
Cytotoxic Effect In Vitro of Acalypha monostachya Extracts over Human Tumor Cell Lines

 , ,
, ,  , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Taxonomic Identification of the Plant
2.2. Extract Yields and Phytochemical Screening
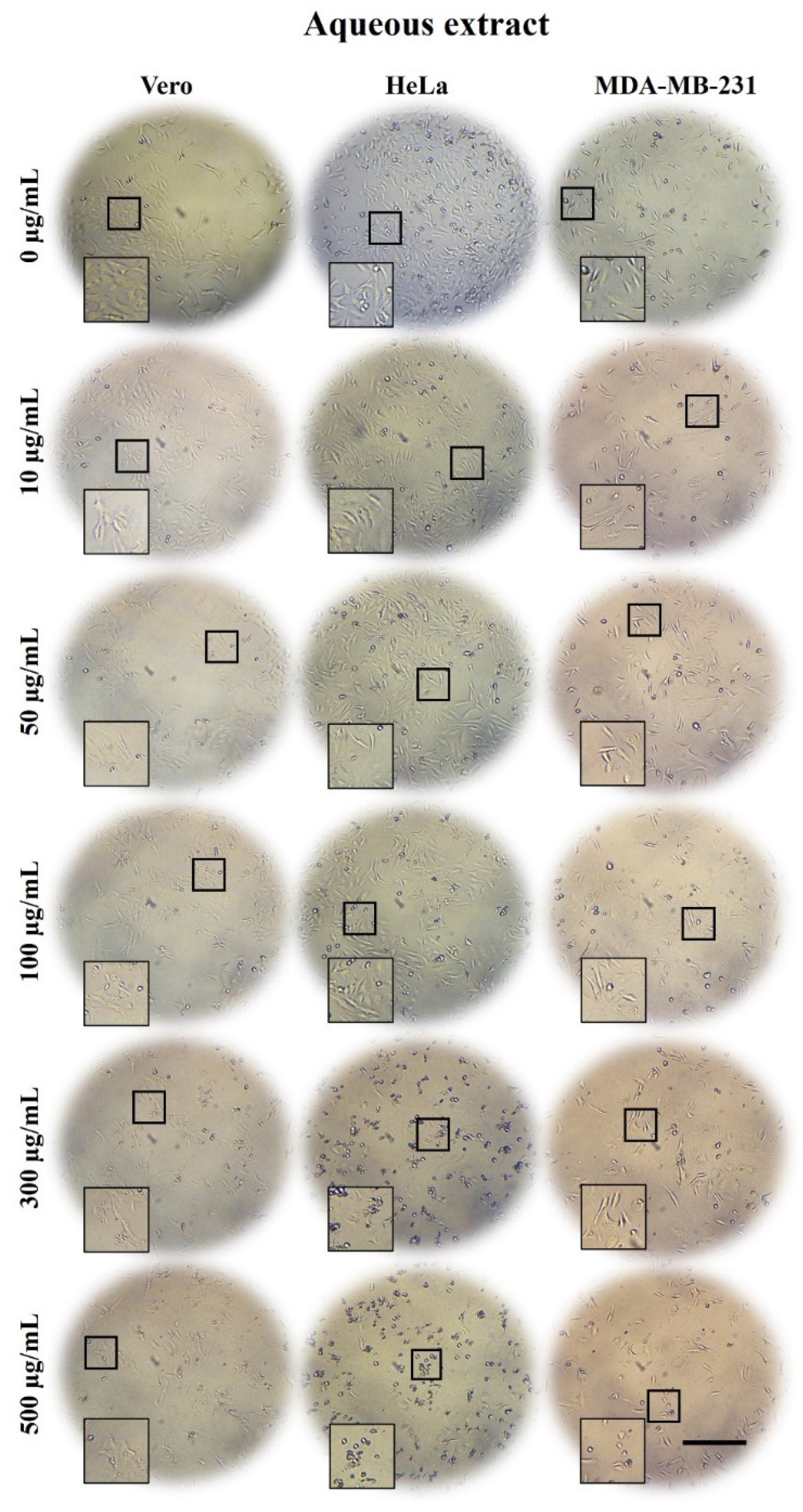
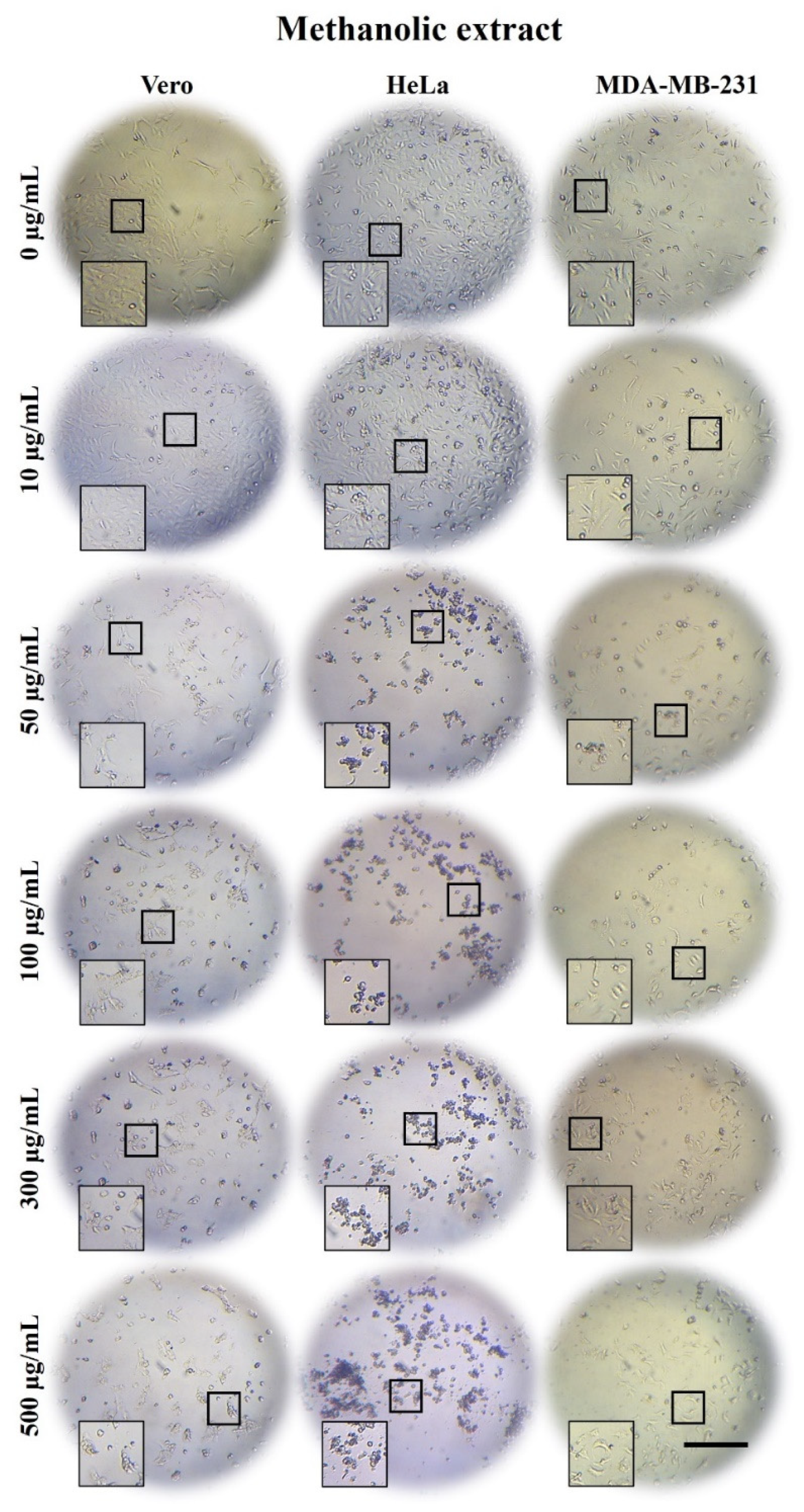
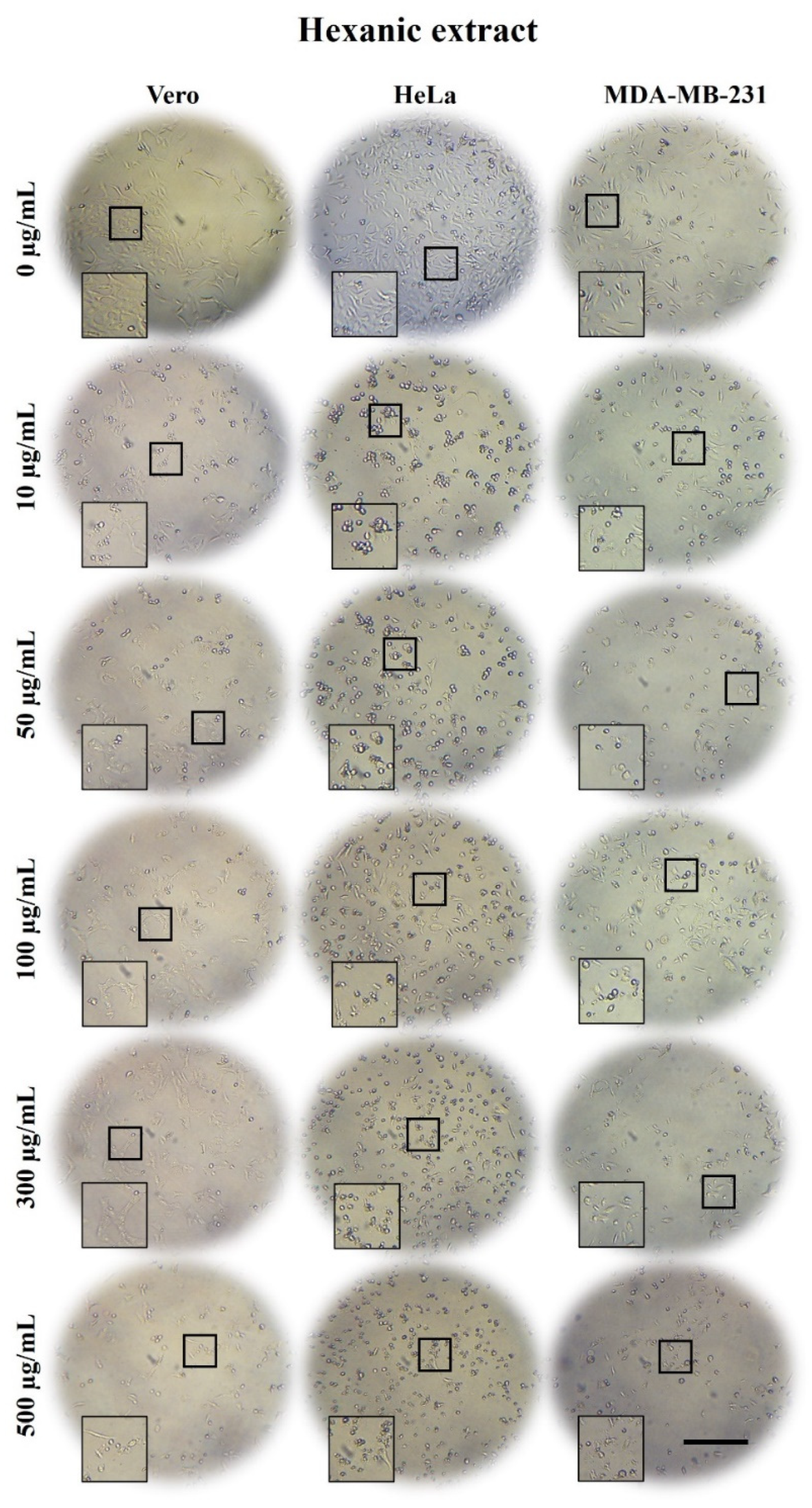
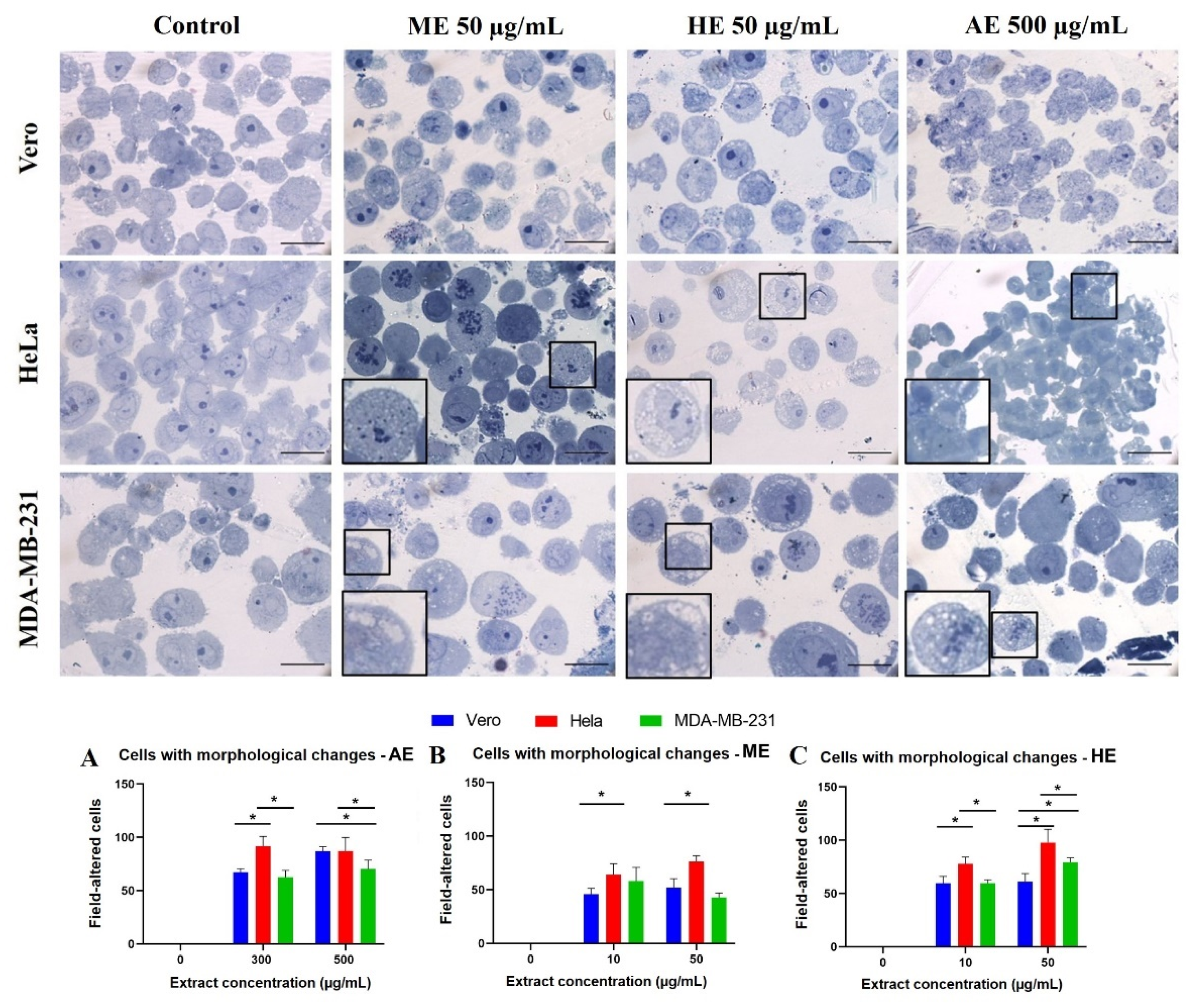
2.3. A. Monostachya Extracts Induce Morphological Changes in Cultured Human Tumor Cells
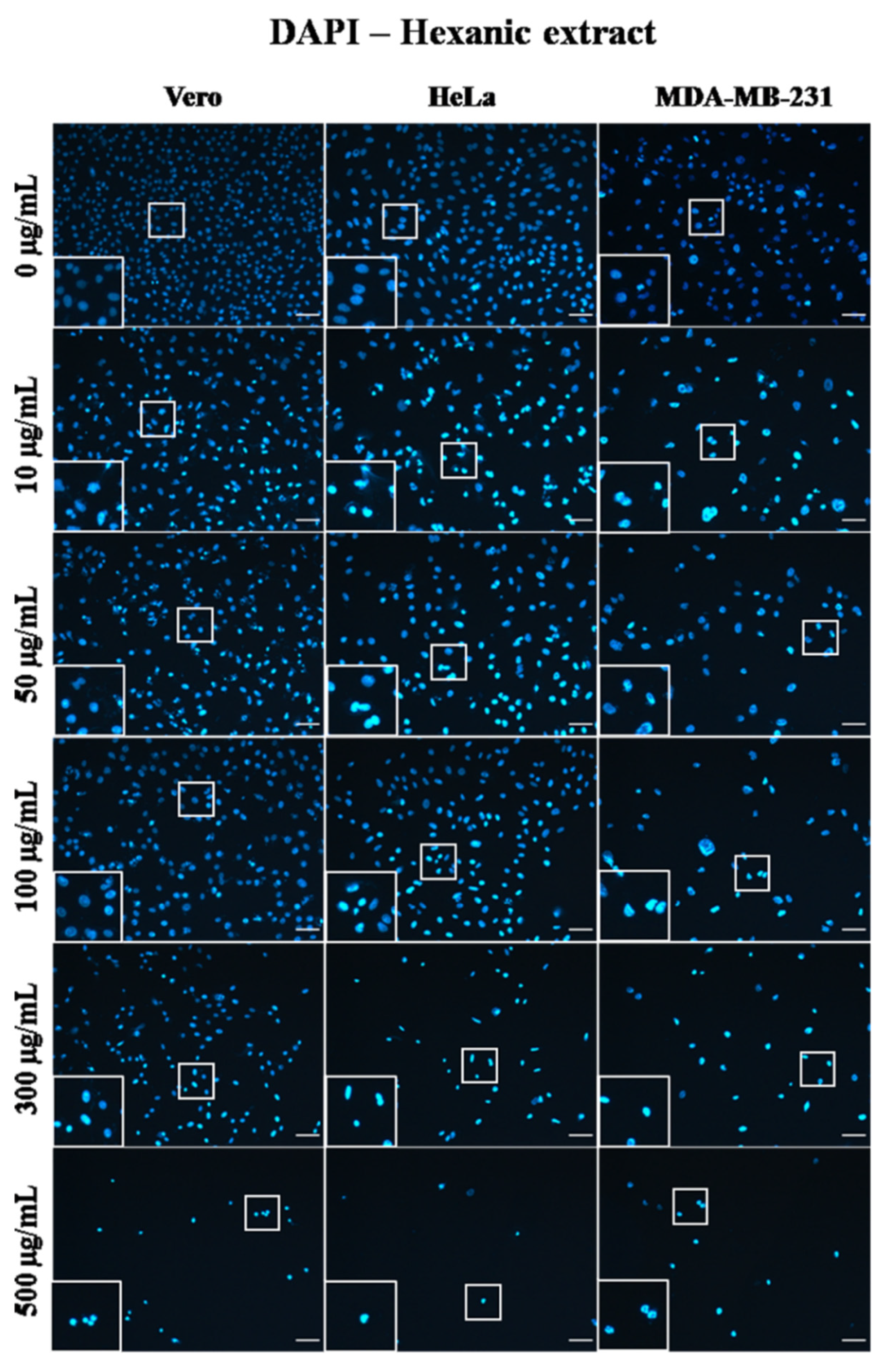
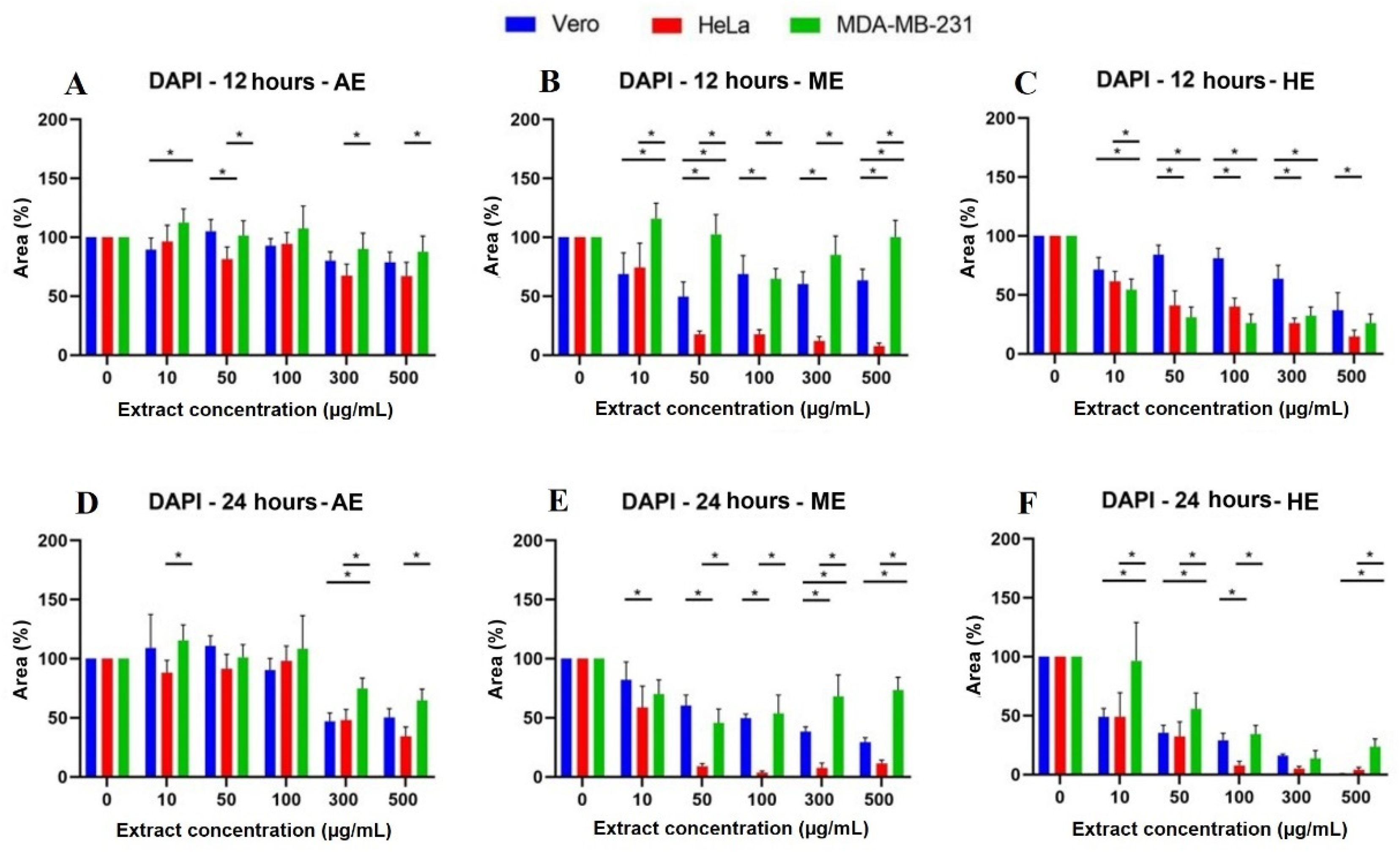
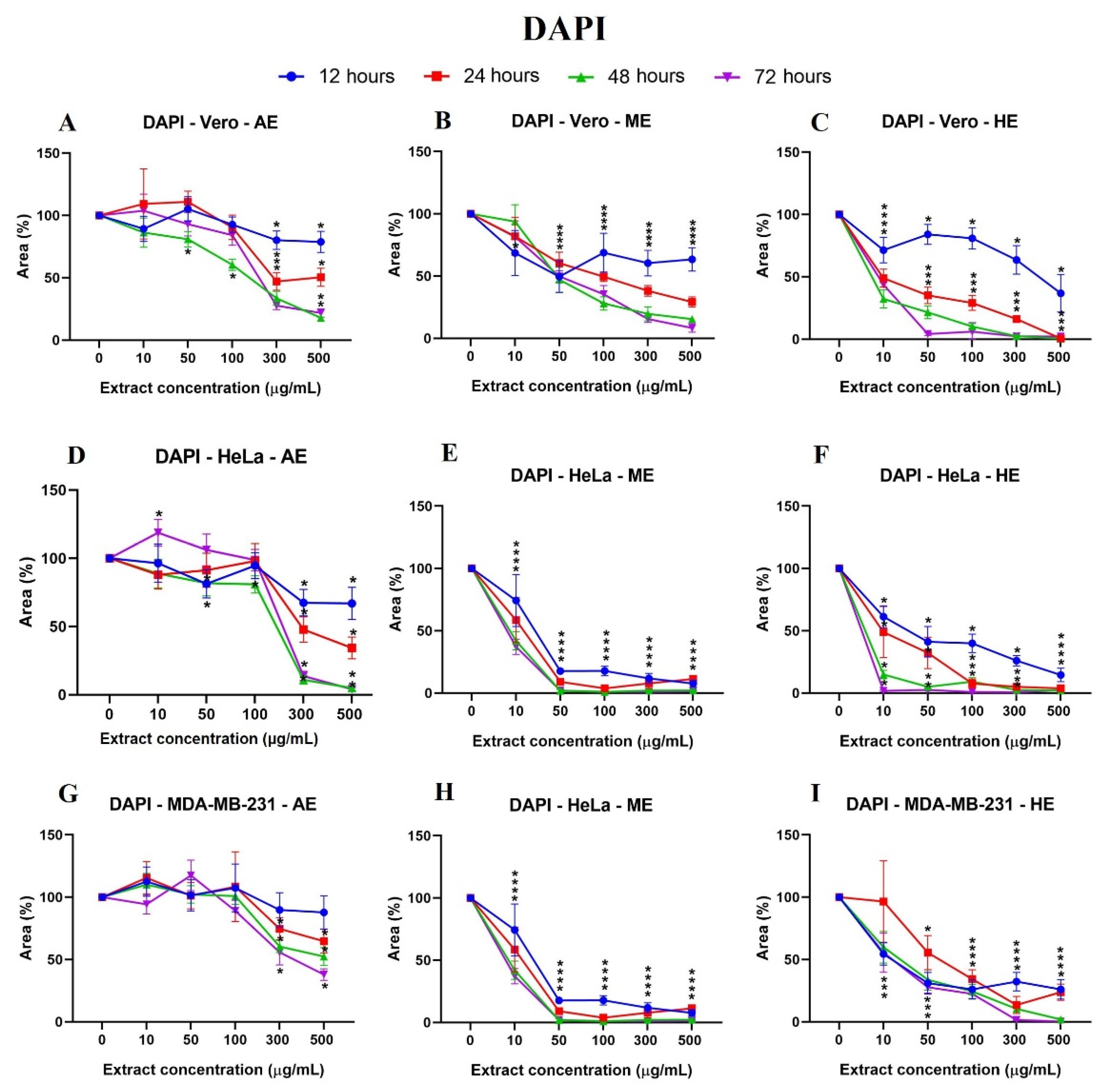
2.4. A. Monostachya Extracts Induce Changes in the Nuclear Morphology of Human Tumor Cells
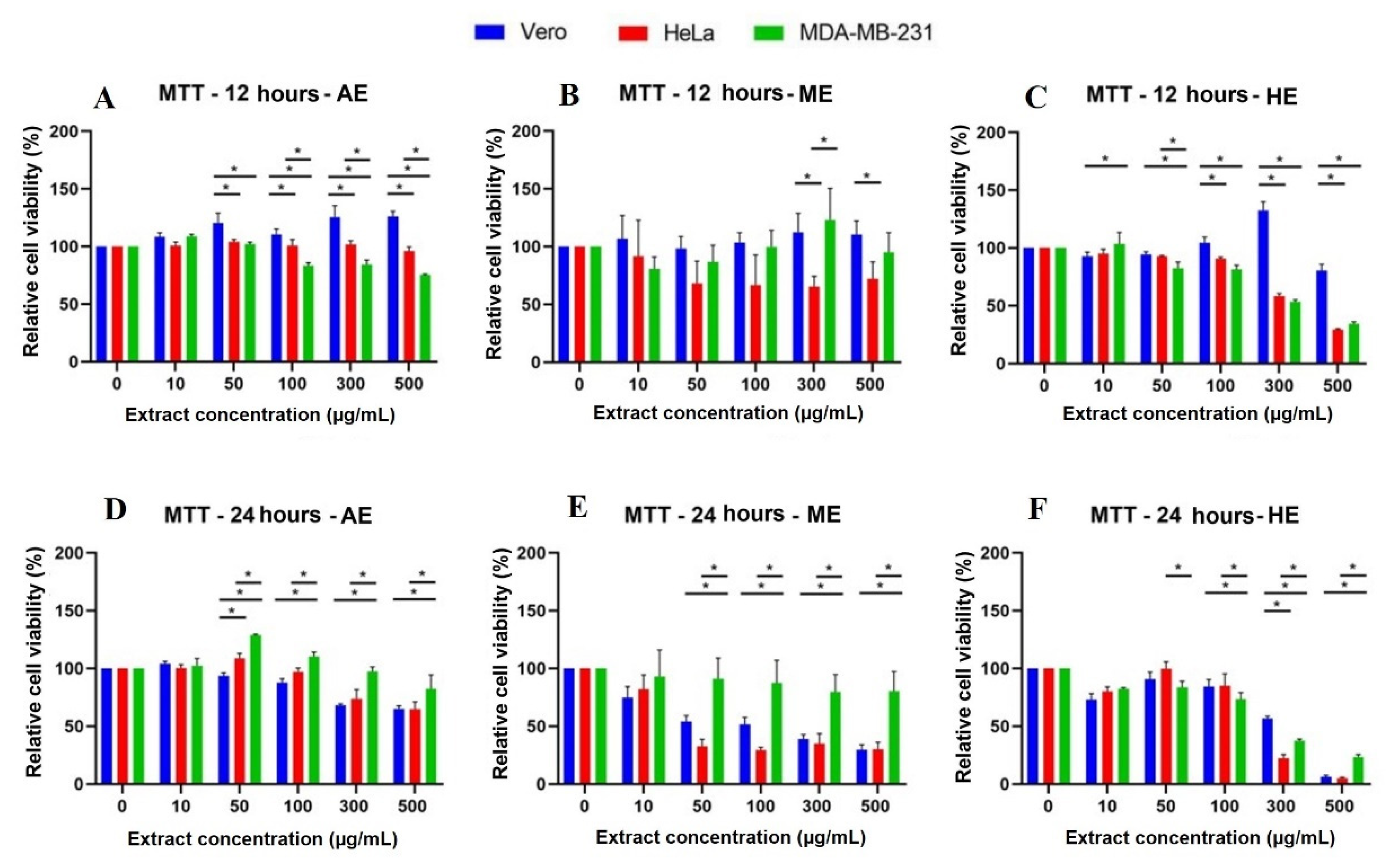
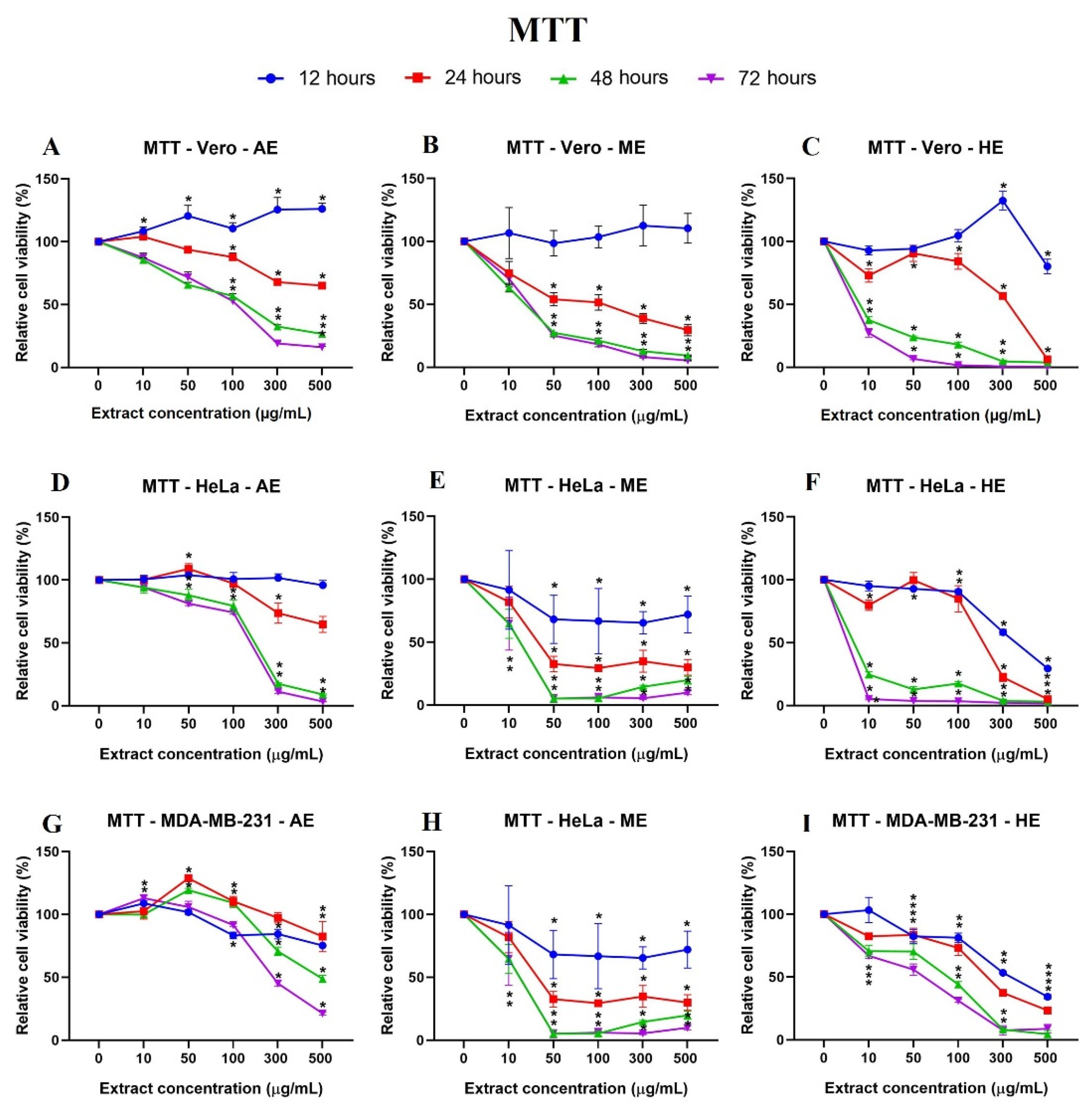
2.5. A. Monostachya Extracts Decrease the Viability in Human Tumor Cells
2.6. Human Tumor Cells Show More Morphological and Nuclear Alterations upon Exposure to A. Monostachya Extracts
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. A. Monostachya Extracts
4.3. Partial Characterization (Partial Phytochemical Screening)
4.3.1. Instaurations (KMnO4 Test)
4.3.2. Carbonyl Group (2-4 Dinitrophenylhydrazine Test)
4.3.3. Phenolic Compounds (Vegetable Tannins) (FeCl3 Test)
4.3.4. Steroids and Terpenes (Salkowski Test)
4.3.5. Carbohydrates
Molish Test
Coumarin Test
Lactone Test
4.3.6. Sesquiterpene Lactones (Baljet Test)
4.3.7. Flavonoids (H2SO4 Test)
4.3.8. Alkaloids (Dragendorff Test)
4.3.9. Saponins
NaHCO3 Test
Salkowski Test for Saponins
Aromaticity (H2SO4- CH2O Test)
4.4. Cell Lines
4.4.1. Vero Cell Line
4.4.2. HeLa Cell Line
4.4.3. MDA-MB-231 Cell Line
4.5. Preparation of Working Solutions
4.6. Cytotoxicity Test with MTT(3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide)
4.7. DAPI Nucleus Labeling Assay
