
Potential of Inulin-Fructooligosaccharides Extract Produced from Red Onion (Allium cepa var. viviparum (Metz) Mansf.) as an Alternative Prebiotic Product

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Inulin-FOSs Extract Obtained from Red Onions
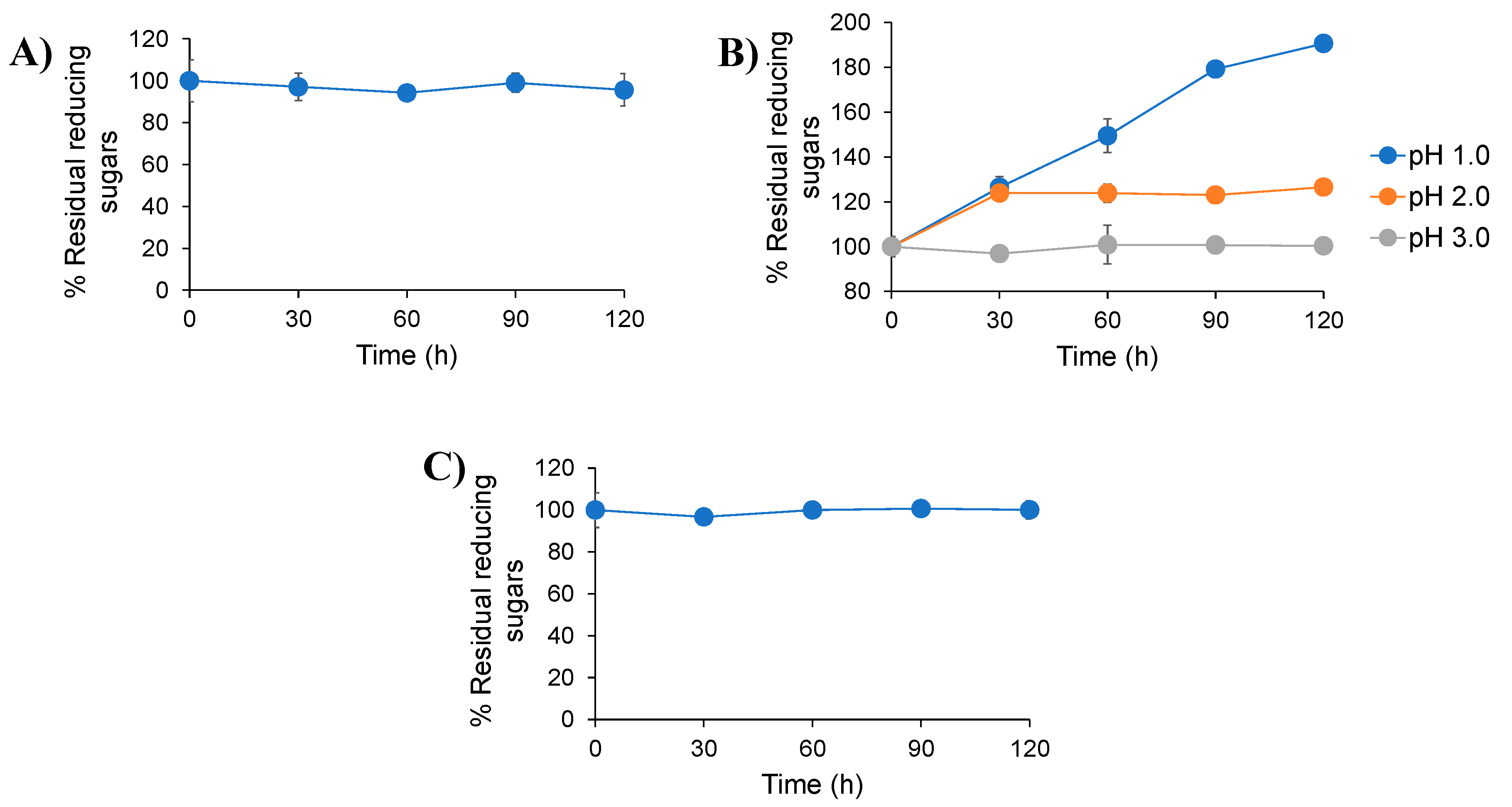
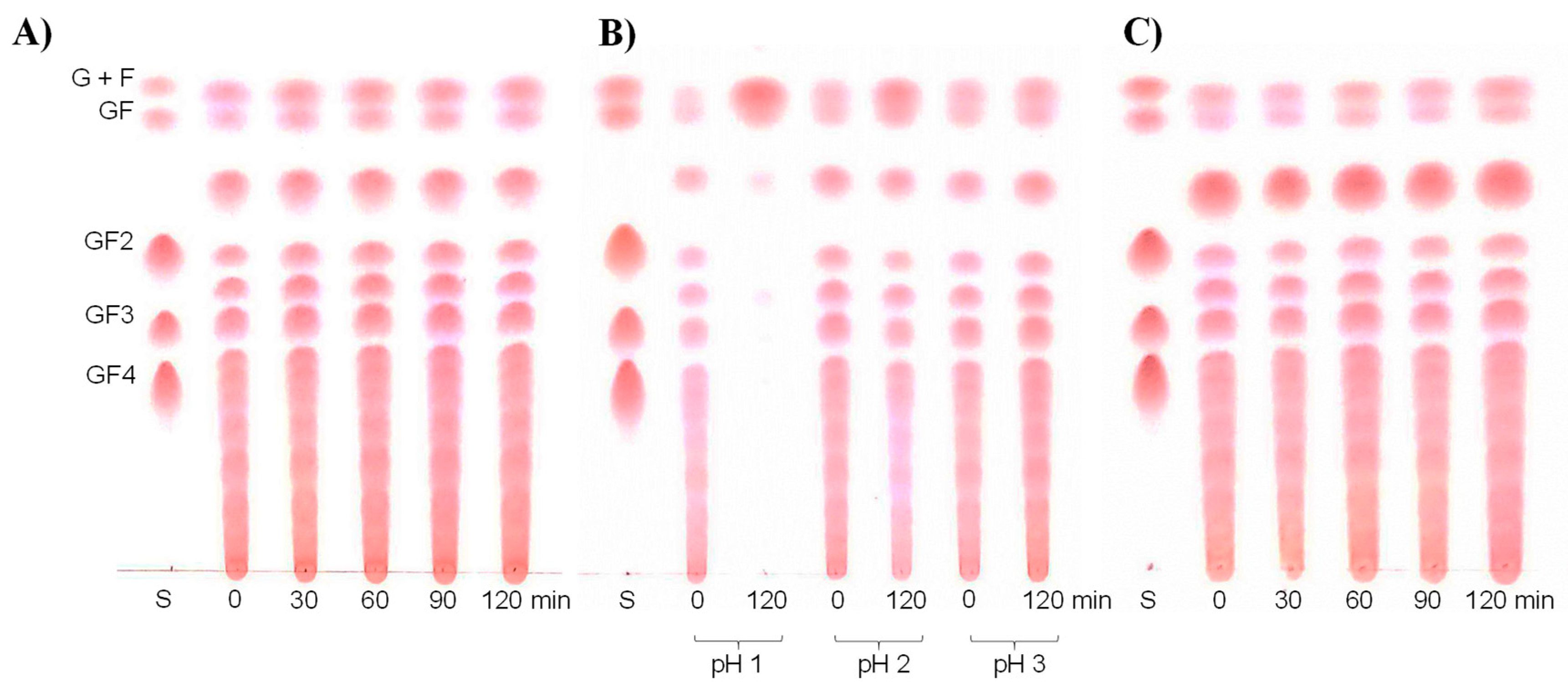
2.2. Digestibility of Inulin-FOSs Extract under Simulated Gastrointestinal Tract (GIT) Conditions
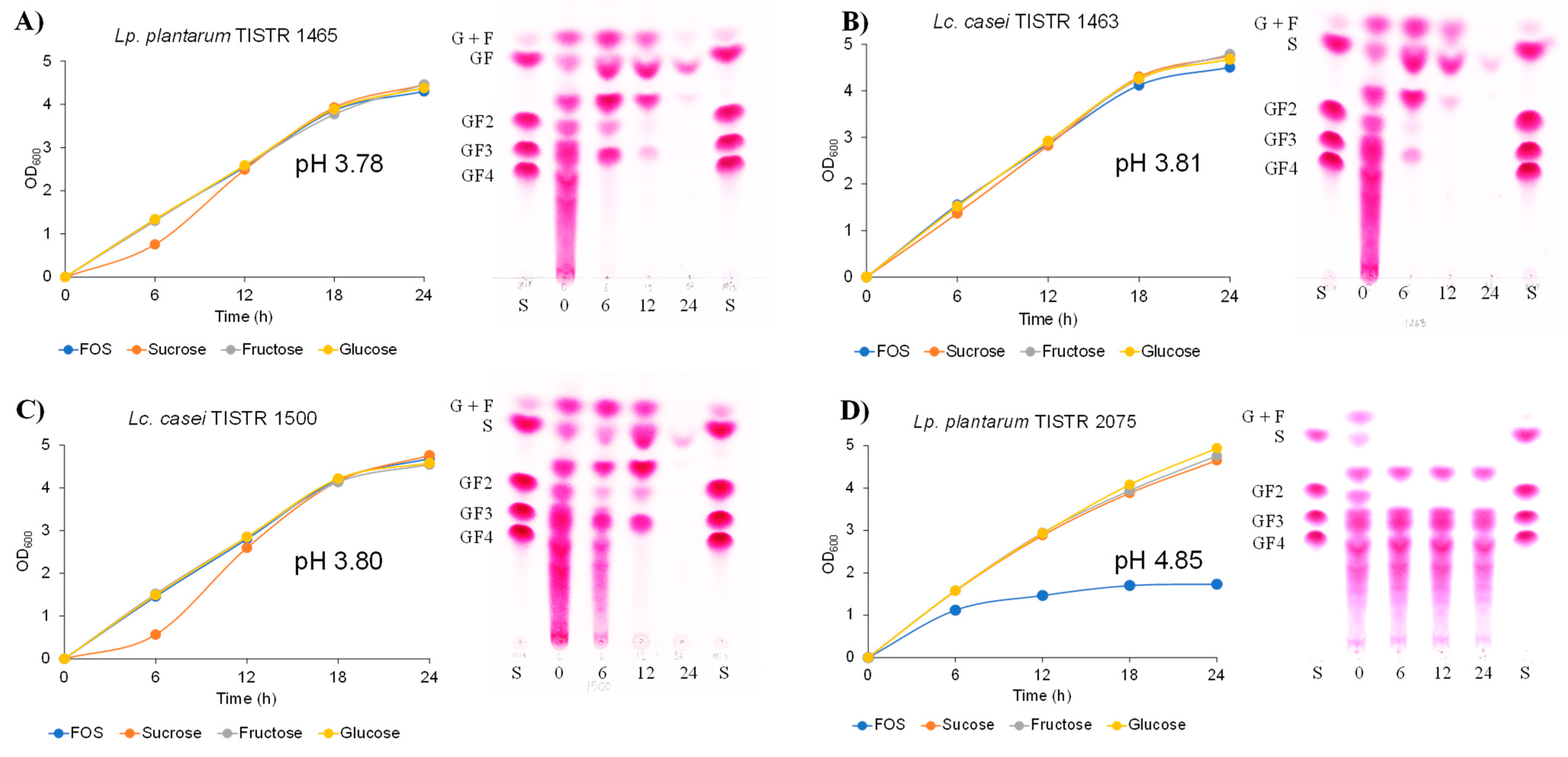
2.3. Inulin-FOSs Extract Fermentation and Inhibitory Effect on Foodborne Pathogenic Bacteria
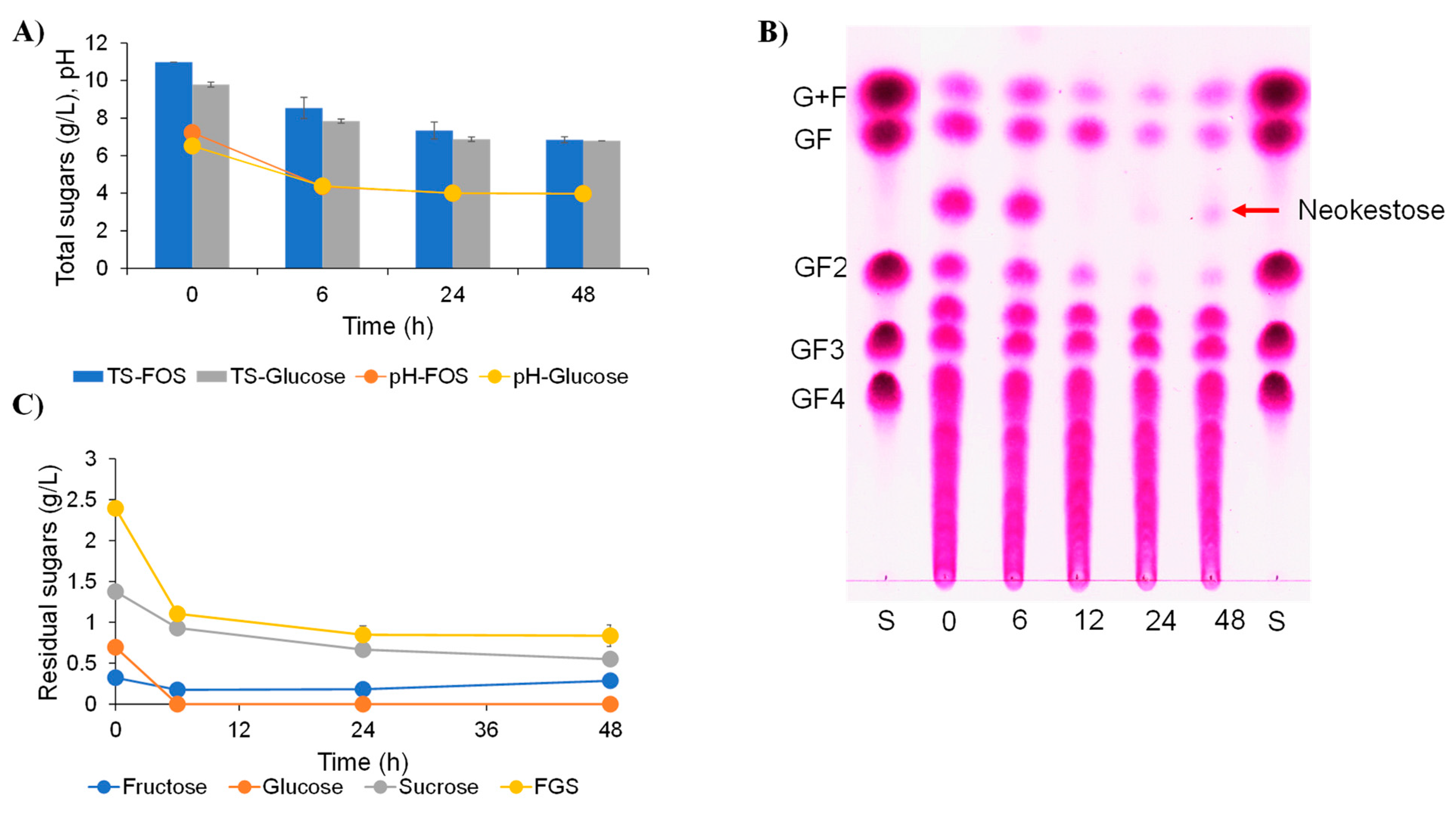
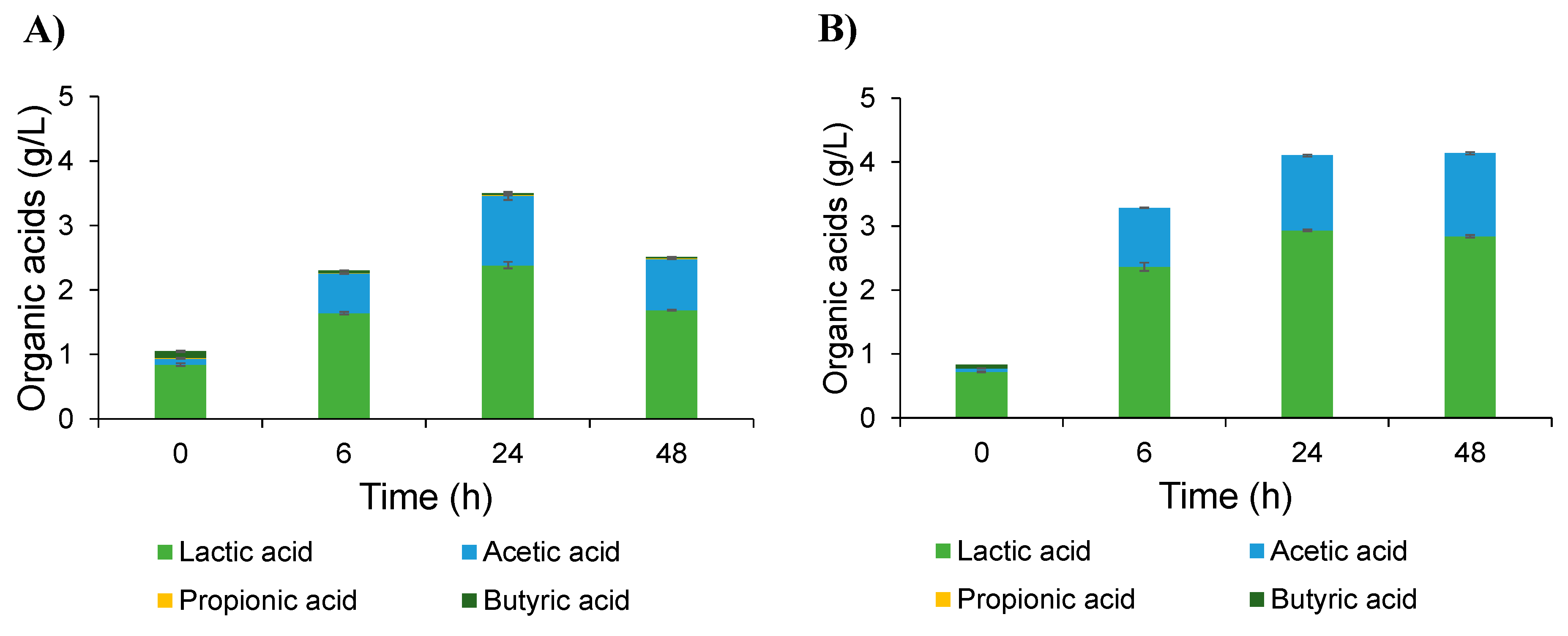
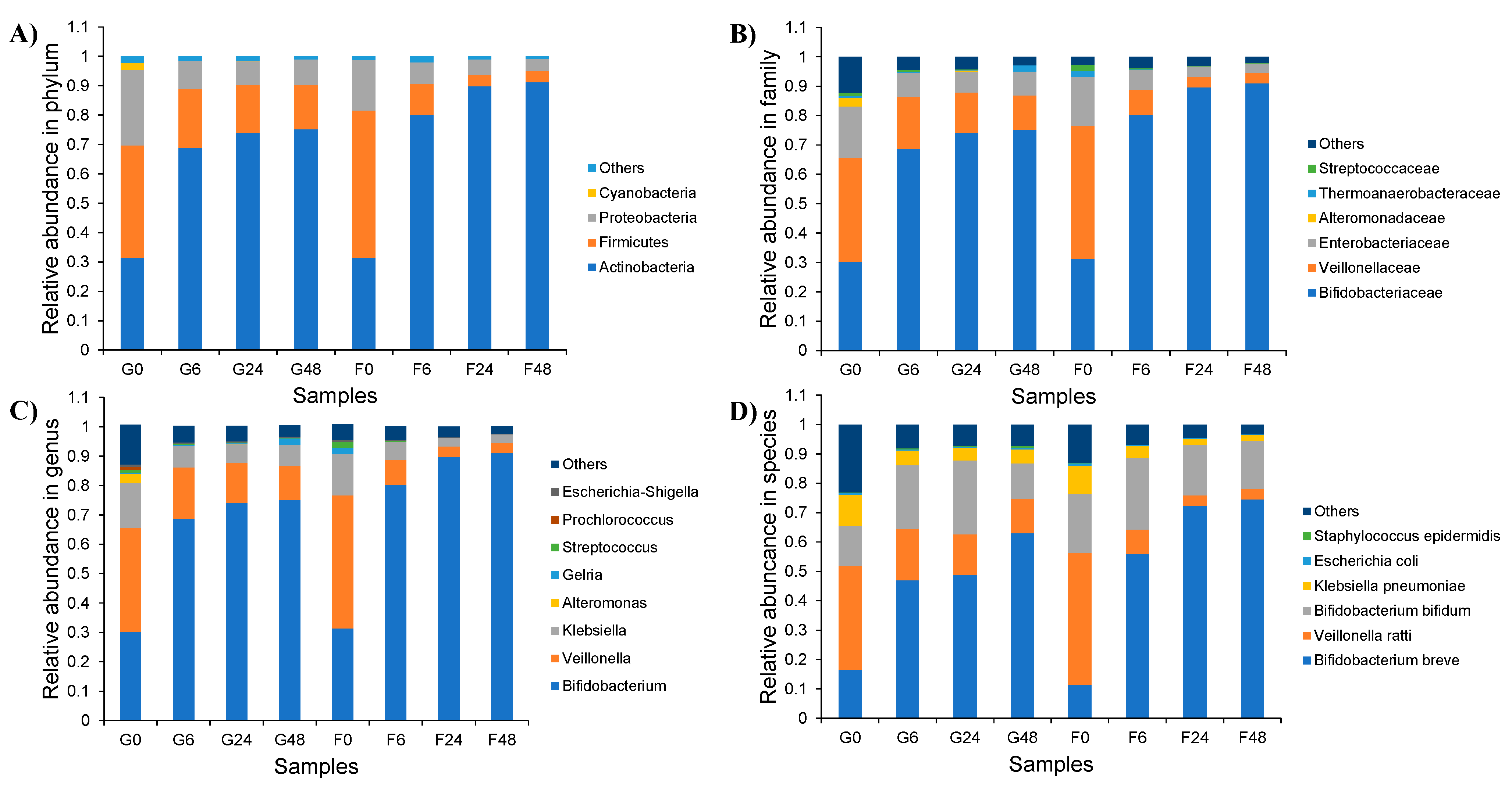
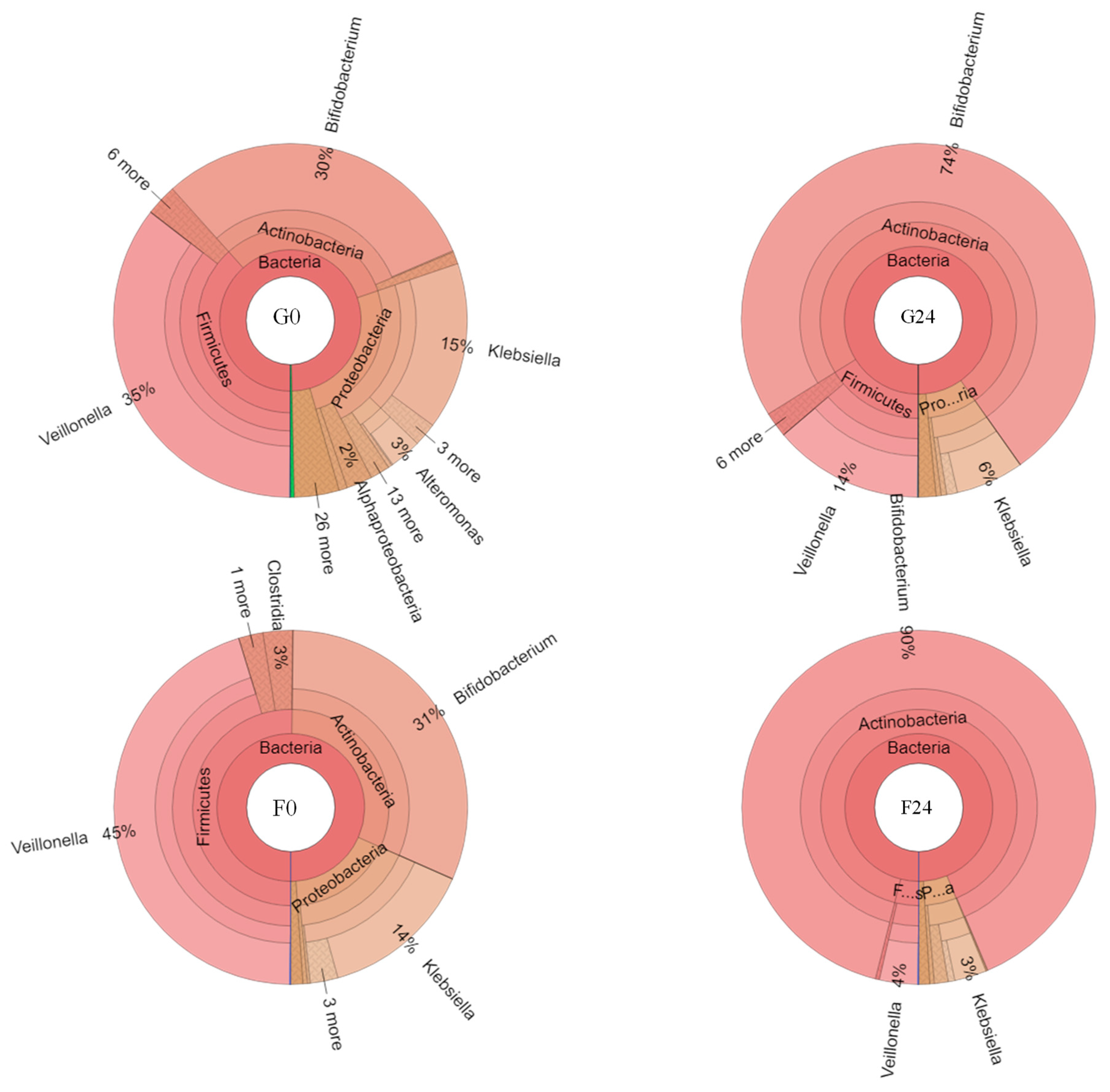
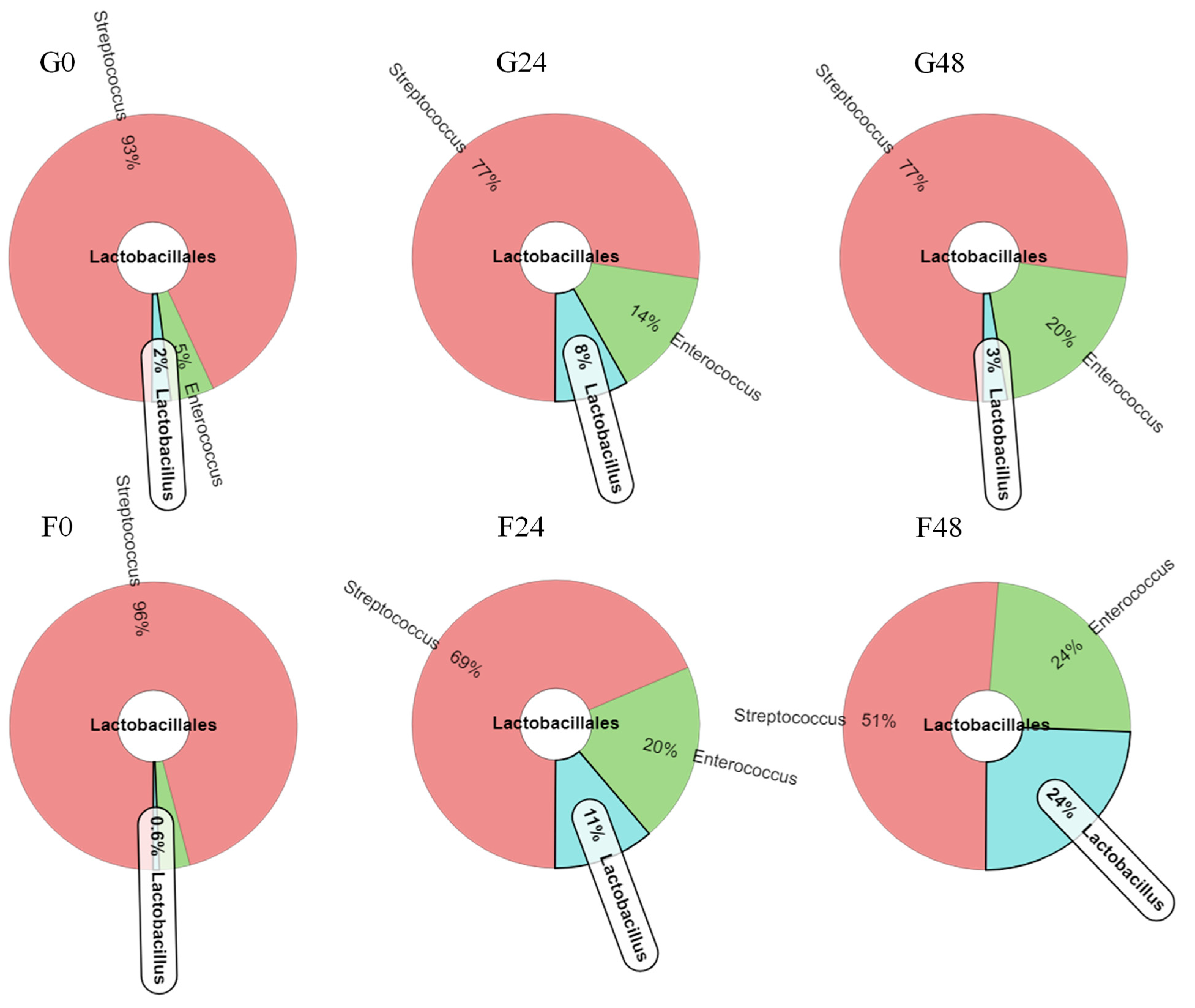
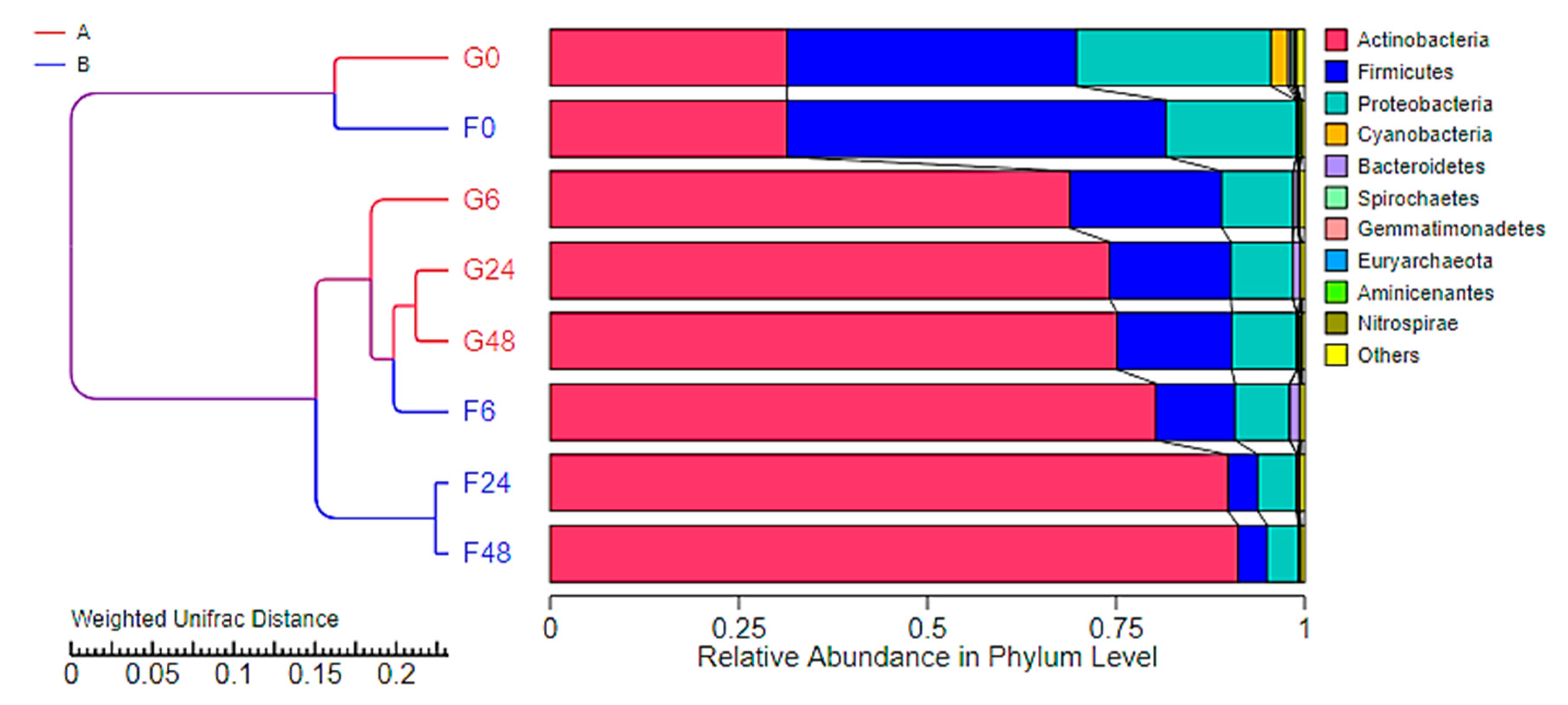
2.4. Effect of Inulin-FOSs Extract on Natural Mixed Culture
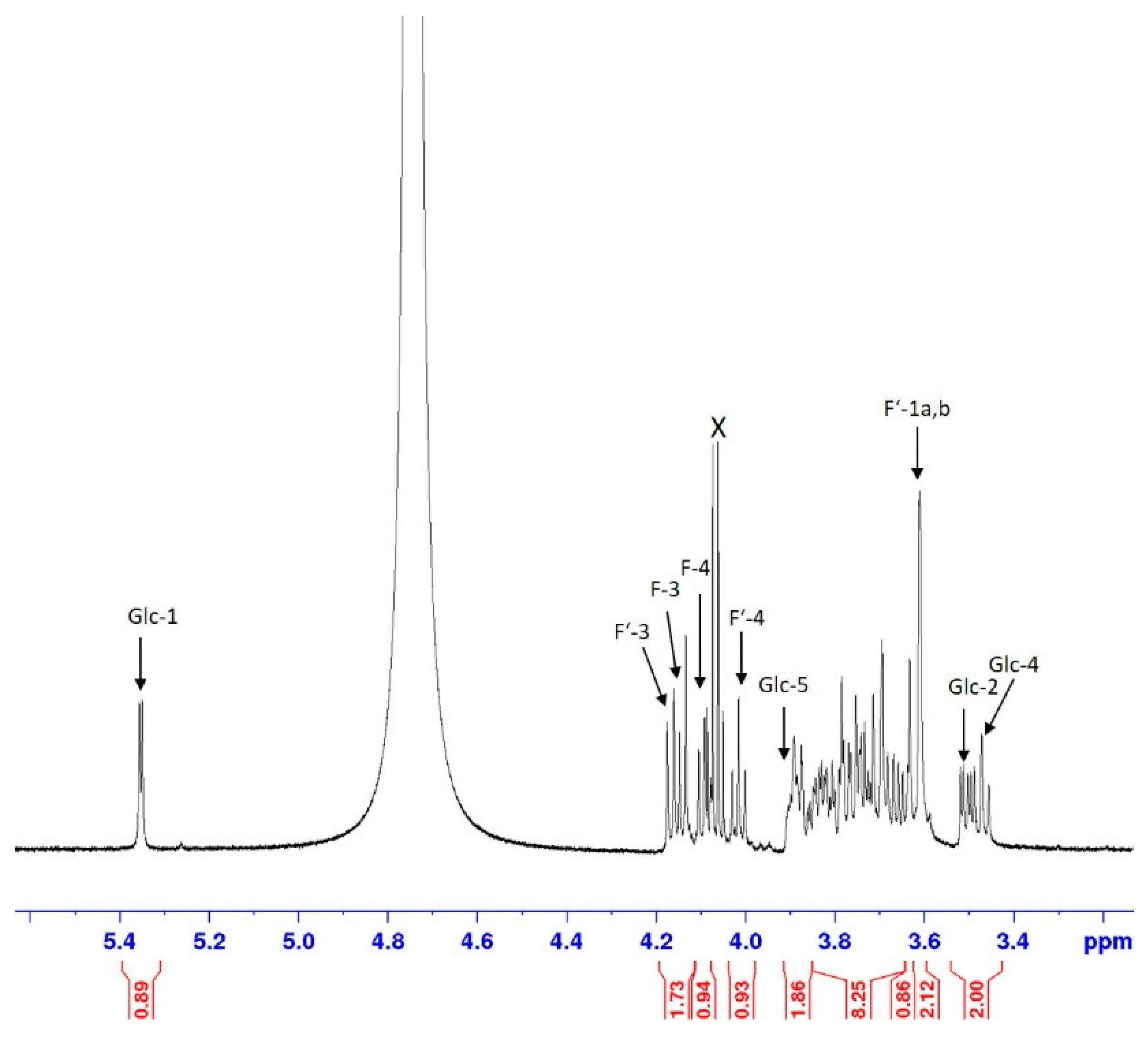
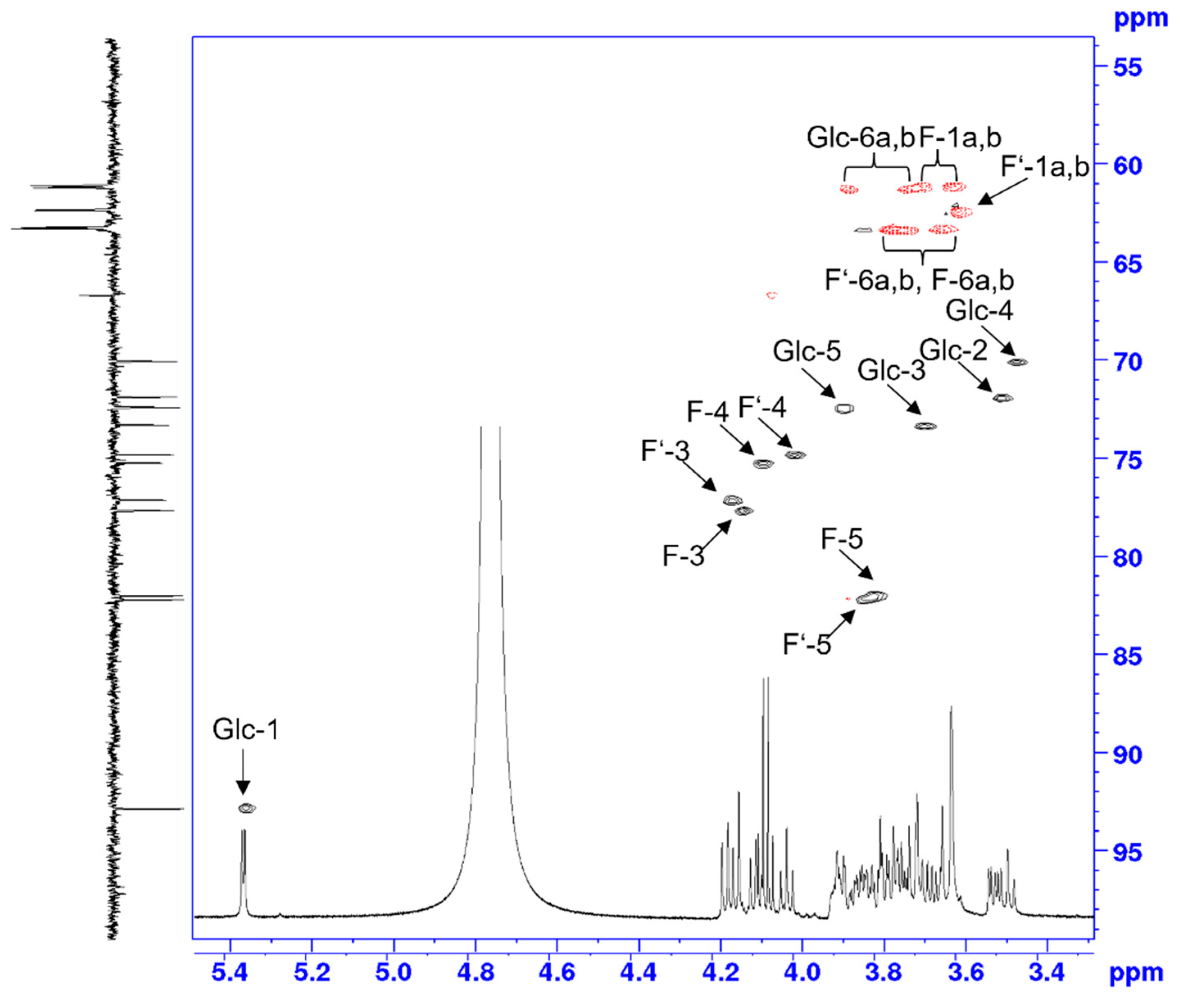
2.5. Identification of Unknown Sugar
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Microorganisms and Culture Conditions
4.3. Extraction and Standardization of Inulin-FOSs Extract
4.4. Simulated Oral Conditions
4.5. Simulated Gastric Conditions
4.6. Simulated Pancreatic Conditions
4.7. Fermentation of Inulin-FOSs by Various Probiotic Bacteria and Inhibitory Effect of Fermentation Broth
4.8. In Vitro Batch Culture Fermentation
4.9. Extraction of Genomic DNA and Amplicon Sequencing of 16S rRNA Gene
4.10. Bioinformatics
