
Plant Growth Regulators INCYDE and TD-K Underperform in Cereal Field Trials



Abstract
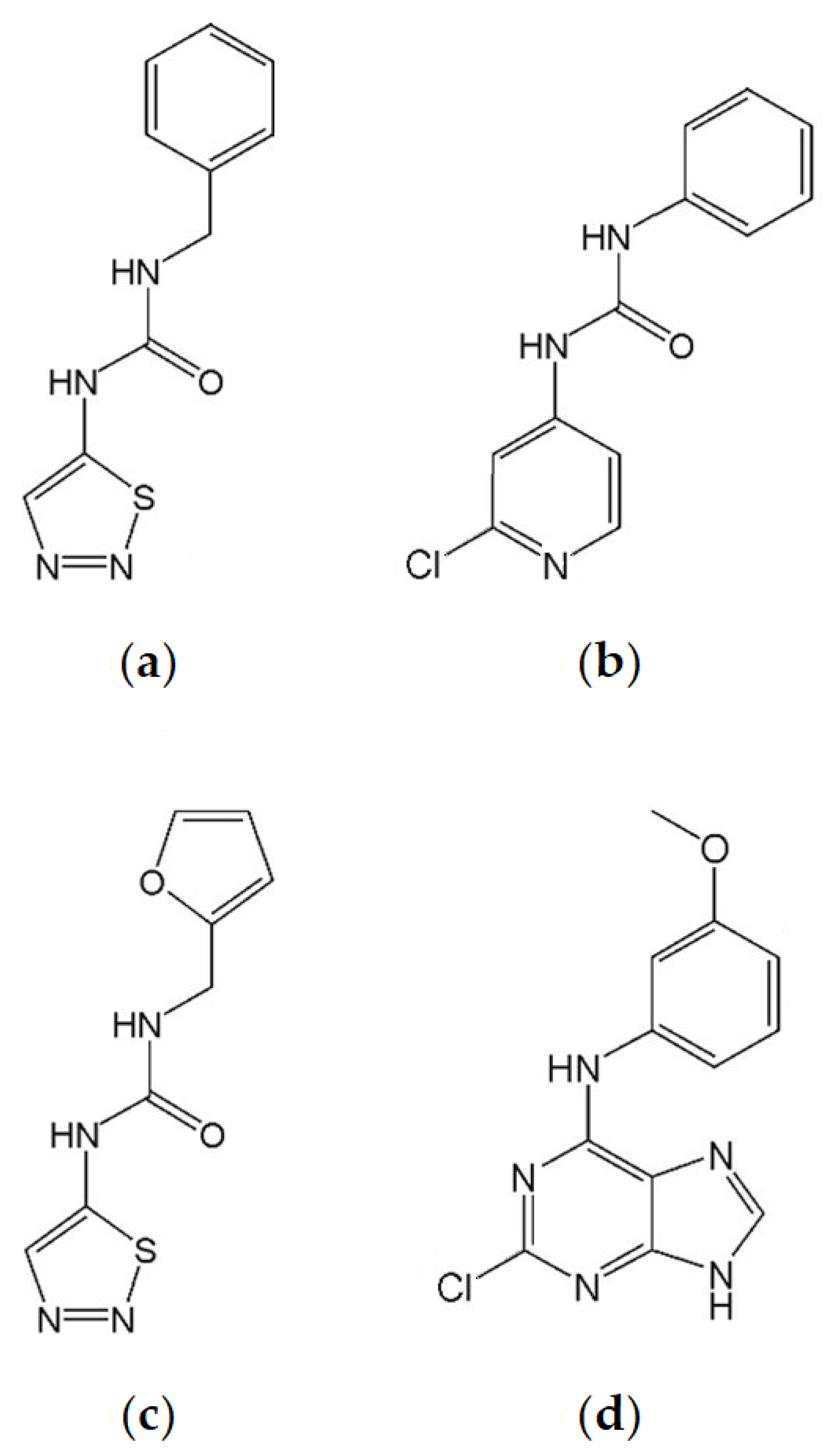
:1. Introduction
2. Results
2.1. Field Trial Analyses
2.2. LC–MS/MS Analyses in Grain
3. Discussion
4. Materials and Methods
4.1. Field Trials
4.2. Plant Material
4.2.1. Yield and Protein Composition Analyses
4.2.2. LC–MS/MS Analyses
4.3. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baldos, U.L.C.; Hertel, T.W. Global food security in 2050: The role of agricultural productivity and climate change. Aust. J. Agric. Econ. 2014, 58, 554–570. [Google Scholar] [CrossRef] [Green Version]
- Schmidhuber, J.; Tubiello, F.N. Global food security under climate change. Proc. Natl. Acad. Sci. USA 2007, 104, 19703–19708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nellemann, C.; MacDevette, M.; Manders, T.; Eickhout, B.; Svihus, B.; Prins, A.G.; Kaltenborn, B.P. The Environmental Food Crisis: The Environment’s Role in Averting Future Food Crises: A UNEP Rapid Response Assessment; UNEP/Earthprint: Stevenage, UK, 2009. [Google Scholar]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- USDA Foreign Agricultural Service. Grain: World Markets and Trade; USDA Foreign Agricultural Service: Washington, DC, USA, 2021.