4.8. Morphological Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Health Topics: Cancer. Available online: https://www.who.int/topics/cancer/es/ (accessed on 15 March 2021).
- Globocan: All Cancers. Available online: https://gco.iarc.fr/today/data/factsheets/cancers/39-All-cancers-fact-sheet.pdf (accessed on 15 March 2021).
- Globocan: Source Mexico. Available online: https://gco.iarc.fr/today/data/factsheets/populations/484-mexico-fact-sheets.pdf (accessed on 15 March 2021).
- National Cancer Institute: Types of Cancer Treatment. Available online: https://www.cancer.gov/about-cancer/treatment/types (accessed on 10 May 2020).
- Karunamoorthi, K.; Jegajeevanram, K.; Vijayalakshmi, J.; Mengistie, E. Traditional Medicinal Plants: A Source of Phytotherapeutic Modality in Resource-Constrained Health Care Settings. J. Evid.-Based Complement. Altern. Med. 2013, 18, 67–74. [Google Scholar] [CrossRef]
- Seebaluck, R.; Gurib-Fakim, A.; Mahomoodally, F. Medicinal plants from the genus Acalypha (Euphorbiaceae) A review of their ethnopharmacology and phytochemistry. J. Ethnopharmacol. 2015, 159, 137–157. [Google Scholar] [CrossRef] [PubMed]
- Soladoye, M.O.; Sonibare, M.A.; Rosanwo, T.O. Phytochemical and morphometric analysis of the genus Acalypha Linn. (Euphorbiaceae). J. Appl. Sci. 2008, 8, 3044–3049. [Google Scholar] [CrossRef]
- Emeka, P.M.; Badger-Emeka, L.I.; Fateru, F. In vitro antimicrobial activities of Acalypha ornate leaf extracts on bacterial and fungal clinical isolates. J. Herb. Med. 2012, 2, 136–142. [Google Scholar] [CrossRef]
- Gurib-Fakim, A.; Gueho, J. Plantes Médicinal de Maurice, 2nd ed.; Edition deL’Ocean Indien: Beau Bassin-Rose Hill, Mauritius, 1996; pp. 71–77.
- Gurib-Fakim, A.; Brendler, J. Medicinal and aromatic plants of Indian Ocean Islands, 1st ed.; MedPharm: Stuttgart, Germany, 2004; pp. 87–89. [Google Scholar]
