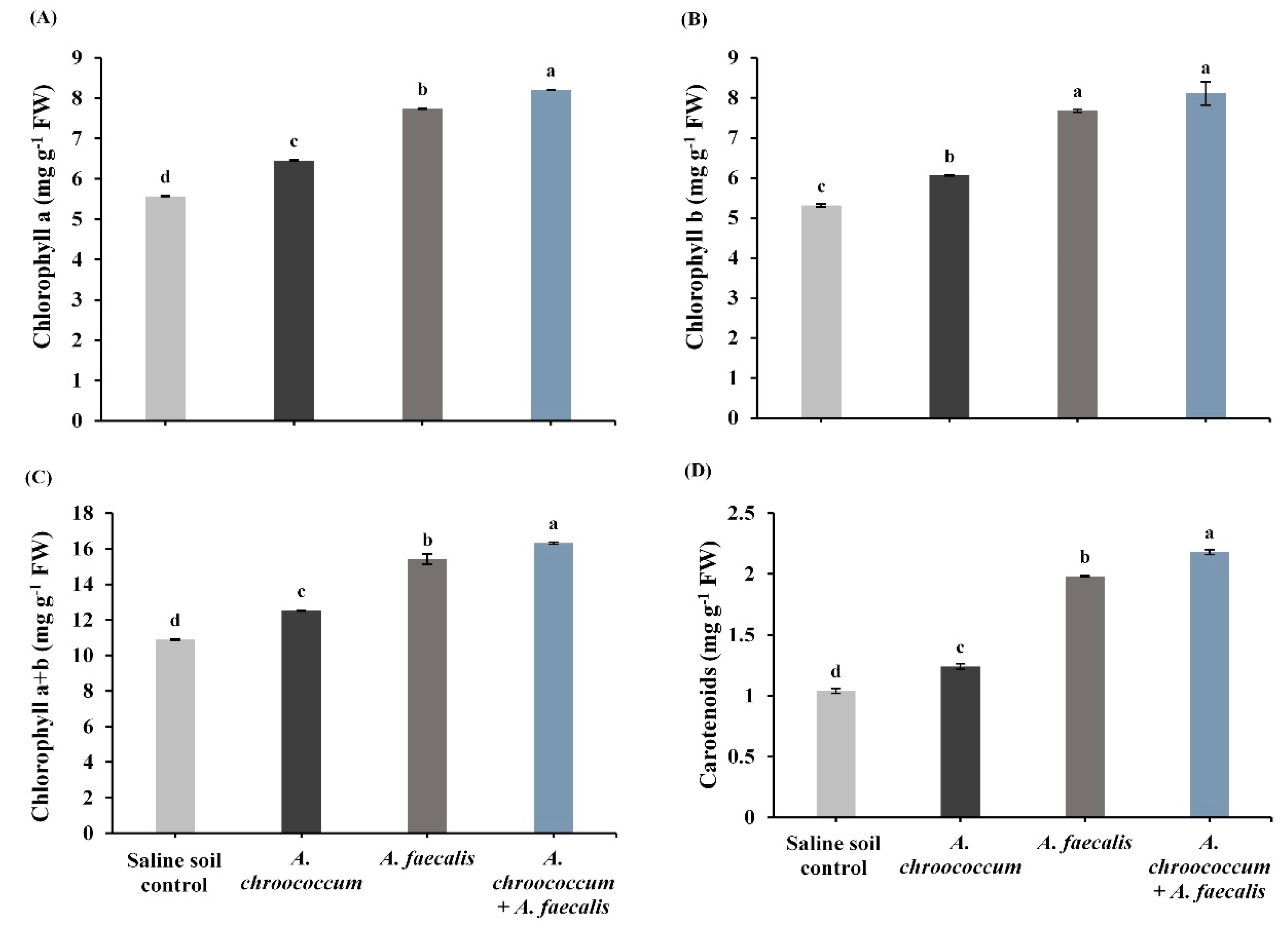
2.3. PGPR Protect Photosynthetic Pigments in Leaves of Canola Plant under Salinity Stress
2.4. PGPR Regulate Osmolytes’ Contents in Salinity-Stressed Canola Plants
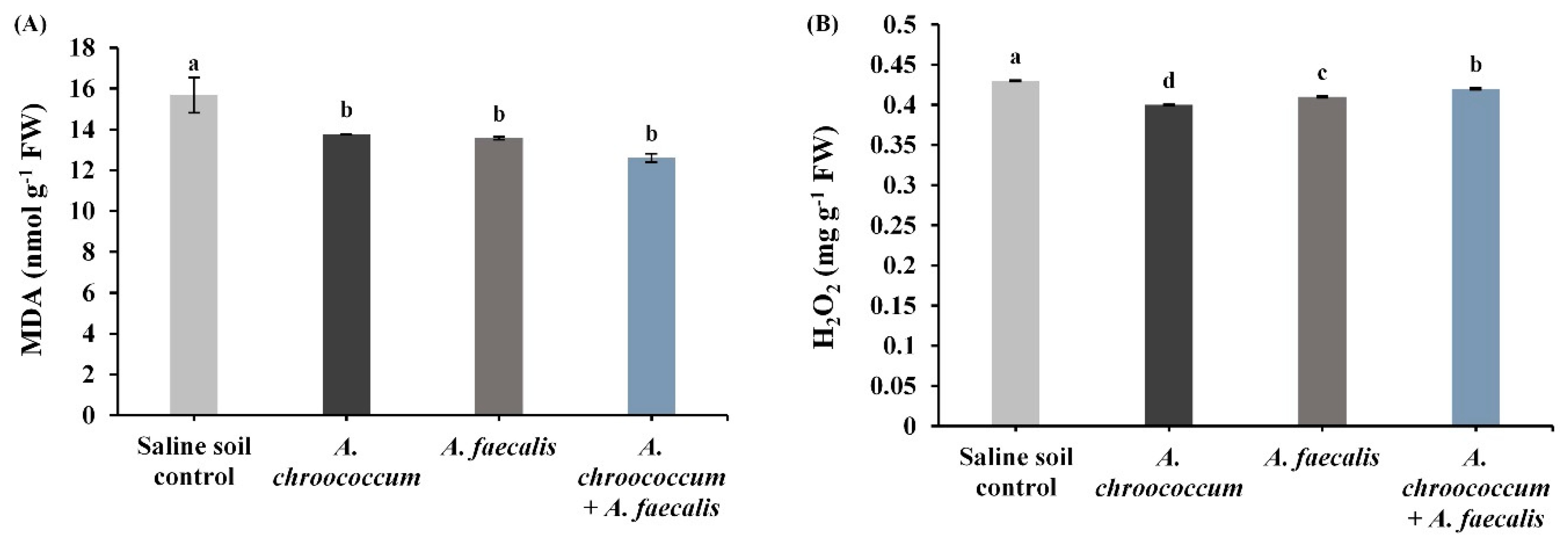
2.5. PGPR Lessen MDA and H2O2 Contents in Leaves of Salinity-Stressed Canola Plants
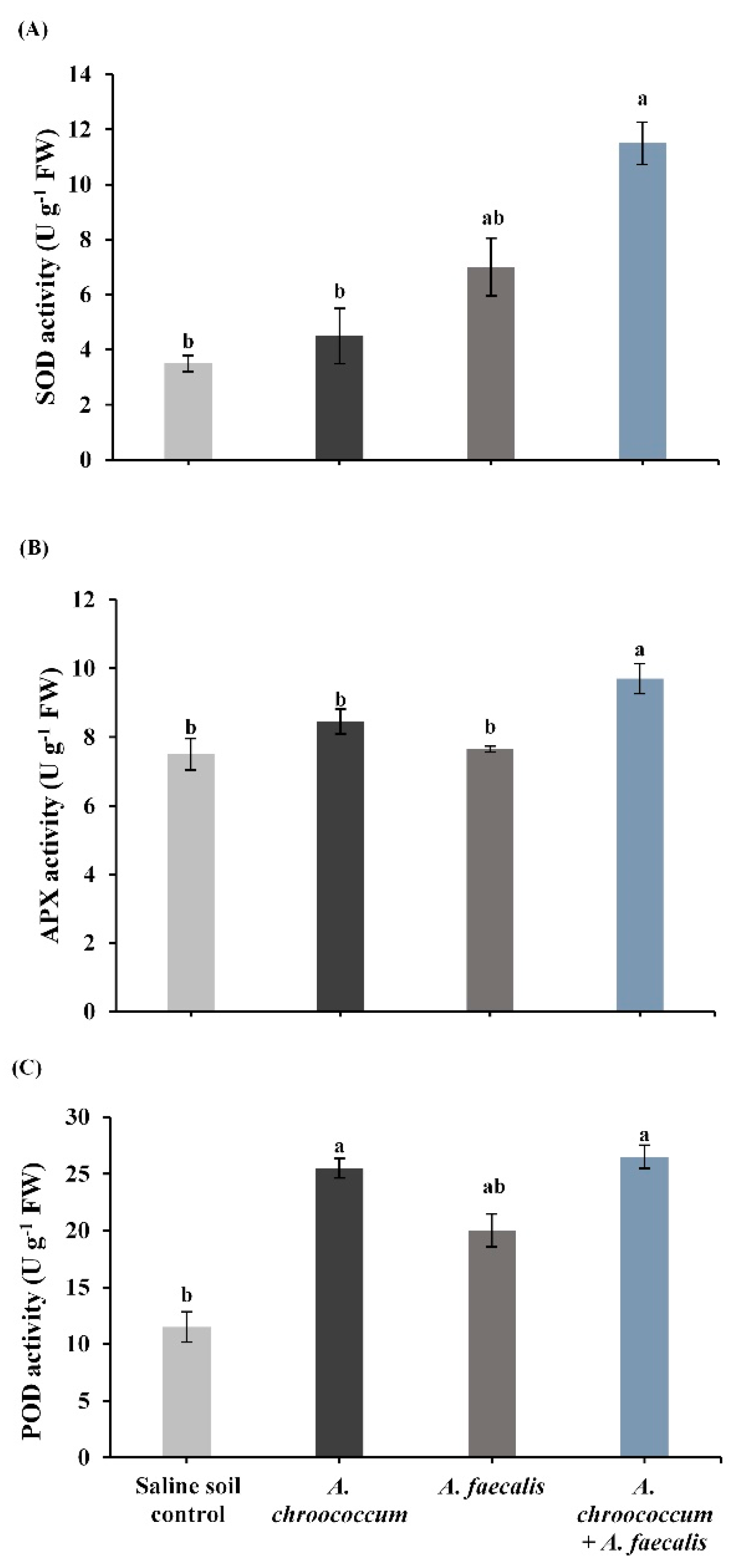
2.6. PGPR Stimulate Antioxidant Enzymes under Salinity-Stress Conditions
2.7. PGPR Regulate Mineral Uptake in Salinity-Stressed Canola Plants
3. Discussion
4. Materials and Methods
4.1. Isolation, Identification, and Description of PGPR (Salt-Tolerant Bacteria)
4.2. Pot Experiment
4.3. Microbiological Analysis of Canola Rhizosphere
4.4. Determination of Photosynthetic Pigments
4.5. Determination of Osmolyte Contents
4.6. Estimation of Malondialdehyde Content
4.7. Determination of Hydrogen Peroxide (H2O2) Content
4.8. Extraction and Assay of Antioxidant Enzymes
4.9. Determination of Mineral Contents
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Machado, R.M.A.; Serralheiro, R.P. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Srivastava, A.K.; El-sadek, M.S.A.; Kordrostami, M.; Tran, L.P. Titanium dioxide nanoparticles improve growth and enhance tolerance of broad bean plants under saline soil conditions. Land Degrad. Dev. 2018, 29, 1065–1073. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Mostofa, M.G.; Rahman, M.M.; Abdel-Farid, I.B.; Tran, L.-S.P. Extracts from yeast and carrot roots enhance maize performance under seawater-induced salt stress by altering physio-biochemical characteristics of stressed plants. J. Plant Growth Regul. 2019, 38, 966–979. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Alhmad, M.F.A.; Kordrostami, M.; Abo–Baker, A.-B.A.-E.; Zakir, A. Inoculation with Azospirillum lipoferum or Azotobacter chroococcum reinforces maize growth by improving physiological activities under saline conditions. J. Plant Growth Regul. 2020, 39, 1293–1306. [Google Scholar] [CrossRef]
- Jadhav, G.G.; Salunkhe, D.S.; Nerkar, D.P.; Bhadekar, R.K. Isolation and characterization of salt-tolerant nitrogen-fixing microorganisms from food. EurAsian J. Biosci. 2010, 4, 33–40. [Google Scholar] [CrossRef]
- Paul, D. Osmotic stress adaptations in rhizobacteria. J. Basic Microbiol. 2013, 53, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Osman, M.S.; Badawy, A.A.; Osman, A.I.; Latef, A.A.H.A. Ameliorative impact of an extract of the halophyte Arthrocnemum macrostachyum on growth and biochemical parameters of soybean under salinity stress. J. Plant Growth Regul. 2020. [Google Scholar] [CrossRef]