- Langer, R.H.M.; Prasad, P.C.; Laude, H.M. Effects of kinetin on tiller bud elongation in wheat (Triticum aestivum L.). Ann. Bot. 1973, 37, 565–571. [Google Scholar] [CrossRef]
- Darwinkel, A. Ear development and formation of grain yield in winter wheat. Neth. J. Agric. Sci. 1980, 28, 156–163. [Google Scholar] [CrossRef]
- Harrison, M.A.; Kaufman, P.B. Hormonal regulation of lateral bud (tiller) release in oats (Avena sativa L.). Plant Physiol. 1980, 66, 1123–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhao, J.; Song, J.; Jameson, P.E. Cytokinin dehydrogenase: A genetic target for yield improvement in wheat. Plant Biotechnol. J. 2020, 18, 614–630. [Google Scholar] [CrossRef] [PubMed]
- Van Daele, I.; Gonzalez, N.; Vercauteren, I.; de Smet, L.; Inzé, D.; Roldán-Ruiz, I.; Vuylsteke, M. A comparative study of seed yield parameters in Arabidopsis thaliana mutants and transgenics. Plant Biotechnol. J. 2012, 10, 488–500. [Google Scholar] [CrossRef]
- Guo, T.; Chen, K.; Dong, N.-Q.; Shi, C.-L.; Ye, W.-W.; Gao, J.-P.; Shan, J.-X.; Lin, H.-X. GRAIN SIZE AND NUMBER1 negatively regulates the OsMKKK10-OsMKK4-OsMPK6 cascade to coordinate the trade-off between grain number per panicle and grain size in rice. Plant Cell 2018, 30, 871–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holubová, K.; Hensel, G.; Vojta, P.; Tarkowski, P.; Bergougnoux, V.; Galuszka, P. Modification of barley plant productivity through regulation of cytokinin content by reverse-genetics approaches. Front. Plant Sci. 2018, 9, 1676. [Google Scholar] [CrossRef] [Green Version]
- Jabłoński, B.; Ogonowska, H.; Szala, K.; Bajguz, A.; Orczyk, W.; Nadolska-Orczyk, A. Silencing of TaCKX1 mediates expression of other TaCKX genes to increase yield parameters in wheat. Int. J. Mol. Sci. 2020, 21, 4809. [Google Scholar] [CrossRef]
- Jabłoński, B.; Szala, K.; Przyborowski, M.; Bajguz, A.; Chmur, M.; Gasparis, S.; Orczyk, W.; Nadolska-Orczyk, A. TaCKX2.2 genes coordinate expression of other TaCKX family members, regulate phytohormone content and yield-related traits of wheat. Int. J. Mol. Sci. 2021, 22, 4142. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, P.W.; Kirkegaard, J.A.; Lilley, J.M.; Gregory, P.J.; Rebetzke, G.J. A tillering inhibition gene influences root–shoot carbon partitioning and pattern of water use to improve wheat productivity in rainfed environments. J. Exp. Bot. 2016, 67, 327–340. [Google Scholar] [CrossRef]
- Kebrom, T.H.; Chandler, P.M.; Swain, S.M.; King, R.W.; Richards, R.A.; Spielmeyer, W. Inhibition of tiller bud outgrowth in the tin mutant of wheat is associated with precocious internode development. Plant Physiol. 2012, 160, 308–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, T.; Motyka, V.; Strnad, M.; Schmülling, T. Regulation of plant growth by cytokinin. Proc. Natl. Acad. Sci. USA 2001, 98, 10487–10492. [Google Scholar] [CrossRef] [Green Version]
- Werner, T.; Motyka, V.; Laucou, V.; Smets, R.; Van Onckelen, H.; Schmülling, T. Cytokinin-deficient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity. Plant Cell 2003, 15, 2532–2550. [Google Scholar] [CrossRef] [Green Version]
- Brenner, W.G.; Schmülling, T. Transcript profiling of cytokinin action in Arabidopsis roots and shoots discovers largely similar but also organ-specific responses. BMC Plant Biol. 2012, 12, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmülling, T. Cytokinin regulates the activity of reproductive meristems, flower organ size, ovule formation, and thus seed yield in Arabidopsis thaliana. Plant Cell 2011, 23, 69–80. [Google Scholar] [CrossRef] [Green Version]
- D’Aloia, M.; Bonhomme, D.; Bouché, F.; Tamseddak, K.; Ormenese, S.; Torti, S.; Coupland, G.; Périlleux, C. Cytokinin promotes flowering of Arabidopsis via transcriptional activation of the FT paralogue TSF. Plant J. 2011, 65, 972–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takei, K.; Ueda, N.; Aoki, K.; Kuromori, T.; Hirayama, T.; Shinozaki, K.; Yamaya, T.; Sakakibara, H. AtIPT3 is a key determinant of nitrate-dependent cytokinin biosynthesis in Arabidopsis. Plant Cell Physiol. 2004, 45, 1053–1062. [Google Scholar] [CrossRef] [Green Version]
- Shtratnikova, V.Y.; Kudryakova, N.V.; Kudoyarova, G.R.; Korobova, A.V.; Akhiyarova, G.R.; Danilova, M.N.; Kusnetsov, V.V.; Kulaeva, O.N. Effects of nitrate and ammonium on growth of Arabidopsis thaliana plants transformed with the ARR5::GUS construct and a role for cytokinins in suppression of disturbances induced by the presence of ammonium. Russ. J. Plant Physiol. 2015, 62, 741–752. [Google Scholar] [CrossRef]
