Effects of Water Deficits on Prosopis tamarugo Growth, Water Status and Stomata Functioning



Abstract
:1. Introduction
2. Results
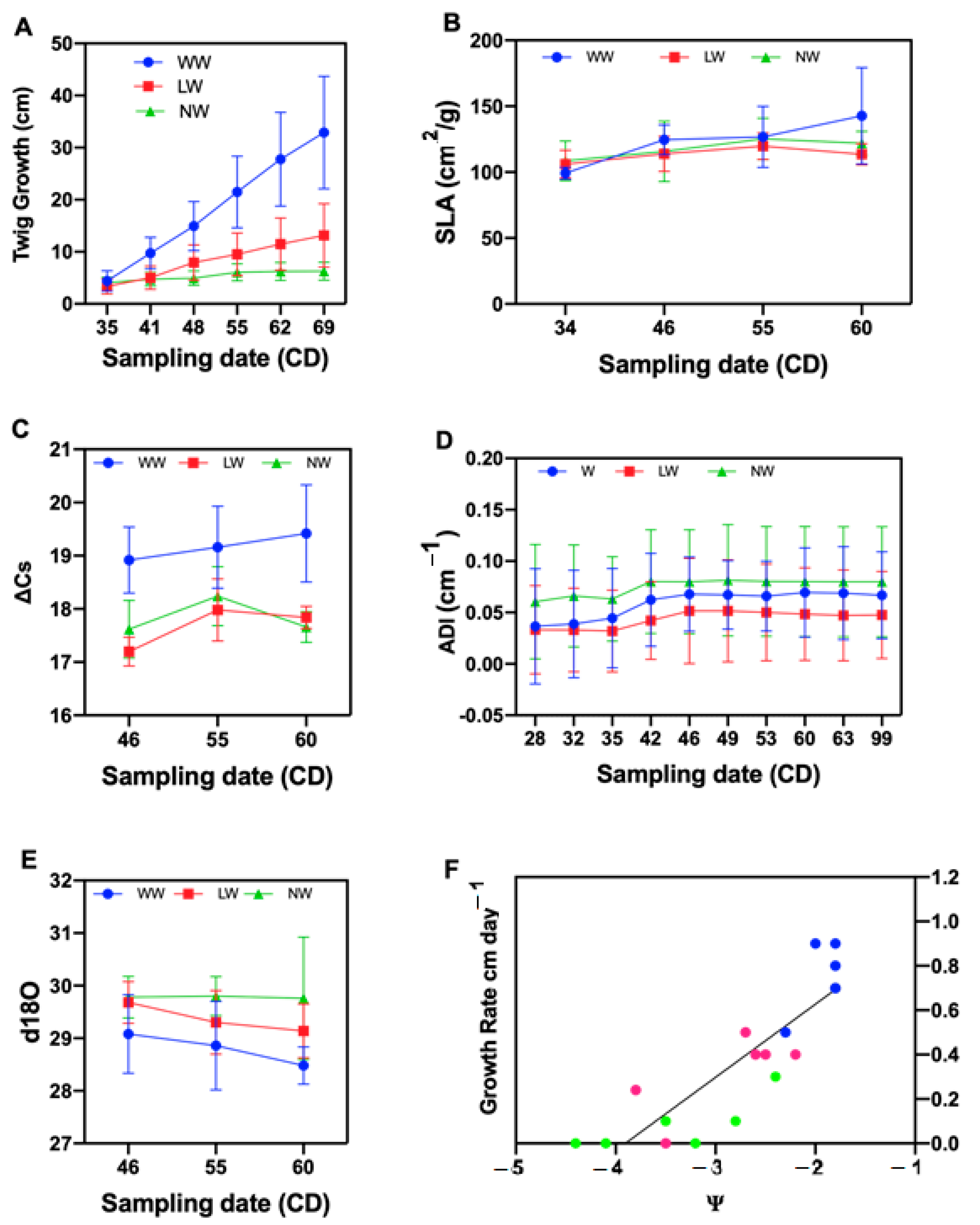
2.1. Effect of Water Stress on Tamarugo Growth, Isotopic Discrimination of 13C and Enrichment of 18O
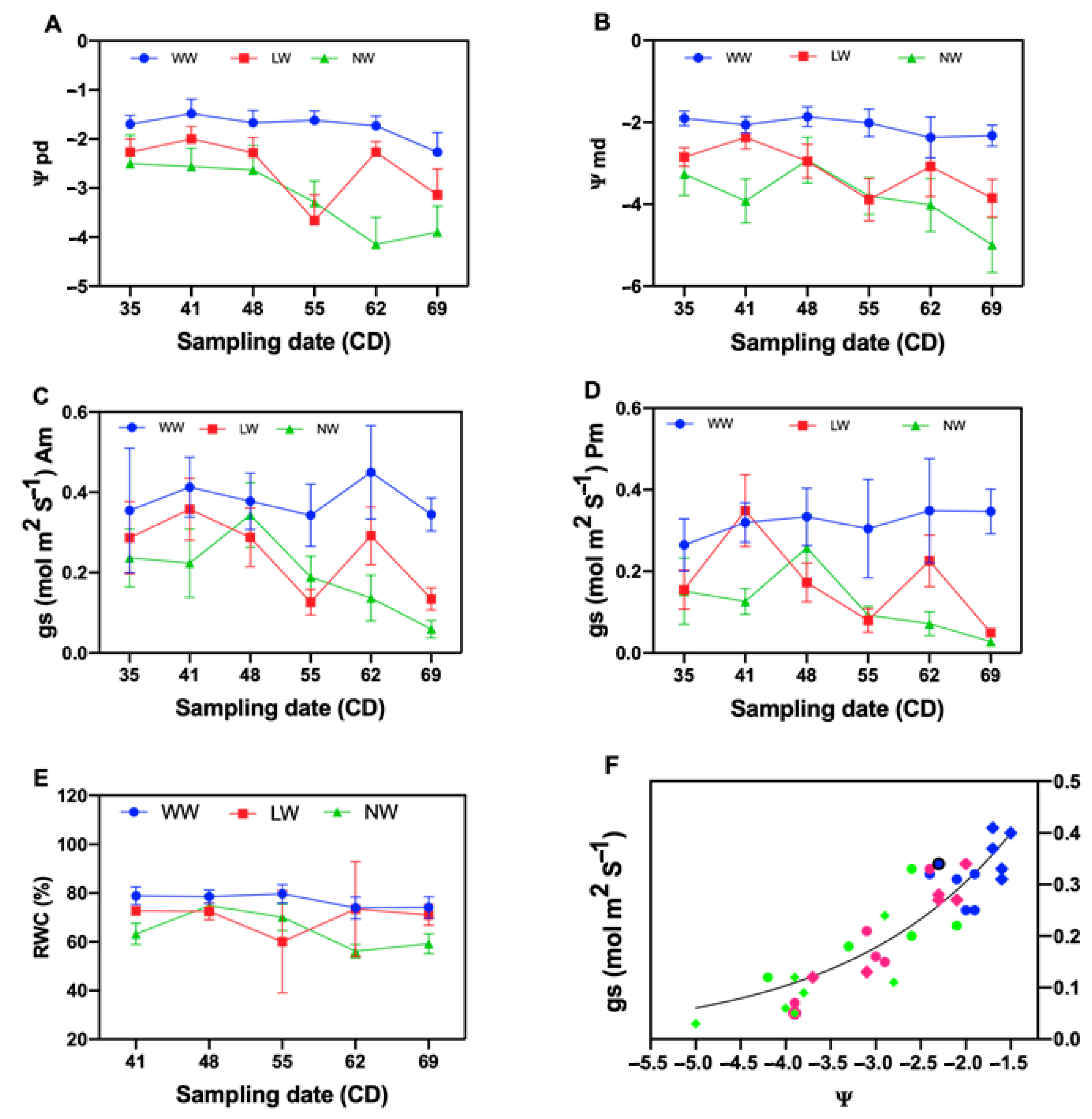
2.2. Effect of Water Stress on Tamarugo Water Status and Stomata Functioning
3. Discussion
3.1. Tamarugo Water Status and Stomatal Functioning at Different Water Condition Levels
3.2. Tamarugo Growth at Different Water Levels
4. Materials and Methods
4.1. Site Description
4.2. Study Design
4.3. Measured Variables
4.4. Branching Architecture
4.5. Specific Leaf Area
4.6. Twig Growth Rate (TGR)
4.7. Relative Water Content (RWC)
4.8. Predawn Leaf (Ψpd) and Midday Leaf Water Potential (Ψmd)
4.9. Stomata Conductance
4.10. Isotopic Discrimination of 13C and Enrichment of 18O
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calderon, G.; Garrido, M.; Acevedo, E. Prosopis tamarugo Phil.: A native tree from the Atacama Desert groundwater table depth thresholds for conservation. Rev. Chil. Hist. Nat. 2015, 88, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Frelich, L. Forest Dynamics and Disturbance Regimes: Studies from Temperate Evergreen-Deciduous Forest; Cambridge University Press: Cambridge, UK, 2002; Volume 117, pp. 502–503. [Google Scholar]
- Rojas, R.; Dessargues, A. Groundwater flow modelling of the regional acquifer of the Pampa del Tamarugal, northern Chile. Hydrogeol. J. 2007, 15, 537–551. [Google Scholar] [CrossRef] [Green Version]
- Tardieu, F.; Simonneau, T. Variability among species of stomatal control under fluctuating soil water status and evaporative demand: Modelling isohydric and anisohydric behaviours. J. Exp. Bot. 1998, 49, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Rood, S.B.; Zanewich, K.; Stefura, C.; Mahoney, J.M. Influence of water table decline on growth allocation and endogenous gibberellins in black cottonwood. Tree Physiol. 2000, 20, 831–836. [Google Scholar] [CrossRef] [Green Version]
- Horton, J.L.; Kolb, T.E.; Hart, S.C. Physiological response to groundwater depth varies among species and with river flow regulation. Ecol. Appl. 2001, 11, 1046–1059. [Google Scholar] [CrossRef]
- Cooper, D.J.; D’Amico, D.R.; Scott, M.L. Physiological and morphological response patterns of Populus deltoides to alluvial groundwater pumping. Environ. Manag. 2003, 31, 215–226. [Google Scholar] [CrossRef]
