
Below-Ground Growth of Alpine Plants, Not Above-Ground Growth, Is Linked to the Extent of Its Carbon Storage

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Species
2.2. Sampling and Analysis of NSCs
2.3. Statistical Analysis
3. Results
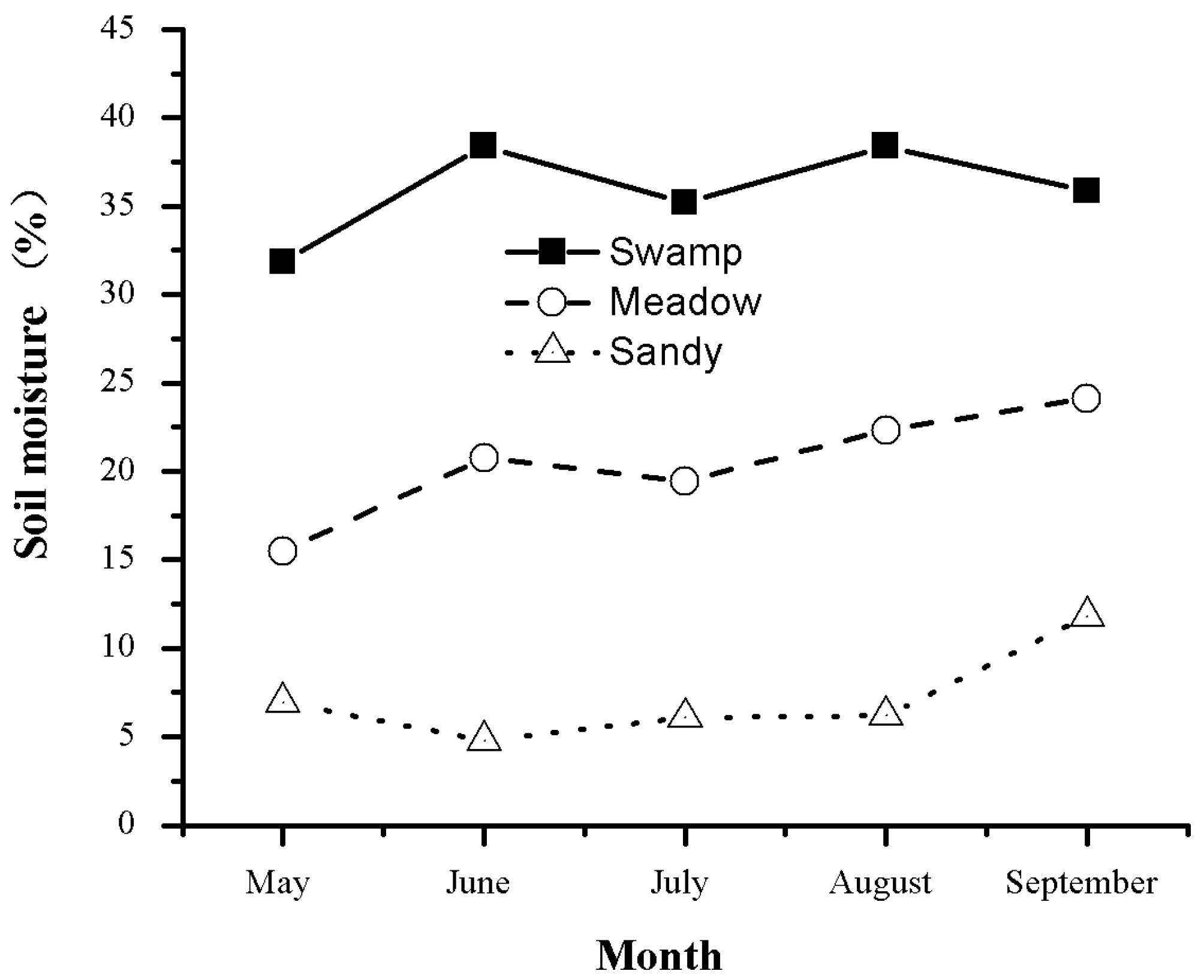
3.1. Seasonal Variation in Vegetation Biomass
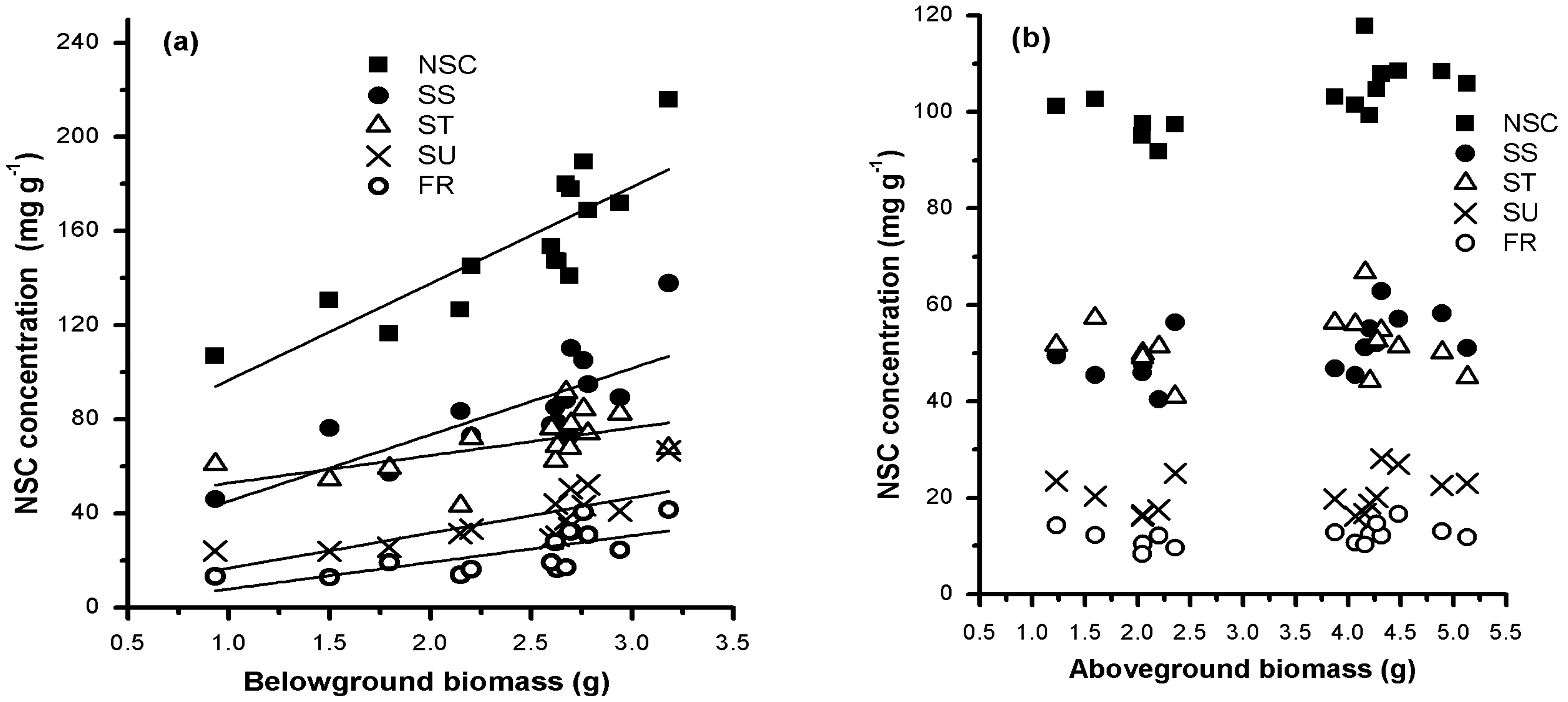
3.2. Relationships between Biomass and NSCs in Above- and Below-Ground Organs
3.3. Seasonal Variance Features of NSCs in Above- and Below-Ground Organs
3.4. Differences in NSCs Concentrations in Above- and Below-Ground Organs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Bahn, M.; Lattanzi, F.A.; Hasibeder, R.; Wild, B.; Koranda, M.; Danese, V.; Brüggemann, N.; Schmitt, M.; Siegwolf, R.; Richter, A. Responses of belowground carbon allocation dynamics to extended shading in mountain grassland. New Phytol. 2013, 198, 116–126. [Google Scholar] [CrossRef]
- Taeger, S.; Sparks, T.H.; Menzel, A.; Rennenberg, H. Effects of temperature and drought manipulations on seedlings of scots pine provenances. Plant Biol. 2015, 17, 361–372. [Google Scholar] [CrossRef]
- Sun, J.; Ma, B.; Lu, X. Grazing enhances soil nutrient effects: Trade-offs between aboveground and belowground biomass in alpine grasslands of the Tibetan Plateau. Land Degrad. Dev. 2018, 29, 337–348. [Google Scholar] [CrossRef]