4.11. Thin Layer Chromatography and Isolation of Unknown Sugar
4.12. Structural Identification of Unknown Sugar by Nuclear Magnetic Resonance
4.13. Analytical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fujishima, M.; Furuyama, K.; Ishihiro, Y.; Onodera, S.; Fukushi, E.; Benkeblia, N.; Shiomi, N. Isolation and structural analysis in vivo of newly synthesized fructooligosaccharides in onion bulbs tissues (Allium cepa L.) during storage. Int. J. Carbohydr. Chem. 2009, 2009, 493737. [Google Scholar] [CrossRef] [Green Version]
- Kumar, C.G.; Sripada, S.; Poornachandra, Y. Chapter 14-Status and future prospects of fructooligosaccharides as nutraceuticals. In Role of Materials Science in Food Bioengineering; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: London, UK, 2018; pp. 451–503. [Google Scholar]
- Shiomi, N.; Benkeblia, N.; Onodera, S. The Metabolism of the Fructooligosaccharides in Onion Bulbs: A Comprehensive Review. J. Appl. Glycosci. 2005, 52, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Rastall, R.A. Functional oligosaccharides: Application and manufacture. Annu. Rev. Food Sci. Technol. 2010, 1, 305–339. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.-H.; Seo, J.M.; Kim, N.-H.; Arasu, M.V.; Kim, S.; Yoon, M.K.; Kim, S.-J. Variation of quercetin glycoside derivatives in three onion (Allium cepa L.) varieties. Saudi J. Biol. Sci. 2017, 24, 1387–1391. [Google Scholar] [CrossRef] [Green Version]