- Estrada-Castillón, E.; Villarreal-Quintanilla, J.Á.; Rodríguez-Salinas, M.M.; Encinas-Domínguez, J.A.; González-Rodríguez, H.; Figueroa, G.R.; Arévalo, J.R. Ethnobotanical Survey of Useful Species in Bustamante, Nuevo León, Mexico. Hum. Ecol. 2018, 46, 117–132. [Google Scholar] [CrossRef]
- Canales, M.; Hernández, T.; Rodríguez, M.; Flores, O.; Jiménez, E.; Hernández, L.; Hernández, M.; Trejo, N.; Ramírez, J.E.; Martínez, K. Evaluation of the antimicrobial activity of Acalypha monostachya Cav. (Euphorbiales: Euphorbiaceae). Afr. J. Pharm. Pharmacol. 2011, 5, 640–647. [Google Scholar] [CrossRef] [Green Version]
- Maiti, R.; González, R.H.; Kumari, A. Nutrient Profile of Native Woody Species and Medicinal Plants in Northeastern Mexico: A Synthesis. J. Bioprocess. Biotech. 2016, 6, 283. [Google Scholar]
- Maiti, R.; Rodriguez, H.G.; Degu, H.D.; Kumari, C.A.; Sarkar, N.C. Macro and Micronutrients of 44 Medicinal Plant Species Used Traditionally in Nuevo Leon, Mexico. Int. J. Bio-Resour. Stress Manag. 2016, 7, 1054–1062. [Google Scholar] [CrossRef]
- Ramírez, A.; García, G.; Werner, O.; Navarro-Pedreño, J.; Ros, R.M. Implications of the Phytoremediation of Heavy Metal Contamination of Soils and Wild Plants in the Industrial Area of Haina, Dominican Republic. Sustainability 2021, 13, 1403. [Google Scholar] [CrossRef]
- Macías, K.L.; Juárez, B.I.; Cárdenas, N.C.; Aguirre, J.R.; Jasso, Y. Evaluación de plantas tradicionalmente utilizadas en la desinfección de heridas. Rev. Mex. Cienc. Farm. 2009, 40, 5–10. [Google Scholar]
- Sanseera, D.; Niwatananun, W.; Liawruangrath, B.; Liawruangrath, S.; Baramee, A.; Trisuwan, K.; Pyne, S.G. Antioxidant and anticancer activities from aerial parts of Acalypha indica Linn. CMU J. Nat. Sci. 2012, 11, 157–168. [Google Scholar]