- Bloom, A.; Chapin, F.; Mooney, H. Resource limitations in plants—An economic analogy. Annu. Rev. Ecol. Sys. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Brouwer, R. Functional equilibrium: Sense or nonsense? Neth. J. Agric. Sci. 1983, 31, 335–348. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life; Springer: Berlin/Heidelberg, Germany, 2003; pp. 201–214. [Google Scholar]
- Gibon, Y.; Bläsing, O.E.; Palacios-Rojas, N.; Pankovic, D.; Hendriks, J.H.M.; Fisahn, J.; Höhne, M.; Günther, M.; Stitt, M. Adjustment of diurnal starch turnover to short days: Depletion of sugar during the night leads to a temporary inhibition of carbohydrate utilization, accumulation of sugars and post-translational activation of ADP-glucose pyrophosphorylase in the following light period. Plant J. 2010, 39, 847–862. [Google Scholar]
- Millard, P.; Sommerkorn, M.; Grelet, G. Environmental change and carbon limitation in trees: A biochemical, ecophysiological and ecosystem. New Phytol. 2007, 175, 11–28. [Google Scholar] [CrossRef]
- Prescott, C.E.; Grayston, S.J.; Helmisaari, H.S.; Katovská, E.; Ostonen, I. Surplus carbon drives allocation and plant-soil interactions. Trends Ecol. Evol. 2020, 35, 1110–1118. [Google Scholar] [CrossRef]