- Bowman, D.C.; Devitt, D.A.; Miller, W.W. The effect of moderate salinity on nitrate leaching from bermudagrass turf: A lysimeter study. Water Air Soil Pollut. 2006, 175, 49–60. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H. Changes of antioxidative enzymes in salinity tolerance among different wheat cultivars. Cereal Res. Commun. 2010, 38, 43–55. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Li, Y.; Li, X.; Han, W.-Y.; Chen, S. Epigallocatechin-3-gallate alleviates salinity-retarded seed germination and oxidative stress in tomato. J. Plant Growth Regul. 2018, 37, 1349–1356. [Google Scholar] [CrossRef]
- Shokri-Gharelo, R.; Noparvar, P.M. Molecular response of canola to salt stress: Insights on tolerance mechanisms. PeerJ 2018, 6, e4822. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.; Jha, P.N. The plant-growth-promoting bacterium Klebsiella sp. SBP-8 confers induced systemic tolerance in wheat (Triticum aestivum) under salt stress. J. Plant Physiol. 2015, 184, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, S.; Gaurav, A.K.; Srivastava, S.; Verma, J.P. Plant Growth-Promoting Bacteria: Biological Tools for the Mitigation of Salinity Stress in Plants. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Zhu, J.; Fan, Y.; Shabala, S.; Li, C.; Lv, C.; Guo, B.; Xu, R.; Zhou, M. Understanding mechanisms of salinity tolerance in barley by proteomic and biochemical analysis of near-isogenic lines. Int. J. Mol. Sci. 2020, 21, 1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Kohler, J.; Caravaca, F.; Carrasco, L.; Roldan, A. Contribution of Pseudomonas mendocina and Glomus intraradices to aggregate stabilization and promotion of biological fertility in rhizosphere soil of lettuce plants under field conditions. Soil Use Manag. 2006, 22, 298–304. [Google Scholar] [CrossRef]
- Glick, B.R. Modulation of plant ethylene levels by the bacterial enzyme ACC deaminase. FEMS Microbiol. Lett. 2005, 251, 1–7. [Google Scholar] [CrossRef]
- Kausar, R.; Shahzad, S.M. Effect of ACC-deaminase containing rhizobacteria on growth promotion of maize under salinity stress. J. Agric. Soc. Sci. 2006, 2, 216–218. [Google Scholar]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoğlu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2018, 32, 261–285. [Google Scholar] [CrossRef] [Green Version]