- Rajkumar, V.; Guha, G.; Ashok Kumar, R. Therapeutic potential of Acalypha fruticosa. Food. Chem. Toxicol. 2010, 48, 1709–1713. [Google Scholar] [CrossRef]
- Lim, S.W.; Ting, K.N.; Bradshaw, T.D.; Zeenathul, N.A.; Wiart, C.; Khoo, T.J.; Loh, H.S. Acalypha wilkesiana extracts induce apoptosis by causing single strand and double strand DNA breaks. J. Ethnopharmacol. 2011, 138, 616–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, J.H.-Y. Investigating In Vitro Anticancer Properties of Malaysian Rainforest Plants: Acalypha wilkesiana Mull, Arg. Archidendron ellipticum (Blume) Hassk. Duabanga grandiflora Walp. Pseuduvaria macrophylla (Oliv.) Merr. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 2014. [Google Scholar]
- Abubakar, I.B.; Lim, S.; Loh, H. Synergistic Apoptotic Effects of Tocotrienol Isomers and Acalypha wilkesiana on A549 and U87MG Cancer Cells Cancer is a group of disease caused by internal and external factors characterized by uncontrolled growth and spread of abnormal cells, resultant. Trop. Life Sci. Res. 2018, 29, 229–238. [Google Scholar] [CrossRef]
- El-Raey, M.A.; Mohamed, T.K.; El-Kashak, W.A.; Fayad, W.O. Phenolic constituents and biological activities of Acalypha wilkesiana forma tricolor muell arg seeds. Int. J. Pharmacogn. Phytochem. Res. 2016, 8, 386–392. [Google Scholar]
- Rascón-Valenzuela, L.; Jiménez-Estrada, M.; Velázquez-Contreras, C.; Garibay-Escobar, A.; Vilegas, W.; Campaner, L.; Robles-Zepeda, R.E. Chemical composition and antiproliferative activity of Acalypha californica. Ind. Crops Prod. 2015, 69, 48–54. [Google Scholar] [CrossRef]
- Rehab, A.; Hussein; El-Anssary, A.A. Plants Secondary Metabolites: The Key Drivers of the Pharmacological Actions of Medicinal Plants, Herbal Medicine, Philip F. Builders. Available online: https://www.intechopen.com/chapters/61866 (accessed on 16 May 2020).