- Jaime, L.; Martín-Cabrejas, M.A.; Mollá, E.; López-Andréu, F.J.; Esteban, R.M. Effect of storage on fructan and fructooligosaccharide of onion (Allium cepa L.). J. Agric. Food Chem. 2001, 49, 982–988. [Google Scholar] [CrossRef]
- Wongputtisin, P. Selection of Oligosaccharides from Some Local Plants for Utilizing as Prebiotics. Master’s Thesis, Chiang Mai University, Chiang Mai, Thailand, 2003. [Google Scholar]
- Nanti, S.; Wongputtisin, P.; Chomsri, N.-o.; Deejing, S.; Niamsup, P. Primary prebiotic properties of Thai white sausage (Moo-yor) supplemented with fructooligosaccharides extracted from onion (Allium cepa L.) and chicory root. J. Agric. Sci. 2016, 33, 277–290. [Google Scholar]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Spacova, I.; Dodiya, H.B.; Happel, A.-U.; Strain, C.; Vandenheuvel, D.; Wang, X.; Reid, G. Future of probiotics and prebiotics and the implications for early career researchers. Front. Microbiol. 2020, 11, 1400. [Google Scholar] [CrossRef]
- Samanwong, T.; Ruangdechboonyarit, N.; Muangham, S.; Aeamsri, S. Survival of encasulated probiotics bacteria in yoghurt from corn milk, cow milk and soy milk as preserved at low temperature. Huachiew Chalermprakiet Sci. Technol. J. 2016, 2, 24–35. [Google Scholar]
- Chaipojjana, R. Production of Bioactive Powder by Double Emulsion. Master’s Thesis, Silpakorn University, Nakhon Pathom, Thailand, 2014. [Google Scholar]