- Robson, R.L.; Jones, R.; Robson, R.M.; Schwartz, A.; Richardson, T.H. Azotobacter genomes: The genome of Azotobacter chroococcum NCIMB 8003 (ATCC 4412). PLoS ONE 2015, 10, e0127997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aly, M.M.; El-Sabbagh, S.M.; El-Shouny, W.A.; Ebrahim, M.K.H. Physiological response of Zea mays to NaCl stress with respect to Azotobacter chroococcum and Streptomyces niveus. Pak. J. Biol. Sci. 2003, 6, 2073–2080. [Google Scholar] [CrossRef]
- Ravikumar, S.; Kathiresan, K.; Alikhan, S.L.; Williams, G.P.; Gracelin, N.A.A. Growth of Avicennia marina and Ceriops decandra seedlings inoculated with halophilic azotobacters. J. Environ. Biol. 2007, 28, 601. [Google Scholar] [PubMed]
- Chaudhary, D.; Narula, N.; Sindhu, S.S.; Behl, R.K. Plant growth stimulation of wheat (Triticum aestivum L.) by inoculation of salinity tolerant Azotobacter strains. Physiol. Mol. Biol. Plants 2013, 19, 515–519. [Google Scholar] [CrossRef] [Green Version]
- Austin, B. The Family Alcaligenaceae; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2014; pp. 729–757. [Google Scholar]
- Neethu, S.; Vishnupriya, S.; Mathew, J. Isolation and functional characterisation of endophytic bacterial isolates from curcuma longa. Int. J. Pharm. Biol. Sci. 2016, 7, 455–464. [Google Scholar]
- Akhtar, S.; Ali, B. Evaluation of rhizobacteria as non-rhizobial inoculants for mung beans. Aust. J. Crop Sci. 2011, 5, 1723. [Google Scholar]
- Omer, A.M. Inducing plant resistance against salinity using some rhizobacteria. Egypt. J. Desert Res. 2017, 67, 187–208. [Google Scholar] [CrossRef]
- Ashraf, M.; McNeilly, T. Salinity tolerance in Brassica oilseeds. CRC Crit. Rev. Plant Sci. 2004, 23, 157–174. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H. Ameliorative effect of calcium chloride on growth, antioxidant enzymes, protein patterns and some metabolic activities of canola (Brassica napus L.) under seawater stress. J. Plant Nutr. 2011, 34, 1303–1320. [Google Scholar] [CrossRef]
- Lohani, N.; Jain, D.; Singh, M.B.; Bhalla, P.L. Engineering Multiple Abiotic Stress Tolerance in Canola, Brassica napus. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Hu, X.; Sullivan-Gilbert, M.; Gupta, M.; Thompson, S.A. Mapping of the loci controlling oleic and linolenic acid contents and development of fad2 and fad3 allele-specific markers in canola (Brassica napus L.). Theor. Appl. Genet. 2006, 113, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Milazzo, M.F.; Spina, F.; Vinci, A.; Espro, C.; Bart, J.C.J. Brassica biodiesels: Past, present and future. Renew. Sustain. Energy Rev. 2013, 18, 350–389. [Google Scholar] [CrossRef]