- Gu, J.; Li, Z.; Mao, Y.; Struik, P.C.; Zhang, H.; Liu, L.; Wang, Z.; Yang, J. Roles of nitrogen and cytokinin signals in root and shoot communications in maximizing of plant productivity and their agronomic applications. Plant Sci. 2018, 274, 320–331. [Google Scholar] [CrossRef]
- Gan, S.; Amasino, R.M. Cytokinins in plant senescence: From spray and pray to clone and play. Bioessays 1996, 18, 557–565. [Google Scholar] [CrossRef]
- Zwack, P.J.; Rashotte, A.M. Cytokinin inhibition of leaf senescence. Plant Signal. Behav. 2013, 8, e24737. [Google Scholar] [CrossRef]
- Cortleven, A.; Leuendorf, J.E.; Frank, M.; Pezzetta, D.; Bolt, S.; Schmülling, T. Cytokinin action in response to abiotic and biotic stresses in plants. Plant Cell Environ. 2019, 42, 998–1018. [Google Scholar] [CrossRef]
- Schwarz, I.; Scheirlinck, M.T.; Otto, E.; Bartrina, I.; Schmidt, R.C.; Schmülling, T. Cytokinin regulates the activity of the inflorescence meristem and components of seed yield in oilseed rape. J. Exp. Bot. 2020, 71, 7146–7159. [Google Scholar] [CrossRef]
- Riefler, M.; Novak, O.; Strnad, M.; Schmülling, T. Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. Plant Cell 2006, 18, 40–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miransari, M.; Smith, D.L. Plant hormones and seed germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Jameson, P.E.; Song, J. Cytokinin: A key driver of seed yield. J. Exp. Bot. 2016, 67, 593–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jameson, P.E.; Song, J. Will cytokinins underpin the second ‘Green Revolution’? J. Exp. Bot. 2020, 71, 6872–6875. [Google Scholar] [CrossRef]
- Wang, Z.; Cao, W.; Dai, T.; Zhou, Q. Effects of exogenous hormones on floret development and grain setting in wheat. Plant Growth Regul. 2001, 35, 225–231. [Google Scholar] [CrossRef]
- Gupta, N.K.; Gupta, S.; Shukla, D.S.; Deshmukh, P.S. Differential responses of BA injection on yield and specific grain growth in contrasting genotypes of wheat (Triticum aestivum L.). Plant Growth Regul. 2003, 40, 201–205. [Google Scholar] [CrossRef]
- Williams, R.H.; Cartwright, P.M. The effect of applications of a synthetic cytokinin on shoot dominance and grain yield in spring barley. Ann. Bot. 1980, 46, 445–452. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Poustini, K.; Ahmadi, A. Effects of foliar application of BAP on source and sink strength in four six-rowed barley (Hordeum vulgare L.) cultivars. Plant Growth Regul. 2008, 54, 231–239. [Google Scholar] [CrossRef]
- Warrier, A.; Bhardwaj, S.N.; Pande, P.C. Effect of benzyladenine on grain growth in Aestivum wheat. Plant Cell Physiol. 1987, 28, 735–739. [Google Scholar] [CrossRef]
- Sivakumar, T.; Srivastava, G.C. Effects of benzyl adenine and abscisic acid on grain yield and yield components in Triticale and wheat. J. Agron. Crop. Sci. 2001, 186, 43–46. [Google Scholar] [CrossRef]
- Koprna, R.; De Diego, N.; Dundálková, L.; Spíchal, L. Use of cytokinins as agrochemicals. Bioorg. Med. Chem. 2016, 24, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Nagel, L.; Brewster, R.; Riedell, W.E.; Reese, R.N. Cytokinin regulation of flower and pod set in soybeans (Glycine max (L.) Merr.). Ann. Bot. 2001, 88, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhao, J.; Song, J.; Jameson, P.E. Cytokinin glucosyl transferases, key regulators of cytokinin homeostasis, have potential value for wheat improvement. Plant Biotechnol. J. 2021, 19, 878–896. [Google Scholar] [CrossRef] [PubMed]
- McGaw, B.A.; Horgan, R. Cytokinin oxidase from Zea mays kernels and Vinca rosea crown-gall tissue. Planta 1983, 159, 30–37. [Google Scholar] [CrossRef]
- Galuszka, P.; Frébort, I.; Šebela, M.; Peč, P. Degradation of cytokinins by cytokinin oxidases in plants. Plant Growth Regul. 2000, 32, 315–327. [Google Scholar] [CrossRef]
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Plant science: Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef]
- Gemrotová, M.; Kulkarni, M.G.; Stirk, W.A.; Strnad, M.; Van Staden, J.; Spíchal, L. Seedlings of medicinal plants treated with either a cytokinin antagonist (PI-55) or an inhibitor of cytokinin degradation (INCYDE) are protected against the negative effects of cadmium. Plant Growth Regul. 2013, 71, 137–145. [Google Scholar] [CrossRef]
- Nisler, J.; Kopečný, D.; Končitíková, R.; Zatloukal, M.; Bazgier, V.; Berka, K.; Zalabák, D.; Briozzo, P.; Strnad, M.; Spíchal, L. Novel thidiazuron-derived inhibitors of cytokinin oxidase/dehydrogenase. Plant Mol. Biol. 2016, 92, 235–248. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Wardle, D.A.; Zurowski, C.; Looney, N.E. Phenylureas CPPU and thidiazuron affect yield components, fruit composition, and storage potential of four seedless grape selections. J. Am. Soc. Hortic. Sci. 1992, 117, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Nisler, J.; Zatloukal, M.; Spíchal, L.; Koprna, R.; Doležal, K.; Strnad, M. 1,2,3-thiadiazol-5yl-urea Derivatives, Use Thereof for Regulating Plant Senescence and Preparations Containing These Derivatives. U.S. Patent 9,993,002, 12 June 2018. [Google Scholar]