- Lambers, H.; Champin, F.S.I.; Pons, T.L. Photosynthesis, respiration, and long distance transport. In Plant Physiological Ecology; Lambers, H., Chapin, F.S., III, Pons, T.L., Eds.; Springer: Berlin, Germany, 1998; pp. 10–95. [Google Scholar] [CrossRef]
- Baohua, G.; Ying, G.; Meiying, F.; Xiaoyin, N.; Yijun, L.; Jie, C. Phenotypic plasticity of growth and morphology in Mosla chinensis responds to diverse relative soil water content. Acta Ecol. Sin. 2003, 23, 259–563. [Google Scholar]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, T.C.; Acevedo, E. Plant responses to water deficits, water-use efficiency, and drought resistance. Agric. Meteorol. 1974. [Google Scholar] [CrossRef]
- Flexas, J.; Diaz-Espejo, A.; Gago, J.; Gallé, A.; Galmés, J.; Gulías, J.; Medrano, H. Photosynthetic limitations in Mediterranean plants: A review. Environ. Exp. Bot. 2014, 103, 12–23. [Google Scholar] [CrossRef]
- Time, A.; Garrido, M.; Acevedo, E. Water relations and growth response to drought stress of Prosopis tamarugo Phil. A review. J. Soil Sci. Plant Nutr. 2018, 18, 329–343. [Google Scholar] [CrossRef]
- Lehner, G.; Delatorre, J.; Lütz, C.; Cardemil, L. Field studies on the photosynthesis of two desert Chilean plants: Prosopis chilensis and Prosopis tamarugo. J. Photochem. Photobiol. B Biol. 2001. [Google Scholar] [CrossRef]
- Delatorre, J.; Pinto, M.; Cardemil, L. Effects of water stress and high temperature on photosynthetic rates of two species of Prosopis. J. Photochem. Photobiol. B Biol. 2008. [Google Scholar] [CrossRef] [PubMed]
- Chávez, R.O.; Clevers, J.G.P.W.; Herold, M.; Acevedo, E.; Ortiz, M. Assessing water stress of desert tamarugo trees using in situ data and very high spatial resolution remote sensing. Remote Sens. 2013, 5, 5064–5088. [Google Scholar] [CrossRef] [Green Version]
- Barnes, F.J. Carbon Gain and Water Relations in Pinyon-Juniper Habitat Types (Photosynthesis, Water Stress, Gradient Analysis, Ecophysiology); New Mexico State University: Las Cruces, NM, USA, 1987. [Google Scholar]
- Loewenstein, N.J.; Pallardy, S.G. Drought tolerance, xylem sap abscisic acid and stomatal conductance during soil drying: A comparison of canopy trees of three temperate deciduous angiosperms. Tree Physiol. 1998, 18, 431–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franks, P.J.; Drake, P.L.; Froend, R.H. Anisohydric but isohydrodynamic: Seasonally constant plant water potential gradient explained by a stomatal control mechanism incorporating variable plant hydraulic conductance. Plant Cell Environ. 2007, 30, 19–30. [Google Scholar] [CrossRef]