- Noumedem, J.A.; Tamokou, J.; Teke, G.N.; Momo, R.C.; Kuete, V.; Kuiate, J.R. Phytochemical Analysis, Antimicrobial and Radical-scavenging Properties of Acalypha manniana Leaves; SpringerPlus: Berlin/Heidelberg, Germany, 2013; Volume 2, p. 503. [Google Scholar]
- Iniaghe, O.M.; Malomo, S.O.; Adebayo, J.O. Proximate Composition and Phytochemical Constituents of Leaves of Some Acalypha Species. Pak. J. Nutr. 2009, 8, 256–258. [Google Scholar] [CrossRef] [Green Version]
- Nazri, N.M.; Hazali, N.; Ibrahim, M.; Masri, M.; Ayob, M.K. Preliminary studies on Acalypha indica: Proximate analysis and phytochemical screening. Int. J. Pharm. Pharm. Sci. 2016, 8, 406–408. [Google Scholar]
- Evanjelene, V.K.; Natarajan, D. In Vitro Antioxidant and Phytochemical Analysis of Acalypha alnifolia Klein Ex Willd. IOSR J. Pharm. Biol. Sci. 2012, 1, 43–47. [Google Scholar] [CrossRef]
- Onocha, P.A.; Oloyede, G.K.; Afolabi, Q.O. Phytochemical investigation, cytotoxicity and free radical scavenging activities of non-polar fractions of Acalypha hispida (leaves and twigs). EXCLI J. 2011, 10, 1–8. [Google Scholar]
- Sepúlveda-Vázquez, J.; Torres-Acosta, F.; Sandoval-Castro, C.; Martínez-Puc, J.; Chan-Pérez, J.I. La importancia de los metabolitos secundarios en el control de nematodos gastrointestinales en ovinos con énfasis en Yucatán, México. J. Selva Andina Anim. Sci. 2018, 5, 79–95. [Google Scholar] [CrossRef]
- Reddy, T. Exploring the Anti-inflammatory and Anti-cancer compounds from the leaves of Acalypha indica. IOSR J. Pharm. Biol. Sci. 2012, 4, 1–7. [Google Scholar] [CrossRef]