- Van Voorthuizen, M.J.; Nisler, J.; Song, J.; Spíchal, L.; Jameson, P.E. Targeting cytokinin homeostasis in rapid cycling Brassica rapa with plant growth regulators INCYDE and TD-K. Plants 2021, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Chatfield, J.M.; Armstrong, D.J. Regulation of cytokinin oxidase activity in callus tissues of Phaseolus vulgaris L. cv Great Northern. Plant Physiol. 1986, 80, 493–499. [Google Scholar] [CrossRef] [Green Version]
- Hare, P.D.; Van Staden, J. Inhibitory effect of thidiazuron on the activity of cytokinin oxidase isolated from soybean callus. Plant Cell Physiol. 1994, 35, 1121–1125. [Google Scholar] [CrossRef]
- Kopečný, D.; Briozzo, P.; Popelková, H.; Šebela, M.; Končitíková, R.; Spíchal, L.; Nisler, J.; Madzak, C.; Frébort, I.; Laloue, M.; et al. Phenyl-and benzylurea cytokinins as competitive inhibitors of cytokinin oxidase/dehydrogenase: A structural study. Biochimie 2010, 92, 1052–1062. [Google Scholar] [CrossRef]
- Yamada, H.; Suzuki, T.; Terada, K.; Takei, K.; Ishikawa, K.; Miwa, K.; Yamashino, T.; Mizuno, T. The Arabidopsis AHK4 histidine kinase is a cytokinin-binding receptor that transduces cytokinin signals across the membrane. Plant Cell Physiol. 2001, 42, 1017–1023. [Google Scholar] [CrossRef]
- Spíchal, L.; Rakova, N.Y.; Riefler, M.; Mizuno, T.; Romanov, G.A.; Strnad, M.; Schmülling, T. Two cytokinin receptors of Arabidopsis thaliana, CRE1/AHK4 and AHK3, differ in their ligand specificity in a bacterial assay. Plant Cell Physiol. 2004, 45, 1299–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatloukal, M.; Gemrotová, M.; Doležal, K.; Havlíček, L.; Spíchal, L.; Strnad, M. Novel potent inhibitors of A. thaliana cytokinin oxidase/dehydrogenase. Bioorg. Med. Chem. 2008, 16, 9268–9275. [Google Scholar] [CrossRef]
- Mok, M.C.; Mok, D.W.S.; Armstrong, D.J.; Shudo, K.; Isogai, Y.; Okamoto, T. Cytokinin activity of N-phenyl N’-1,2,3-thiadiazol-5-ylurea (Thidiazuron). Phytochemistry 1982, 21, 1509–1511. [Google Scholar] [CrossRef]
- Ferrante, A.; Hunter, D.A.; Hackett, W.P.; Reid, M.S. Thidiazuron—A potent inhibitor of leaf senescence in Alstroemeria. Postharvest Biol. Technol. 2002, 25, 333–338. [Google Scholar] [CrossRef]
- Malik, K.A.; Saxena, P.K. Regeneration in Phaseolus vulgaris L.: High-frequency induction of direct shoot formation in intact seedlings by N6-benzylaminopurine and thidiazuron. Planta 1992, 186, 384–389. [Google Scholar] [CrossRef]
- Singh, N.D.; Sahoo, L.; Sarin, N.B.; Jaiwal, P.K. The effect of TDZ on organogenesis and somatic embryogenesis in pigeonpea (Cajanus cajan L. Millsp). Plant Sci. 2003, 164, 341–347. [Google Scholar] [CrossRef]
- Zhihui, S.; Tzitzikas, M.; Raemakers, K.; Zhengqiang, M.; Visser, R. Effect of TDZ on plant regeneration from mature seeds in pea (Pisum sativum). Vitr. Cell. Dev. Biol. Plant 2009, 45, 776. [Google Scholar] [CrossRef]
- Guo, B.; Abbasi, B.H.; Zeb, A.; Xu, L.L.; Wei, Y.H. Thidiazuron: A multi-dimensional plant growth regulator. Afr. J. Biotechnol. 2011, 10, 8984–9000. [Google Scholar]
- Piao, Y.L.; Wang, H.T.; Zhang, H.M. Effects of TDZ on grapes quality in cold area. J. Agric. Sci. Yanbian Univ. 2006, 4, 012. [Google Scholar]
- Suttle, J.C. Involvement of ethylene in the action of the cotton defoliant thidiazuron. Plant Physiol. 1985, 78, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Hayata, Y.; Niimi, Y.; Iwasaki, N. Synthetic cytokinin-1-(2chloro4pyridyl)-3-phenylurea (CPPU)-promotes fruit set and induces parthenocarpy in watermelon. J. Am. Soc. Hort. Sci. 1995, 120, 997–1000. [Google Scholar] [CrossRef] [Green Version]
- Bilyeu, K.D.; Cole, J.L.; Laskey, J.G.; Riekhof, W.R.; Esparza, T.J.; Kramer, M.D.; Morris, R.O. Molecular and biochemical characterization of a cytokinin oxidase from maize. Plant Physiol. 2001, 125, 378–386. [Google Scholar] [CrossRef] [Green Version]
- Nisler, J.; Kopečný, D.; Pěkná, Z.; Končitíková, R.; Koprna, R.; Murvanidze, N.; Werbrouck, S.P.O.; Havlíček, L.; De Diego, N.; Kopečná, M.; et al. Diphenylurea-derived cytokinin oxidase/dehydrogenase inhibitors for biotechnology and agriculture. J. Exp. Bot. 2021, 72, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Piao, Y.L.; Lv, H.; Bai, H.J.; Li, M.D. Effect of TDZ on fruiting and fruit growth of Xiaoxiangshui pear of Yanbian [J]. J. Agric. Sci. Yanbian Univ. 2009, 3, 007. [Google Scholar]