- Carré, P.; Pouzet, A. Rapeseed market, worldwide and in Europe. Ocl 2014, 21, D102. [Google Scholar] [CrossRef]
- Ghallab, K.H.; Sharaan, A.N. Selection in canola (Brassica napus L.) germplasm under conditions of newly reclaimed land. II. Salt tolerant selections. Egypt. J. Plant Breed 2002, 6, 15–30. [Google Scholar]
- El Sabagh, A.; Omar, A.E.; Saneoka, H.; Barutçular, C. Evaluation agronomic traits of canola (Brassica napus L.) under organic, bio-and chemical fertilizers. Dicle Univ. J. Inst. Nat. Appl. Sci. 2015, 4, 59–67. [Google Scholar]
- Abdel Latef, A.A.H.; Kordrostami, M.; Zakir, A.; Zaki, H.; Saleh, O.M. Eustress with H2O2 facilitates plant growth by improving tolerance to salt stress in two wheat cultivars. Plants 2019, 8, 303. [Google Scholar] [CrossRef] [Green Version]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Abdel Latef, A.A.H.; Chaoxing, H. Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Sci. Hortic. (Amsterdam) 2011, 127, 228–233. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Alhmad, M.F.A.; Abdelfattah, K.E. The possible roles of priming with ZnO nanoparticles in mitigation of salinity stress in lupine (Lupinus termis) plants. J. Plant Growth Regul. 2017, 36, 60–70. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Abu Alhmad, M.; Ahmad, S. Foliar application of fresh moringa leaf extract overcomes salt stress in fenugreek (Trigonella foenum-graecum) plants. Egypt. J. Bot. 2017, 57, 157–179. [Google Scholar]
- Abdel Latef, A.A.H.A.; Srivastava, A.K.; Saber, H.; Alwaleed, E.A.; Tran, L.-S.P. Sargassum muticum and Jania rubens regulate amino acid metabolism to improve growth and alleviate salinity in chickpea. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhise, K.K.; Dandge, P.B. Mitigation of salinity stress in plants using plant growth promoting bacteria. Symbiosis 2019, 79, 191–204. [Google Scholar] [CrossRef]
- Foo, E.; Plett, J.M.; Lopez-Raez, J.A.; Reid, D. The Role of plant hormones in plant-microbe symbioses. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Ashrafuzzaman, M.; Hossen, F.A.; Ismail, M.R.; Hoque, A.; Islam, M.Z.; Shahidullah, S.M.; Meon, S. Efficiency of plant growth-promoting rhizobacteria (PGPR) for the enhancement of rice growth. Afr. J. Biotechnol. 2009, 8. [Google Scholar]
- Mathew, M.; Obbard, J.P. Optimisation of the dehydrogenase assay for measurement of indigenous microbial activity in beach sediments contaminated with petroleum. Biotechnol. Lett. 2001, 23, 227–230. [Google Scholar] [CrossRef]