- Lapsiri, W.; Nitisinprasert, S.; Wanchaitanawong, P. Lactobacillus plantarum strains from fermented vegetables as potential probiotics. Kasetsart J. Nat. Sci. 2011, 45, 1071–1082. [Google Scholar]
- Liu, J.; Waterhouse, A.L.; Chatterton, N.J. Proton and carbon chemical-shift assignments for 6-kestose and neokestose from two-dimensional n.m.r. measurements. Carbohydr. Res. 1991, 217, 43–49. [Google Scholar] [CrossRef]
- Rodríguez Galdón, B.; Tascón Rodríguez, C.; Rodríguez Rodríguez, E.M.; Díaz Romero, C. Fructans and major compounds in onion cultivars (Allium cepa). J. Food Compost. Anal. 2009, 22, 25–32. [Google Scholar] [CrossRef]
- Jurgiel-Malecka, G.; Gibszynska, M.; Nawrocka-Pezik, M. Comparison of chemical composition of selected cultivars of white, yellow and red onions. Bulg. J. Agric. Sci. 2015, 21, 2015. [Google Scholar]
- Roberfroid, M. Prebiotics: The concept revisited. J. Nutr. 2007, 137, 830s–837s. [Google Scholar] [CrossRef] [Green Version]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas, A.; Regal, P.; Vázquez, B.; Cepeda, A.; Franco, C.M. Short chain fatty acids commonly produced by gut microbiota influence Salmonella enterica motility, biofilm formation, and gene expression. Antibiotics 2019, 8, 265. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; O’Riordan, M.X.D. Regulation of bacterial pathogenesis by intestinal short-chain fatty acids. Adv. Appl. Microbiol. 2013, 85, 93–118. [Google Scholar] [PubMed] [Green Version]
- Mitsuoka, T.; Hidaka, H.; Eida, T. Effect of fructo-oligosaccharides on intestinal microflora. Nahrung 1987, 31, 427–436. [Google Scholar] [CrossRef]