- Guo, D.P.; Zhu, Z.J.; Hu, X.X.; Zheng, S.J. Effect of cytokinins on shoot regeneration from cotyledon and leaf segment of stem mustard (Brassica juncea var. tsatsai). Plant Cell Tissue Organ. Cult. 2005, 83, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Biasi, R.; Costa, G.; Giuliani, R.; Succi, F.; Sansavini, S. Effects of CPPU on kiwifruit performance. In II International Symposium on Kiwifruit; International Society for Horticultural Science: Korbeek-Lo, Belgium, 1991; Volume 297, pp. 367–374. [Google Scholar]
- Flaishman, M.A.; Shargal, A.; Shlizerman, L.; Stern, R.A. The synthetic cytokinins CPPU and TDZ prolong the phase of cell division in developing pear (Pyrus communis L.) fruit. Acta Hortic. 2006, 671, 151–157. [Google Scholar] [CrossRef]
- Stern, R.A.; Ben-Arie, R.; Neria, O.; Flaishman, M. CPPU and BA increase fruit size of ‘Royal Gala’ (Malus domestica) apple in a warm climate. J. Hort. Sci. Biotechnol. 2003, 78, 297–302. [Google Scholar] [CrossRef]
- Ferrara, G.; Mazzeo, A.; Netti, G.; Pacucci, C.; Matarrese, A.M.S.; Cafagna, I.; Mastrorilli, P.; Vezzoso, M.; Gallo, V. Girdling, gibberellic acid, and forchlorfenuron: Effects on yield, quality, and metabolic profile of table grape cv Italia. Am. J. Enol. Vitic. 2014, 65, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, N.; Oba, H.; Mizuta, D.; Yoshida, H.; Olsen, J.E. Timing of blue and red light exposure and CPPU application during the raising of seedlings can control flowering timing of petunia. Acta. Hortic. 2016, 1134, 171–178. [Google Scholar] [CrossRef]
- Jianchang, H.; Yan, X.; Chunxiang, Z.; Hongbin, L. Protective effect of CPPU in papaya plant under drought stress. J. Fruit Sci. 2003, 3, 013. [Google Scholar]
- Yip, W.K.; Yang, S.F. Effect of thidiazuron, a cytokinin-active urea derivative, in cytokinin-dependent ethylene production systems. Plant Physiol. 1986, 80, 515–519. [Google Scholar] [CrossRef] [Green Version]
- Zatloukal, M.; Plihalova, L.; Klaskova, J.; Spíchal, L.; Koprna, R.; Dolezal, K.; Strnad, M. Substituted 6-Anilino-9-heterocyclylpurine Derivatives for Inhibition of Plant Stress. U.S. Patent 10,662,194, 26 May 2020. [Google Scholar]
- Aremu, A.O.; Masondo, N.A.; Sunmonu, T.O.; Kulkarni, M.G.; Zatloukal, M.; Spichal, L.; Doležal, K.; Van Staden, J. A novel inhibitor of cytokinin degradation (INCYDE) influences the biochemical parameters and photosynthetic apparatus in NaCl-stressed tomato plants. Planta 2014, 240, 877–889. [Google Scholar] [CrossRef]
- Aremu, A.O.; Stirk, W.A.; Masondo, N.A.; Plačková, L.; Novák, O.; Pěnčík, A.; Zatloukal, M.; Nisler, J.; Spíchal, L.; Doležal, K.; et al. Dissecting the role of two cytokinin analogues (INCYDE and PI-55) on in vitro organogenesis, phytohormone accumulation, phytochemical content and antioxidant activity. Plant Sci. 2015, 238, 81–94. [Google Scholar] [CrossRef]
- Reusche, M.; Klásková, J.; Thole, K.; Truskina, J.; Novák, O.; Janz, D.; Strnad, M.; Spíchal, L.; Lipka, V.; Teichmann, T. Stabilization of cytokinin levels enhances Arabidopsis resistance against Verticillium longisporum. Am. Phytopath. Soc. 2013, 26, 850–860. [Google Scholar] [CrossRef] [Green Version]
- Koprna, R.; Humplík, J.F.; Špíšek, Z.; Bryksová, M.; Zatloukal, M.; Mik, V.; Novák, O.; Nisler, J.; Doležal, K. Improvement of tillering and grain yield by application of cytokinin derivatives in wheat and barley. Agronomy 2021, 11, 67. [Google Scholar] [CrossRef]
- Van Voorthuizen, M.J. Novel plant growth regulator effects on yield, senescence and cytokinin homeostasis in wheat, barley and rapid cycling Brassica rapa. Ph.D. Thesis, University of Canterbury, Christchurch, New Zealand, 2018. [Google Scholar]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. Konzak: “A decimal code for the growth stages of cereals”. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Berková, V.; Kameniarová, M.; Ondrisková, V.; Berka, M.; Menšíková, S.; Kopecká, R.; Luklová, M.; Novák, J.; Spíchal, L.; Rashotte, A.M.; et al. Arabidopsis response to inhibitor of cytokinin degradation INCYDE: Modulations of cytokinin signaling and plant proteome. Plants 2020, 9, 1563. [Google Scholar] [CrossRef] [PubMed]