- Abou-Aly, H.E. Stimulatory effect of some yeast applications on response of tomato plants to inoculation with biofertilizers. Ann. Agric. Sci. Moshtohor 2005, 43, 595–609. [Google Scholar]
- Fatima, T.; Mishra, I.; Verma, R.; Kumar, N. Mechanisms of halotolerant plant growth promoting Alcaligenes sp. involved in salt tolerance and enhancement of the growth of rice under salinity stress. 3 Biotech 2020. [Google Scholar] [CrossRef]
- Li, H.; Lei, P.; Pang, X.; Li, S.; Xu, H.; Xu, Z.; Feng, X. Enhanced tolerance to salt stress in canola (Brassica napus L.) seedlings inoculated with the halotolerant Enterobacter cloacae HSNJ4. Appl. Soil Ecol. 2017, 119, 26–34. [Google Scholar] [CrossRef]
- Shukla, P.S.; Agarwal, P.K.; Jha, B. Improved salinity tolerance of Arachishypogaea (L.) by the interaction of halotolerant plant-growth-promoting rhizobacteria. J. Plant Growth Regul. 2012, 31, 195–206. [Google Scholar] [CrossRef]
- Ullah, S.; Bano, A. Isolation of plant-growth-promoting rhizobacteria from rhizospheric soil of halophytes and their impact on maize (Zea mays L.) under induced soil salinity. Can. J. Microbiol. 2015, 61, 307–313. [Google Scholar] [CrossRef]
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci. Rep. 2016, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orhan, F. Alleviation of salt stress by halotolerant and halophilic plant growth-promoting bacteria in wheat (Triticum aestivum). Braz. J. Microbiol. 2016, 47, 621–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majeed, A.; Abbasi, M.K.; Hameed, S.; Imran, A.; Rahim, N. Isolation and characterization of plant growth-promoting rhizobacteria from wheat rhizosphere and their effect on plant growth promotion. Front. Microbiol. 2015, 6, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etesami, H.; Glick, B.R. Halotolerant plant growth–promoting bacteria: Prospects for alleviating salinity stress in plants. Environ. Exp. Bot. 2020, 104124. [Google Scholar] [CrossRef]
- Yildirim, E.; Turan, M.; Ekinci, M.; Dursun, A.; Cakmakci, R. Plant growth promoting rhizobacteria ameliorate deleterious effect of salt stress on lettuce. Sci. Res. Essays 2011, 6, 4389–4396. [Google Scholar]
- Kang, S.-M.; Khan, A.L.; Waqas, M.; You, Y.-H.; Kim, J.-H.; Kim, J.-G.; Hamayun, M.; Lee, I.-J. Plant growth-promoting rhizobacteria reduce adverse effects of salinity and osmotic stress by regulating phytohormones and antioxidants in Cucumis sativus. J. Plant Interact. 2014, 9, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Aslam, F.; Ali, B. Halotolerant bacterial diversity associated with Suaeda fruticosa (L.) forssk. improved growth of maize under salinity stress. Agronomy 2018, 8, 131. [Google Scholar] [CrossRef] [Green Version]