- Moongkarndi, P.; Kosem, N.; Kaslungka, S.; Luanratana, O.; Pongpan, N.; Neungton, N. Antiproliferation, antioxidation and induction of apoptosis by Garcinia mangostana (mangosteen) on SKBR3 human breast cancer cell line. J. Ethnopharmacol. 2004, 90, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V.; Karaosmanoğlu, O.; Sivas, H. Anticancer Activities of African Medicinal Spices and Vegetables in Medicinal Spices and Vegetables from África, 1st. ed.; Academic Press: Dschang, Cameroon, 2017; pp. 271–297. [Google Scholar]
- Chekuri, S.; Panjala, S.; Anupalli, R.R. Cytotoxic activity of Acalypha indica L. hexane extract on breast cancer cell lines (MCF-7). J. Phytopharmacol. 2017, 6, 264–268. [Google Scholar] [CrossRef]
- Motadi, L.R.; Choene, M.S.; Mthembu, N.N. Anticancer properties of Tulbaghia violacea regulate the expression of p53-dependent mechanisms in cancer cell lines. Sci. Rep. 2020, 10, 12924. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.N.S.A.; Wahab, N.a.; AbdMalek, S.N. In Vitro Morphological Assessment of Apoptosis Induced by Antiproliferative Constituents from the Rhizomes of Curcuma zedoaria. Evid.-Based Complementary Altern. Med. 2013, 2013, 257108. [Google Scholar]
- Häcker, G. The morphology of apoptosis. Cell Tissue Res. 2000, 301, 5–17. [Google Scholar] [CrossRef]
- Majno, G.; Joris, I. Apoptosis, oncosis, and necrosis. An overview of cell death. Am. J. Pathol. 1995, 146, 3–15. [Google Scholar] [PubMed]
- Hengartner, M.O. Apoptosis: Corralling the corpses. Cell 2001, 104, 325–328. [Google Scholar] [CrossRef] [Green Version]
- Clarke, P.G. Developmental cell death: Morphological diversity and multiple mechanisms. Anat. Embryol. 1990, 181, 195–213. [Google Scholar] [CrossRef]
- Ziegler, U.; Groscurth, P. Morphological Features of Cell Death. Physiology 2004, 19, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Polo, R.A. The apoptosis/autophagy paradox: Autophagic vacuolization before apoptotic death. J. Cell Sci. 2005, 118, 3091–3102. [Google Scholar] [CrossRef] [Green Version]