- Aremu, A.O.; Bairu, M.W.; Novák, O.; Plačková, L.; Zatloukal, M.; Doležal, K.; Finnie, J.F.; Strnad, M.; Van Staden, J. Physiological responses and endogenous cytokinin profiles of tissue-cultured “Williams” bananas in relation to roscovitine and an inhibitor of cytokinin oxidase/dehydrogenase (INCYDE) treatments. Planta 2012, 236, 1775–1790. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.J.; Wang, J.Q.; Huang, Z.L.; Mi, L.; Xu, K.F.; Wu, J.J.; Fan, Y.H.; Ma, S.Y.; Jiang, D.G. Effects of low temperature at booting stage on sucrose metabolism and endogenous hormone contents in winter wheat spikelet. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Erena, M.F.; Lohraseb, I.; Munoz-Santa, I.; Taylor, J.D.; Emebiri, L.C.; Collins, N.C. The WtmsDW locus on wheat chromosome 2B controls major natural variation for floret sterility responses to heat stress at booting stage. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Jameson, P.E.; McWha, J.A.; Wright, G.J. Cytokinins and changes in their activity during the development of grains of wheat (Triticum aestivum L.). Zeit. Pflanzenphysiol. 1982, 106, 27–36. [Google Scholar] [CrossRef]
- Lenton, J.R.; Appleford, N.E.J. Cytokinins and Early Growth in Wheat; Monograph, British Plant Growth Regulator Group: Wantage, UK, 1986; Volume 14, pp. 99–113. [Google Scholar]
- Banowetz, G.M.; Ammar, K.; Chen, D.D. Postanthesis temperatures influence cytokinin accumulation and wheat kernel weight. Plant Cell Environ. 1999, 22, 309–316. [Google Scholar] [CrossRef]
- Hess, J.R.; Carman, J.G.; Banowetz, G.M. Hormones in wheat kernels during embryony. J. Plant Physiol. 2002, 159, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.N.; Perry, L.; Kisiala, A.; Olechowski, H.; Emery, R.N. Cytokinin activity during early kernel development corresponds positively with yield potential and later stage ABA accumulation in field-grown wheat (Triticum aestivum L.). Planta 2020, 252, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Faix, B.; Radchuk, V.; Nerlich, A.; Hümmer, C.; Radchuk, R.; Emery, R.J.N.; Keller, H.; Götz, K.-P.; Weschke, W.; Geigenberger, P.; et al. Barley grains, deficient in cytosolic small subunit of ADP-glucose pyrophosphorylase, reveal coordinate adjustment of C: N metabolism mediated by an overlapping metabolic-hormonal control. Plant J. 2012, 69, 1077–1093. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.F.; Paleczny, A.R.; Olechowski, H.; Emery, R.N. Changes in cytokinin form and concentration in developing kernels correspond with variation in yield among field-grown barley cultivars. Plant Physiol. Biochem. 2013, 64, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Motyka, V.; Faiss, M.; Strnad, M.; Kamínek, M.; Schmulling, T. Changes in cytokinin content and cytokinin oxidase activity in response to derepression of ipt gene transcription in transgenic tobacco calli and plants. Plant Physiol. 1996, 112, 1035–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motyka, V.; Vaňková, R.; Čapková, V.; Petrášek, J.; Kamínek, M.; Schmülling, T. Cytokinin-induced upregulation of cytokinin oxidase activity in tobacco includes changes in enzyme glycosylation and secretion. Physiol. Plant 2003, 117, 11–21. [Google Scholar] [CrossRef]
- Brugière, N.; Jiao, S.; Hantke, S.; Zinselmeier, C.; Roessler, J.A.; Niu, X.; Jones, R.J.; Habben, J.E. Cytokinin oxidase gene expression in maize is localized to the vasculature, and is induced by cytokinins, abscisic acid, and abiotic stress. Plant Physiol. 2003, 132, 1228–1240. [Google Scholar] [CrossRef] [Green Version]
- Blagoeva, E.; Dobrev, P.I.; Malbeck, J.; Motyka, V.; Gaudinová, A.; Vaňková, R. Effect of exogenous cytokinins, auxins and adenine on cytokinin N-glucosylation and cytokinin oxidase/dehydrogenase activity in de-rooted radish seedlings. Plant Growth Regul. 2004, 44, 15–23. [Google Scholar] [CrossRef]
- Hirose, N.; Takei, K.; Kuroha, T.; Kamada-Nobusada, T.; Hayashi, H.; Sakakibara, H. Regulation of cytokinin biosynthesis, compartmentalization and translocation. J. Exp. Bot. 2008, 59, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Vyroubalová, Š.; Václavíková, K.; Turečková, V.; Novák, O.; Šmehilová, M.; Hluska, T.; Ohnoutková, L.; Frébort, I.; Galuszka, P. Characterization of new maize genes putatively involved in cytokinin metabolism and their expression during osmotic stress in relation to cytokinin levels. Plant Physiol. 2009, 151, 433–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasparis, S.; Przyborowski, M.; Kała, M.; Nadolska-Orczyk, A. Knockout of the HvCKX1 or HvCKX3 gene in barley (Hordeum vulgare L.) by RNA-Guided Cas9 nuclease affects the regulation of cytokinin metabolism and root morphology. Cells 2019, 8, 782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Bai, W.; Zeng, Q.; Song, S.; Zhang, M.; Li, X.; Hou, L.; Xiao, Y.; Luo, M.; Li, D.; et al. Moderately enhancing cytokinin level by down-regulation of GhCKX expression in cotton concurrently increases fiber and seed yield. Mol. Breed. 2015, 35, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Song, G.; Gao, J.; Zhang, S.; Zhang, R.; Li, W.; Chen, M.; Liu, M.; Xia, X.; Risacher, T.; et al. Enhancement of grain number per spike by RNA interference of cytokinin oxidase 2 gene in bread wheat. Hereditas 2018, 155, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalewski, W.; Galuszka, P.; Gasparis, S.; Orczyk, W.; Nadolska-Orczyk, A. Silencing of the HvCKX1 gene decreases the cytokinin oxidase/dehydrogenase level in barley and leads to higher plant productivity. J. Exp. Bot. 2010, 61, 1839–1851. [Google Scholar] [CrossRef] [Green Version]