- Pinnola, A.; Staleva-musto, H.; Capaldi, S.; Ballottari, M.; Bassi, R. Biochimica et Biophysica Acta Electron transfer between carotenoid and chlorophyll contributes to quenching in the LHCSR1 protein from Physcomitrella patens. Biochimica et Biophysica Acta (BBA)—Bioenergetics 2016, 1870–1878. [Google Scholar] [CrossRef]
- Enebe, M.C.; Babalola, O.O. The influence of plant growth-promoting rhizobacteria in plant tolerance to abiotic stress: A survival strategy. Appl. Microbiol. Biotechnol. 2018, 102, 7821–7835. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Liu, Y.; Wu, G.; Veronican, K.; Shen, Q.; Zhang, N. Induced maize salt tolerance by rhizosphere inoculation of Bacillus amyloliquefaciens SQR9. Physiol. Plant. 2016, 34–44. [Google Scholar] [CrossRef]
- Hmaeid, N.; Wali, M.; Mahmoud, O.M.-B.; Pueyo, J.J.; Ghnaya, T.; Abdelly, C. Efficient rhizobacteria promote growth and alleviate NaCl-induced stress in the plant species Sulla carnosa. Appl. Soil Ecol. 2019, 133, 104–113. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Lugtenberg, B. Use of plant growth-promoting rhizobacteria to alleviate salinity stress in plants. In Use of Microbes for the Alleviation of Soil Stresses; Springer: New York, NY, USA, 2014; Volume 1, pp. 73–96. [Google Scholar]
- Pan, J.; Peng, F.; Xue, X.; You, Q.; Zhang, W.; Wang, T.; Huang, C. The growth promotion of two salt-tolerant plant groups with PGPR inoculation: A meta-analysis. Sustainability 2019, 11, 378. [Google Scholar] [CrossRef] [Green Version]
- Hamdia, M.A.E.-S.; Shaddad, M.A.K.; Doaa, M.M. Mechanisms of salt tolerance and interactive effects of Azospirillum brasilense inoculation on maize cultivars grown under salt stress conditions. Plant Growth Regul. 2004, 44, 165–174. [Google Scholar] [CrossRef]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira, J.A.G.; de Almeida Viégas, R.; da Rocha, I.M.A.; Moreira, A.C.D.O.M.; de Azevedo Moreira, R.; Oliveira, J.T.A. Proline accumulation and glutamine synthetase activity are increased by salt-induced proteolysis in cashew leaves. J. Plant Physiol. 2003, 160, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Rai, S.; Luthra, R.; Kumar, S. Salt-tolerant mutants in glycophytic salinity response (GSR) genes in Catharanthus roseus. Theor. Appl. Genet. 2003, 106, 221–230. [Google Scholar] [CrossRef]
- Han, H.S.; Lee, K.D. Physiological responses of soybean-inoculation of Bradyrhizobium japonicum with PGPR in saline soil conditions. Res. J. Agric. Biol. Sci. 2005, 1, 216–221. [Google Scholar]