- Soto-Domínguez, A.; Ballesteros-Elizondo, R.G.; Santoyo-Pérez, M.E.; Rodríguez-Rocha, H.; García-Garza, R.; Nava-Hernández, M.P.; Saucedo-Cárdenas, O. Peroxisomicine A1 (toxin T-514) induces cell death of hepatocytes in vivo by triggering the intrinsic apoptotic pathway. Toxicon 2018, 154, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Shubin, A.V.; Demidyuk, I.V.; Komissarov, A.A.; Rafieva, L.M.; Kostrov, S.V. Cytoplasmic vacuolization in cell death and survival. Oncotarget 2016, 7, 55863–55889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefkó, D.; Kúsz, N.; Barta, A.; Kele, Z.; Bakacsy, L.; Szepesi, Á.; Fazakas, C.; Wilhelm, I.; Krizbai, I.A.; Hohmann, J.; et al. Gerardiins A-L and Structurally Related Phenanthrenes from the Halophyte Plant Juncus gerardii and Their Cytotoxicity against Triple-Negative Breast Cancer Cells. J. Nat. Prod. 2020, 83, 3058–3068. [Google Scholar] [CrossRef]
- Hanane, M.; Mouhcine, E.; Ahmed, M.; Mohammed, E.; Laila, B.; Mohammed, A. Cytotoxicity of the Aqueous Extract and Organic Fractions from Origanum majorana on Human Breast Cell Line MDA-MB-231 and Human Colon Cell Line HT-29. Adv. Pharmacol. Pharm. Sci. 2018, 2018, 9. [Google Scholar]
- Mendoza, N.; Escamilla, E. Introduction to Phytochemicals: Secondary Metabolites from Plants with Active Principles for Pharmacological Importance, Phytochemicals—Source of Antioxidants and Role in Disease Prevention. 2018. Available online: https://www.intechopen.com/books/phytochemicals-source-of-antioxidants-and-role-in-disease-prevention/introduction-to-phytochemicals-secondary-metabolites-from-plants-with-active-principles-for-pharmaco (accessed on 8 June 2021).