- Zalewski, W.; Gasparis, S.; Boczkowska, M.; Rajchel, I.K.; Kała, M.; Orczyk, W.; Nadolska-Orczyk, A. Expression patterns of HvCKX genes indicate their role in growth and reproductive development of barley. PLoS ONE 2014, 9, e115729. [Google Scholar] [CrossRef] [Green Version]
- Yeh, S.Y.; Chen, H.W.; Ng, C.Y.; Lin, C.Y.; Tseng, T.H.; Li, W.H.; Ku, M.S. Down-regulation of cytokinin oxidase 2 expression increases tiller number and improves rice yield. Rice 2015, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Uauy, C.; Wulff, B.B.; Dubcovsky, J. Combining traditional mutagenesis with new high-throughput sequencing and genome editing to reveal hidden variation in polyploid wheat. Annu. Rev. Genet. 2017, 51, 435–454. [Google Scholar] [CrossRef] [Green Version]
- Gil, J.; Andrade-Martínez, J.S.; Duitama, J. Accurate, efficient and user-friendly mutation calling and sample identification for TILLING experiments. Front. Genet. 2021, 12, 624513. [Google Scholar] [CrossRef]
- Lassoued, R.; Phillips, P.W.; Macall, D.M.; Hesseln, H.; Smyth, S.J. Expert opinions on the regulation of plant genome editing. Plant Biotechnol. J. 2021, 19, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Wheat Tilling. Available online: https://wheat-tilling.com/ (accessed on 25 October 2021).
- Krasileva, K.V.; Vasquez-Gross, H.A.; Howell, T.; Bailey, P.; Paraiso, F.; Clissold, L.; Dubcovsky, J. Uncovering hidden variation in polyploid wheat. Proc. Natl. Acad. Sci. USA 2017, 114, E913–E921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Jiang, L.; Jameson, P.E. Co-ordinate regulation of cytokinin gene family members during flag leaf and reproductive development in wheat. BMC Plant Biol. 2012, 12, 78. [Google Scholar] [CrossRef] [Green Version]
- Svačinová, J.; Novák, O.; Plačková, L.; Lenobel, R.; Holík, J.; Strnad, M.; Doležal, K. A new approach for cytokinin isolation from Arabidopsis tissues using miniaturized purification: Pipette tip solid-phase extraction. Plant Methods 2012, 8, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Wheat Cultivar Orator (2013/14) | |||
| Treatment | Yield (T/ha) | TGW (g) | Protein (%) |
| Untreated Control | 10.8 ± 0.2 | 46.0 ± 1.1 | 11.2 ± 0.2 |
| DMSO 50 µM Control (GS 39, 51, 61, 65) | 11.1 ± 0.1 | 46.6 ± 0.2 | 11.2 ± 0.1 |
| DMSO 25 µM Control (GS 61, 65, 65 + 13 d) | 11.2 ± 0.1 | 47.2 ± 0.7 | 11.0 ± 0.1 |
| INCYDE 10 µM (GS 65) | 11.4 ± 0.2 | 45.9 ± 1.1 | 11.4 ± 0 |
| INCYDE 25 µM (GS 39, 51, 61, 65) | 11.3 ± 0.2 | 46.2 ± 0.7 | 11.1 ± 0.1 |
| INCYDE 25 µM (GS 39) | 11.2 ± 0.2 | 45.0 ± 1.1 | 11.2 ± 0.1 |
| INCYDE 25 µM (GS 51) | 11.0 ± 0.1 | 45.4 ± 0.8 | 11.1 ± 0.2 |
| INCYDE 25 µM (GS 61) | 11.1 ± 0.2 | 45.6 ± 0.4 | 11.1 ± 0.1 |
| INCYDE 25 µM (GS 65) | 11.1 ± 0.2 | 45.1 ± 0.5 | 11.2 ± 0.1 |
| INCYDE 50 µM (GS 61) | 11.1 ± 0.1 | 45.5 ± 1.2 | 11.4 ± 0.1 |
| INCYDE 50 µM (GS 65) | 11.1 ± 0.2 | 46.9 ± 0.3 | 11.4 ± 0.1 |
| TD-K 10 µM (GS 61, 65, 65 + 13 d) | 11.1 ± 0.1 | 46.3 ± 1.1 | 11.1 ± 0.1 |
| TD-K 25 µM (GS 61, 65, 65 + 13 d) | 11.4 ± 0.1 | 46.0 ± 0.5 | 11.1 ± 0.1 |
| Wheat Cultivar Torch (2014/15) | |||
| Treatment | Yield (T/ha) | TGW (g) | Protein (%) |
| Untreated Control | 14.5 ± 0.1 | 46.8 ± 1.1 | 9.8 ± 0.2 |
| DMSO Control (GS 51, 61, 65, 65 + 15 d) | 14.5 ± 0.3 | 48.1 ± 0.2 | 9.9 ± 0.2 |
| TD-K 10 µM (GS 61, 65, 65 + 15 d) | 14.5 ± 0.2 | 47.4 ± 0.5 | 10.0 ± 0.2 |
| TD-K 50 µM (GS 61, 65, 65 + 15 d) | 14.7 ± 0.3 | 49.1 ± 0.5 | 9.8 ± 0.05 |
| CPPU 10 µM (GS 61, 65) | 14.4 ± 0.3 | 47.1 ± 0.4 | 10.0 ± 0.2 |
| CPPU 30 µM (GS 61, 65) | 14.5 ± 0.3 | 48.8 ± 0.2 | 9.7 ± 0.1 |
| CPPU 100 µM (GS 61, 65) | 14.6 ± 0.2 | 48.4 ± 0.3 | 9.7 ± 0.3 |
| CPPU 10 µM (GS 51, 65) | 14.5 ± 0.2 | 47.8 ± 0.4 | 9.6 ± 0.1 |
| CPPU 30 µM (GS 51, 65) | 14.7 ± 0.1 | 47.2 ± 0.9 | 9.9 ± 0.1 |
| CPPU 100 µM (GS 51, 65) | 14.4 ± 0.2 | 48.1 ± 0.8 | 9.9 ± 0.2 |
| Barley Cultivar Quench (2014/15) | |||
| Treatment | Yield (T/ha) | TGW (g) | Protein (%) |
| Untreated Control | 10.8 ± 0.2 | 52.3 ± 0.5 | 14.3 ± 0.1 |
| DMSO Control (GS 51, 61, 65, 65 + 15 d) | 11.0 ± 0.1 | 52.0 ± 0.4 | 13.9 ± 0.4 |
| INCYDE 10 µM (GS 65) | 11.0 ± 0.2 | 53.0 ± 0.2 | 13.8 ± 0.3 |