- García-Curbelo, Y.; López, M.G.; Bocourt, R.; Collado, E.; Albelo, N.; Nuñez, O. Structural characterization of fructans from Agave fourcroydes (Lem.) with potential as prebiotic. Cuba. J. Agric. Sci. 2016, 49, 1–6. [Google Scholar]
- Kilian, S.; Kritzinger, S.; Rycroft, C.; Gibson, G.; du Preez, J. The effects of the novel bifidogenic trisaccharide, neokestose, on the human colonic microbiota. World J. Microbiol. Biotechnol. 2002, 18, 637–644. [Google Scholar] [CrossRef]
- Omori, T.; Ueno, K.; Muramatsu, K.; Kikuchi, M.; Onodera, S.; Shiomi, N. Characterization of recombinant β-fructofuranosidase from Bifidobacterium adolescentis G1. Chem. Cent. J. 2010, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Fang, Z.; Wang, H.; Zhai, Q.; Hang, F.; Zhao, J.; Zhang, H.; Lu, W.; Chen, W. Gene–Phenotype associations involving human-residential bifidobacteria (HRB) reveal significant species- and strain-specificity in carbohydrate catabolism. Microorganisms 2021, 9, 883. [Google Scholar] [CrossRef] [PubMed]
- Omori, T.; Ueno, K.; Kikuchi, M.; Onodera, S.; Shiomi, N. Properties of recombinant and β-fructofuranosidase from Bifidobacterium longum JCM1217. J. Appl. Glycosci. 2010, 57, 73–76. [Google Scholar] [CrossRef] [Green Version]