- Kikugawa, K.; Hayatsu, H.; Ukita, T. Modifications of nucleosides and nucleotides. Biochim. Biophys. Acta 1967, 134, 221–231. [Google Scholar] [CrossRef]
- Popescu, N.C.; DiPaolo, J.A.; Amsbaugh, S.C. Integration sites of human papillomavirus 18 DNA sequences on HeLa cell chromosomes. Cytogenet Cell Genet 1987, 44, 58–62. [Google Scholar] [CrossRef]
- Hoppe, F.; Butz, K. Repression of endogenous p53 transactivation function in HeLa cervical carcinoma cells by human papillomavirus type 16 E6, human mdm-2, and mutant p53. J. Virol 1993, 67, 3111–3117. [Google Scholar] [CrossRef] [Green Version]
- Mur, C.; Martínez-Carpio, P.A.; Fernández-Montolí, M.E.; Ramon, J.M.; Rosel, P.; Navarro, M.A. Growth of MDA-MB-231 cell line: Different effects of TGF-beta (1), EGF and estradiol depending on the length of exposure. Cell Boil. Int. 1998, 22, 679–684. [Google Scholar] [CrossRef]
- Chen, X.; Thibeault, S. Effect of DMSO concentration, cell density and needle gauge on the viability of cryopreserved cells in three dimensional hyaluronan hydrogel. In Proceedings of the 35th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Osaka, Japan, 3–7 July 2013. [Google Scholar]
- Van Tonder, A.; Joubert, A.M.; Cromarty, A.D. Limitations of the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay when compared to three commonly used cell enumeration assays. BMC Res. Notes 2015, 8, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytoconstituents | Test | Observations | AE | ME | HE |
|---|---|---|---|---|---|
| Unsaturations | Potassium permanganate test | Brown precipitate | +++ | ++ | + |
| Phenols | Ferric chloride test | Green color | ++ | +++ | + |
| Terpenoids and steroids | Salkowski test | Reddish-brown ring formation | − | ++ | ++ |
| Coumarins and lactones | Sodium hydroxide test | Yellow color | ++ | ++ | + |
| Sesquiterpene lactones | Baljet’s test | Orange color | + | + | − |
| Flavonoids | Sulphuric acid test | Reddish color | + | + | ++ |
| Alkaloids | Dragendorff test | Orange precipitate | + | − | − |
| Saponins | Foam test | Presence of stable foam | +++ | + | + |
| Carbohydrates | Molisch´s test | Purple ring formation | +++ | ++ | + |
| Aromatic compounds | Formaline test | Red color | + | + | + |
| Carbonyl group | 2-4 dinitrophenylhydrazine test | Orange color | ++ | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guillén-Meléndez, G.A.; Villa-Cedillo, S.A.; Pérez-Hernández, R.A.; Castillo-Velázquez, U.; Salas-Treviño, D.; Saucedo-Cárdenas, O.; Montes-de-Oca-Luna, R.; Gómez-Tristán, C.A.; Garza-Arredondo, A.J.; Zamora-Ávila, D.E.; et al. Cytotoxic Effect In Vitro of Acalypha monostachya Extracts over Human Tumor Cell Lines. Plants 2021, 10, 2326. https://doi.org/10.3390/plants10112326
Guillén-Meléndez GA, Villa-Cedillo SA, Pérez-Hernández RA, Castillo-Velázquez U, Salas-Treviño D, Saucedo-Cárdenas O, Montes-de-Oca-Luna R, Gómez-Tristán CA, Garza-Arredondo AJ, Zamora-Ávila DE, et al. Cytotoxic Effect In Vitro of Acalypha monostachya Extracts over Human Tumor Cell Lines. Plants. 2021; 10(11):2326. https://doi.org/10.3390/plants10112326
Chicago/Turabian StyleGuillén-Meléndez, Gloria A., Sheila A. Villa-Cedillo, Raymundo A. Pérez-Hernández, Uziel Castillo-Velázquez, Daniel Salas-Treviño, Odila Saucedo-Cárdenas, Roberto Montes-de-Oca-Luna, Christian A. Gómez-Tristán, Aimé Jazmín Garza-Arredondo, Diana Elisa Zamora-Ávila, and et al. 2021. "Cytotoxic Effect In Vitro of Acalypha monostachya Extracts over Human Tumor Cell Lines" Plants 10, no. 11: 2326. https://doi.org/10.3390/plants10112326
APA StyleGuillén-Meléndez, G. A., Villa-Cedillo, S. A., Pérez-Hernández, R. A., Castillo-Velázquez, U., Salas-Treviño, D., Saucedo-Cárdenas, O., Montes-de-Oca-Luna, R., Gómez-Tristán, C. A., Garza-Arredondo, A. J., Zamora-Ávila, D. E., de Jesús Loera-Arias, M., & Soto-Domínguez, A. (2021). Cytotoxic Effect In Vitro of Acalypha monostachya Extracts over Human Tumor Cell Lines. Plants, 10(11), 2326. https://doi.org/10.3390/plants10112326