| INCYDE 25 µM (GS 39, 51, 61, 65) | 11.2 ± 0.2 | 51.8 ± 0.5 | 14.0 ± 0.2 |
| INCYDE 25 µM (GS 39) | 11.3 ± 0.2 | 52.5 ± 0.5 | 13.8 ± 0.2 |
| INCYDE 25 µM (GS 51) | 11.3 ± 0.1 | 52.4 ± 0.9 | 14.0 ± 0.4 |
| INCYDE 25 µM (GS 61) | 11.1 ± 0.1 | 52.2 ± 0.9 | 13.9 ± 0.5 |
| INCYDE 25 µM (GS 65) | 11.3 ± 0.1 | 52.5 ± 0.7 | 14.2 ± 0.4 |
| INCYDE 50 µM (GS 61) | 11.2 ± 0.1 | 52.5 ± 0.8 | 14.1 ± 0.4 |
| INCYDE 50 µM (GS 65) | 11.2 ± 0.3 | 52.3 ± 0.4 | 13.9 ± 0.5 |
| Wheat Cytokinin Concentrations (pmol/g DW) | |||
|---|---|---|---|
| Type | Control | TD-K 50 µM | CPPU 100 µM |
| tZ | 794.5 ± 71.1 | 705.3 ± 75.7 | 889.2 ± 73.7 |
| tZR | 60.0 ± 6.7 | 60.3 ± 3.2 | 63.5 ± 2.3 |
| tZOG | 20.4 ± 2.8 | 24.7 ± 0.5 | 22.8 ± 1.3 |
| tZROG | 5.8 ± 0.7 | 6.8 ± 0.2 | 6.5 ± 0.5 |
| tZRMP | 115.9 ± 3.6 | 106.5 ± 2.8 | 103.1 ± 10.5 |
| tZ7G | <LOD | <LOD | <LOD |
| tZ9G | 247.3 ± 21.3 | 286.1 ± 3.5 | 268.7 ± 11.5 |
| Total tZ types | 1244.0 ± 104.8 | 1189.7 ± 85.2 | 1353.8 ± 57.0 |
| iP | 1.6 ± 0.2 | 1.3 ± 0.1 | 1.3 ± 0.1 |
| iPR | 2.2 ± 0.2 | 1.8 ± 0.1 | 2.5 ± 0.4 |
| iPRMP | 22.9 ± 1.5 | 22.1 ± 3.8 | 25.9 ± 4.0 |
| iP7G | <LOD | <LOD | <LOD |
| iP9G | <LOD | <LOD | <LOD |
| Total iP types | 26.8 ± 1.7 | 25.1 ± 3.9 | 29.7 ± 4.1 |
| cZ | 9.6 ± 0.5 | 7.8 ± 1.1 | 8.6 ± 1.2 |
| cZR | 33.9 ± 3.9 | 24.7 ± 3.5 | 30.5 ± 4.0 |
| cZOG | 114.3 ± 12.4 | 130.8 ± 11.7 | 116.1 ± 12.9 |
| cZROG | 139.1 ± 13.2 | 154.5 ± 6.6 | 150.6 ± 8.9 |
| cZRMP | 10.0 ± 0.8 | 7.7 ± 1.3 | 10.8 ± 1.2 |
| cZ9G | <LOD | <LOD | <LOD |
| Total cZ types | 306.5 ± 26.5 | 325.5 ± 12.2 | 316.5 ± 17.0 |
| DHZ | 0.23 ± 0.01 | 0.20 ± 0.03 | 0.20 ± 0.03 |
| DHZR | 2.9 ± 0.1 | 2.4 ± 0.3 | 2.6 ± 0.3 |
| DHZOG | 1.4 ± 0.2 | 1.7 ± 0.1 | 1.5 ± 0.1 |
| DHZROG | 9.5 ± 1.1 | 10.7 ± 0.6 | 10.0 ± 0.8 |
| DHZRMP | <LOD | <LOD | <LOD |
| DHZ7G | 15.0 ± 0.3 | 13.3 ± 0.9 | 14.6 ± 2.2 |
| DHZ9G | 0.07 ± 0.003 | 0.06 ± 0.01 | 0.07 ± 0.01 |
| Total DHZ types | 29.1 ± 1.5 | 28.2 ± 0.7 | 28.9 ± 2.6 |
| Total CK bases | 806.0 ± 71.4 | 714.6 ± 76.9 | 899.3 ± 74.9 |
| Total CK ribosides | 98.7 ± 6.8 | 89.1 ± 7.0 | 99.0 ± 7.0 |
| Total CK nucleotides | 148.9 ± 3.7 | 136.3 ± 2.0 | 139.8 ± 12.8 |
| Total CK O-glucosides | 290.5 ± 30.2 | 329.2 ± 18.8 | 307.5 ± 23.7 |
| Total CK N-glucosides | 262.3 ± 21.6 | 299.4 ± 3.9 | 283.4 ± 13.6 |
| Total cytokinins | 1606.4 ± 117.3 | 1568.5 ± 69.5 | 1728.9 ± 43.4 |
| Barley Cytokinin Concentrations (pmol/g DW) | ||
|---|---|---|
| Type | Control | INCYDE 50 µM |
| tZ | 759.5 ± 66.8 | 642.8 ± 30.0 |
| tZR | 432.0 ± 44.3 | 458.9 ± 36.6 |
| tZOG | 89.5 ± 2.7 | 97.6 ± 7.9 |
| tZROG | 28.1 ± 1.8 | 30.2 ± 0.6 |
| tZRMP | 455.3 ± 27.0 | 454.6 ± 13.0 |
| tZ7G | <LOD | <LOD |
| tZ9G | 46.2 ± 2.6 | 65.0 ± 9.1 |
| Total tZ types | 1810.6 ± 142.1 | 1749.1 ± 61.6 |
| iP | 2.0 ± 0.2 | 2.3 ± 0.3 |
| iPR | 4.5 ± 0.6 | 4.7 ± 0.3 |
| iPRMP | 86.8 ± 12.8 | 89.0 ± 11.9 |
| iP7G | <LOD | <LOD |
| iP9G | <LOD | <LOD |
| Total iP types | 93.3 ± 13.6 | 96.0 ± 12.2 |
| cZ | 3.6 ± 0.1 | 3.2 ± 0.1 |
| cZR | 23.4 ± 0.5 | 20.1 ± 0.5 * |
| cZOG | 328.9 ± 11.8 | 417.1 ± 9.7 * |
| cZROG | 242.5 ± 8.4 | 256.8 ± 8.6 |
| cZRMP | 20.0 ± 1.8 | 20.6 ± 1.9 |
| cZ9G | <LOD | <LOD |
| Total cZ types | 618.4 ± 18.8 | 717.9 ± 12.8 * |
| DHZ | 0.9 ± 0.02 | 0.7 ± 0.04 * |
| DHZR | 6.5 ± 0.1 | 5.1 ± 0.3 * |
| DHZOG | 12.6 ± 0.6 | 10.6 ± 0.4 * |
| DHZROG | 47.3 ± 3.1 | 43.0 ± 2.8 |
| DHZRMP | <LOD | <LOD |
| DHZ7G | 6.0 ± 0.3 | 5.0 ± 0.7 |
| DHZ9G | 0.06 ± 0.01 | 0.06 ± 0.01 |
| Total DHZ types | 73.0 ± 3.7 | 64.5 ± 3.8 |
| Total CK bases | 766.0 ± 67.0 | 649.0 ± 30.4 |
| Total CK ribosides | 466.3 ± 44.6 | 488.8 ± 37.1 |
| Total CK nucleotides | 562.2 ± 38.6 | 564.2 ± 11.1 |
| Total CK O-glucosides | 748.9 ± 27.8 | 855.4 ± 25.2 * |
| Total CK N-glucosides | 51.9 ± 2.6 | 70.1 ± 8.7 |
| Total cytokinins | 2595.2 ± 177.1 | 2627.5 ± 68.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Voorthuizen, M.J.; Song, J.; Novák, O.; Jameson, P.E. Plant Growth Regulators INCYDE and TD-K Underperform in Cereal Field Trials. Plants 2021, 10, 2309. https://doi.org/10.3390/plants10112309
van Voorthuizen MJ, Song J, Novák O, Jameson PE. Plant Growth Regulators INCYDE and TD-K Underperform in Cereal Field Trials. Plants. 2021; 10(11):2309. https://doi.org/10.3390/plants10112309
Chicago/Turabian Stylevan Voorthuizen, Matthew J., Jiancheng Song, Ondřej Novák, and Paula E. Jameson. 2021. "Plant Growth Regulators INCYDE and TD-K Underperform in Cereal Field Trials" Plants 10, no. 11: 2309. https://doi.org/10.3390/plants10112309
APA Stylevan Voorthuizen, M. J., Song, J., Novák, O., & Jameson, P. E. (2021). Plant Growth Regulators INCYDE and TD-K Underperform in Cereal Field Trials. Plants, 10(11), 2309. https://doi.org/10.3390/plants10112309