- Tochio, T.; Kadota, Y.; Tanaka, T.; Koga, Y. 1-kestose, the smallest fructooligosaccharide component, which efficiently stimulates Faecalibacterium prausnitzii as well as Bifidobacteria in humans. Foods 2018, 7, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultana, S.; Khan, A.; Safhi, M.M.; Alhazmi, H.A. Cough suppressant herbal drugs: A review. Int. J. Pharm. Sci. Invent. 2016, 5, 15–28. [Google Scholar]
- Grzelak, K.; Milala, J.; Król, B.; Adamicki, F.; Badełek, E. Content of quercetin glycosides and fructooligosaccharides in onion stored in a cold room. Eur. Food Res. Technol. 2009, 228, 1001–1007. [Google Scholar] [CrossRef]
- Ferreres, F.; Gil, M.I.; Tomás-Barberán, F.A. Anthocyanins and flavonoids from shredded red onion and changes during storage in perforated films. Int. Food Res. J. 1996, 29, 389–395. [Google Scholar] [CrossRef]
- Wu, J.S.; Chang, J.Y.; Chen, C.W.; Lin, M.T.; Sheu, D.C.; Lee, S.M. Neokestose suppresses the growth of human melanoma A2058 cells via inhibition of the nuclear factor-κB signaling pathway. Mol. Med. Rep. 2017, 16, 295–300. [Google Scholar] [CrossRef] [Green Version]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Kanpiengjai, A.; Mahawan, R.; Lumyong, S.; Khanongnuch, C. A soil bacterium Rhizobium borbori and its potential for citrinin-degrading application. Ann. Microbiol. 2016, 66, 807–816. [Google Scholar] [CrossRef]
- Unban, K.; Khatthongngam, N.; Pattananandecha, T.; Saenjum, C.; Shetty, K.; Khanongnuch, C. Microbial community dynamics during the non-filamentous fungi growth-based fermentation process of Miang, a traditional fermented tea of north Thailand and their product characterizations. Front. Microbiol. 