- West, A.G.; Hultine, K.R.; Sperry, J.S.; Bush, S.E.; Ehleringer, J.R. Transpiration and hydraulic strategies in a piñon-juniper woodland. Ecol. Appl. 2008. [Google Scholar] [CrossRef] [Green Version]
- Garrido, M.; Silva, P.; Acevedo, E. Water relations and foliar isotopic composition of prosopis tamarugo phil., an endemic tree of the atacama desert growing at three levels of water table depth. Front. Plant Sci. 2016, 7, 375. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Vilagrosa, A.; Peñuelas, J.; Ogaya, R.; Camarero, J.; Corcuera, L.; Sisó, S.; Gil-Pelegrín, E. CAPÍTULO 6 Estrés hídrico: Ecofisiología y escalas de la sequía. Water 2004, 163–190. [Google Scholar]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust. J. Plant Physiol. 1982. [Google Scholar] [CrossRef]
- Hasselquist, N.J.; Allen, M.F.; Santiago, L.S. Water relations of evergreen and drought-deciduous trees along a seasonally dry tropical forest chronosequence. Oecologia 2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poorter, H.; Remkes, C. Leaf area ratio and net assimilation rate of 24 wild species differing in relative growth rate. Oecologia 1990, 83, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Poorter, H. Inherent Variation in Growth Rate Between Higher Plants: A Search for Physiological Causes and Ecological Consequences. Adv. Ecol. Res. 1992, 34, 283–362. [Google Scholar]
- Billings, S.A.; Boone, A.S.; Stephen, F.M. Tree-ring δ13C and δ18O, leaf δ13C and wood and leaf N status demonstrate tree growth strategies and predict susceptibility to disturbance. Tree Physiol. 2016. [Google Scholar] [CrossRef] [Green Version]
- Barbour, M.M. Stable oxygen isotope composition of plant tissue: A review. Funct. Plant Biol. 2007, 34, 83–94. [Google Scholar] [CrossRef]
- Westgate, M.E.; Boyer, J.S. Osmotic adjustment and the inhibition of leaf, root, stem and silk growth at low water potentials in maize. Planta 1985. [Google Scholar] [CrossRef]
- Passioura, J.B. Drought and drought tolerance. Plant Growth Regul. 1996. [Google Scholar] [CrossRef]
- Novoa, R.; Villaseca, S.; del Canto, P.; Sierra, C.; del Pozo, A. Mapa Agroclimático de Chile; Instituto de Investigaciones Agropecuarias: Santiago, Chile, 1989. [Google Scholar]