- Khan, M.H.; Panda, S.K. Alterations in root lipid peroxidation and antioxidative responses in two rice cultivars under NaCl-salinity stress. Acta Physiol. Plant. 2008, 30, 81. [Google Scholar] [CrossRef]
- Samaddar, S.; Chatterjee, P.; Choudhury, A.R.; Ahmed, S.; Sa, T. Interactions between Pseudomonas spp. and their role in improving the red pepper plant growth under salinity stress. Microbiol. Res. 2019, 219, 66–73. [Google Scholar] [CrossRef]
- Abbas, T.; Pervez, M.A.; Ayyub, C.M.; Ahmad, R. Assessment of morphological, antioxidant, biochemical and ionic responses of salttolerant and salt-sensitive okra (Abelmoschus esculentus) under saline regime. Pakistan J. Life Soc. Sci. 2013, 11, 147–153. [Google Scholar]
- Abdelgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R. High Salinity Induces Different Oxidative Stress and Antioxidant Responses in Maize Seedlings Organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, F.; Yasmeen, T.; Arif, M.S.; Ali, S.; Ali, B.; Hameed, S.; Zhou, W. Plant growth promoting bacteria confer salt tolerance in Vigna radiata by up-regulating antioxidant defense and biological soil fertility. Plant Growth Regul. 2016, 80, 23–36. [Google Scholar] [CrossRef]
- Etesami, H.; Mirsyed Hosseini, H.; Alikhani, H.A. In planta selection of plant growth promoting endophytic bacteria for rice (Oryza sativa L.). J. Soil Sci. Plant Nutr. 2014, 14, 491–503. [Google Scholar] [CrossRef]
- Jaiswal, D.K.; Verma, J.P.; Prakash, S.; Meena, V.S.; Meena, R.S. Potassium as an important plant nutrient in sustainable agriculture: A state of the art. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Springer: New York, NY, USA, 2016; pp. 21–29. [Google Scholar]
- Abd-el-Malek, Y.; Ishac, Y.Z. Evaluation of methods used in counting azotobacters. J. Appl. Bacteriol. 1968, 31, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Roskot, N.; Steidle, A.; Eberl, L.; Zock, A.; Smalla, K. Plant-dependent genotypic and phenotypic diversity of antagonistic rhizobacteria isolated from different Verticillium host plants. Appl. Environ. Microbiol. 2002, 68, 3328–3338. [Google Scholar] [CrossRef] [Green Version]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescein. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar]
- Samasegaran, P.; Hoben, H.; Halliday, J. The NIFTAL (Nitrogen Fixation in Tropical Agricultural Legumes) Manual for Methods in Legume Rhizobium Technology; US Agency for International Development: Washington, DC, USA, 1982.