2020, 11, 1515. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Kanpiengjai, A.; Lumyong, S.; Pathom-aree, W.; Khanongnuch, C. Starchy effluent from rice noodle manufacturing process as feasible substrate for direct lactic acid production by Lactobacillus plantarum S21. J. Korean Appl. Biol. Chem. 2014, 57, 217–220. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Red Onion (kg) | Extract Volume (L) | Total Carbohydrate (g/L) | FGS 1 (g/L) | Total Inulin-FOSs 2 (g/L) | Average DP 3 | Inulin-FOSs Yield 4 (g/100 g) |
|---|---|---|---|---|---|---|
| 20.0 | 10.0 | 98.00 ± 2.8 | 24.0 ± 0.38 | 74.0 ± 0.47 | 4.1 ± 0.13 | 3.7 ± 0.14 |
| Probiotic Bacteria | Fermentation Broth | E. coli | B. cereus | S. enterotica ser. Thyphimurium | St. aureus | L. monocytogenes |
|---|---|---|---|---|---|---|
| 1463 | Non-neutralized | 0.75 ± 0.07 a | 1.15 ± 0.07 a | 0.95 ± 0.07 | 0.95 ± 0.07 | 0.90 ± 0.00 a |
| Neutralized | 0.25 ± 0.07 b | 0.25 ± 0.07 b | 0.6 ± 0.14 | 0.70 ± 0.14 | 0.20 ± 0.00 b | |
| 1465 | Non-neutralized | 0.75 ± 0.07 | 0.60 ± 0.14 | 0.65 ± 0.07 a | 1.15 ± 0.07 a | 0.90 ± 0.00 a |
| Neutralized | 0.60 ± 0.00 | 0.45 ± 0.07 | 0.40 ± 0.00 b | 0.50 ± 0.00 b | 0.15 ± 0.07 b | |
| 1500 | Non-neutralized | 0.75 ± 0.07 a | 1.15 ± 0.21 a | 0.90 ± 0.00 | 1.00 ± 0.00 a | 0.85 ± 0.07 a |