- Pérez-Harguindeguy, N.; Diaz, S.; Gamier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Comwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Mitchell, P.J.; O’Grady, A.P.; Tissue, D.T.; White, D.A. Drought response strategies define the relative contributions of hydraulic dysfunction and carbohydrate depletion during tree mortality. New Phytol. 2013, 197, 862–872. [Google Scholar] [CrossRef]
- Pietragalla, J.; Mullan, D. Leaf relative water content. In Physiological Breeding II: A Field Guide to Wheat Phenotyping; CIMMYT: Texcoco, Mexico, 2012; pp. 25–27. [Google Scholar]



{kind=link}
{kind=link}
| Growth Parameters/Water Conditions | TGR (cm day−1) | SLA (cm2 g−1) | BA (ADI) (cm−1) | Δ 13C (‰) | δ18O (‰) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| WW | 0.80 ±0.06 | a | 123.3 ±3.14 | a | 0.06 ±0.01 | b | 19.2 ±0.15 | a | 28.8 ±0.17 | b |
| LW | 0.34 ±0.06 | b | 113.3 ±3.14 | b | 0.04 ±0.01 | b | 17.7 ±0.15 | b | 29.4 ±0.17 | a |
| NW | 0.18 ±0.06 | c | 117.9 ±3.14 | b | 0.08 ±0.01 | a | 17.8± 0.26 | b | 29.8± 0.17 | a |
| Water status Parameters/Water Conditions | PD Ψ (Mpa) | MD Ψ (Mpa) | RWC (%) | am gs (mol m2 S−1) | pm gs (mol m2 S−1) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| WW | −1.75 ± 0.07 | a | −2.09 ± 0.09 | a | 77.04 ± 1.65 | a | 0.38 ± 0.01 | a | 0.32 ± 0.01 | a |
| LW | −2.60 ± 0.07 | b | −3.17 ± 0.09 | b | 69.94 ± 1.65 | b | 0.25 ± 0.01 | b | 0.17 ± 0.01 | b |
| NW | −3.17 ± 0.07 | c | −3.82 ± 0.09 | c | 64.70 ± 1.65 | c | 0.20 ± 0.01 | c | 0.12 ± 0.01 | c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Time, A.; Acevedo, E. Effects of Water Deficits on Prosopis tamarugo Growth, Water Status and Stomata Functioning. Plants 2021, 10, 53. https://doi.org/10.3390/plants10010053
Time A, Acevedo E. Effects of Water Deficits on Prosopis tamarugo Growth, Water Status and Stomata Functioning. Plants. 2021; 10(1):53. https://doi.org/10.3390/plants10010053
Chicago/Turabian StyleTime, Alson, and Edmundo Acevedo. 2021. "Effects of Water Deficits on Prosopis tamarugo Growth, Water Status and Stomata Functioning" Plants 10, no. 1: 53. https://doi.org/10.3390/plants10010053
APA StyleTime, A., & Acevedo, E. (2021). Effects of Water Deficits on Prosopis tamarugo Growth, Water Status and Stomata Functioning. Plants, 10(1), 53. https://doi.org/10.3390/plants10010053