- Allen, O.N. Experiments in Soil Bacteriology/Soil Bacteriology; Burgess: Minneapolis, MN, USA, 1959. [Google Scholar]
- Pepper, I.L.; Gerba, C.P.; Brendecke, J.W. Environmental Microbiology: A Laboratory Manual; Academic Press: Cambridge, MA, USA, 1995; ISBN 0125506554. [Google Scholar]
- Vernon, L.P.; Seely, G.R. The Chlorophylls; Academic Press: New York, NY, USA, 1966. [Google Scholar]
- Lichtenthaler, H.K. [34] Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Umbreit, W.W.; Burris, R.H.; Stauffer, J.F. Manometric Techniques: A Manual Describing Methods Applicable to the Study of Tissue Metabolism; Burgess Publishing Co.: Minneapolis, MN, USA, 1957. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Qu, W.J. The guidance of plant physiology experiments. Chin. Agric. Sci. Technol. Press. Beijing 2004, 120–135. [Google Scholar]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-X.; Asada, K. Inactivation of ascorbate peroxidase by thiols requires hydrogen peroxide. Plant Cell Physiol. 1992, 33, 117–123. [Google Scholar]
- Bergmeyer, H.U. Methods of Enzymatic Analysis; Verlag Chemie: Hoboken, NJ, USA, 1974; ISBN 3527255303. [Google Scholar]
- Williams, V.; Twine, S. Flame photometric method for sodium, potassium and calcium. Mod. Methods Plant Anal. 1960, 5, 3–5. [Google Scholar]
- Pregl, F. Quantitative Organic Microanalysis; JA Churchill Ltd.: London, UK, 1945. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | TBC * × 105 CFU/gm Dry Soil | TBC Increasing % | NFC × 103 CFU/gm Dry Soil | NFC Increasing % | Dehydrogenase (μg TPF/g Dry Soil/24 h) | Dehydrogenase Increasing % |
|---|---|---|---|---|---|---|
| Saline soil control | 78 | 34.4 | 2.3 | 109 | 112 | 26.9 |
| A. chroococcum | 96 | 65.5 | 3.6 | 227 | 144 | 63.3 |
| A. faecalis | 122 | 110.3 | 3.9 | 254 | 191 | 116.6 |
| A. chroococcum + A. faecalis | 124 | 113.7 | 3.9 | 254 | 187 | 112 |
| Treatments | Soluble Sugars | Soluble Proteins | Proline |
|---|---|---|---|
| Saline soil control | 116.7 ± 3.85 b | 16.9 ± 0.26 b | 0.64 ± 0.04 a |
| A. chroococcum | 135.3 ± 11.85 b | 24.6 ± 0.48 a | 0.62 ± 0.009 ab |
| A. faecalis | 157.1 ± 9.76 b | 25.5 ± 0.70 a | 0.59 ± 0.006 bc |
| A. chroococcum + A. faecalis | 226.5 ± 6.69 a | 26.2 ± 0.38 a | 0.57 ± 0.023 c |
| Treatments | Na | K | N | Ca | Mg |
|---|---|---|---|---|---|
| Saline soil control | 14.81 ± 0.96 a | 2.34 ± 0.10 d | 1.34 ± 0.06 d | 0.22 ± 0.05 b | 0.39 ± 0.04 b |
| A. chroococcum | 7.36 ± 0.35 c | 3.56 ± 0.06 c | 2.12 ± 0.09 b | 0.25 ± 0.04 b | 0.31 ± 0.03 b |
| A. faecalis | 9.24 ± 0.70 b | 4.17 ± 0.08 b | 1.97 ± 0.07 c | 0.30 ± 0.04 b | 0.71 ± 0.05 a |
| A. chroococcum + A. faecalis | 6.37 ± 0.11 d | 5.66 ± 0.09 a | 2.41 ± 0.05 a | 0.62 ± 0.08 a | 0.61 ± 0.02 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel Latef, A.A.H.; Omer, A.M.; Badawy, A.A.; Osman, M.S.; Ragaey, M.M. Strategy of Salt Tolerance and Interactive Impact of Azotobacter chroococcum and/or Alcaligenes faecalis Inoculation on Canola (Brassica napus L.) Plants Grown in Saline Soil. Plants 2021, 10, 110. https://doi.org/10.3390/plants10010110
Abdel Latef AAH, Omer AM, Badawy AA, Osman MS, Ragaey MM. Strategy of Salt Tolerance and Interactive Impact of Azotobacter chroococcum and/or Alcaligenes faecalis Inoculation on Canola (Brassica napus L.) Plants Grown in Saline Soil. Plants. 2021; 10(1):110. https://doi.org/10.3390/plants10010110
Chicago/Turabian StyleAbdel Latef, Arafat Abdel Hamed, Amal M. Omer, Ali A. Badawy, Mahmoud S. Osman, and Marwa M. Ragaey. 2021. "Strategy of Salt Tolerance and Interactive Impact of Azotobacter chroococcum and/or Alcaligenes faecalis Inoculation on Canola (Brassica napus L.) Plants Grown in Saline Soil" Plants 10, no. 1: 110. https://doi.org/10.3390/plants10010110
APA StyleAbdel Latef, A. A. H., Omer, A. M., Badawy, A. A., Osman, M. S., & Ragaey, M. M. (2021). Strategy of Salt Tolerance and Interactive Impact of Azotobacter chroococcum and/or Alcaligenes faecalis Inoculation on Canola (Brassica napus L.) Plants Grown in Saline Soil. Plants, 10(1), 110. https://doi.org/10.3390/plants10010110