| Neutralized | 0.35 ± 0.07 b | 0.45 ± 0.07 b | 0.70 ± 0.14 | 0.55 ± 0.07 b | 0.10 ± 0.00 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aisara, J.; Wongputtisin, P.; Deejing, S.; Maneewong, C.; Unban, K.; Khanongnuch, C.; Kosma, P.; Blaukopf, M.; Kanpiengjai, A. Potential of Inulin-Fructooligosaccharides Extract Produced from Red Onion (Allium cepa var. viviparum (Metz) Mansf.) as an Alternative Prebiotic Product. Plants 2021, 10, 2401. https://doi.org/10.3390/plants10112401
Aisara J, Wongputtisin P, Deejing S, Maneewong C, Unban K, Khanongnuch C, Kosma P, Blaukopf M, Kanpiengjai A. Potential of Inulin-Fructooligosaccharides Extract Produced from Red Onion (Allium cepa var. viviparum (Metz) Mansf.) as an Alternative Prebiotic Product. Plants. 2021; 10(11):2401. https://doi.org/10.3390/plants10112401
Chicago/Turabian StyleAisara, Jakkrit, Pairote Wongputtisin, Somkid Deejing, Chutamas Maneewong, Kridsada Unban, Chartchai Khanongnuch, Paul Kosma, Markus Blaukopf, and Apinun Kanpiengjai. 2021. "Potential of Inulin-Fructooligosaccharides Extract Produced from Red Onion (Allium cepa var. viviparum (Metz) Mansf.) as an Alternative Prebiotic Product" Plants 10, no. 11: 2401. https://doi.org/10.3390/plants10112401
APA StyleAisara, J., Wongputtisin, P., Deejing, S., Maneewong, C., Unban, K., Khanongnuch, C., Kosma, P., Blaukopf, M., & Kanpiengjai, A. (2021). Potential of Inulin-Fructooligosaccharides Extract Produced from Red Onion (Allium cepa var. viviparum (Metz) Mansf.) as an Alternative Prebiotic Product. Plants, 10(11), 2401. https://doi.org/10.3390/plants10112401