- Dietze, M.C.; Sala, A.; Carbone, M.S.; Czimczik, C.I.; Mantooth, J.A.; Richardson, A.D.; Vargas, R. Nonstructural carbon in woody plants. Annu. Rev. Plant Biol. 2014, 65, 667–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mooney, H.A.; Billings, W.D. The annual carbohydrate cycle of alpine plants is related to growth. Am. J. Bot. 1960, 47, 594–598. [Google Scholar] [CrossRef]
- Hiltbrunner, E.; Arnaiz, J.; Körner, C. Biomass allocation and seasonal non-structural carbohydrate dynamics do not explain the success of tall forbs in short alpine grassland. Oecologia 2021, 197, 1063–1077. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S., III; Schulze, E.D.; Mooney, H.A. The ecology and economics of storage in plants. Annu. Rev. Ecol. Syst. 1990, 21, 423–447. [Google Scholar] [CrossRef]
- Gent, M.; Seginer, I. A carbohydrate supply and demand model of vegetative growth: Response to temperature and light. Plant Cell Environ. 2012, 35, 1274–1286. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, Y.; Chen, T.; Cui, X.; Wu, Q.; An, L. The effect and implication of human disturbances on an altitudinal variation of non-structural carbohydrates in Kobresia pygmaea. Acta Physiol. Plant 2014, 36, 2511–2519. [Google Scholar] [CrossRef]
- Bhattarai, P.; Zheng, Z.; Bhatta, K.P.; Adhikari, Y.P.; Zhang, Y. Climate-driven plant response and resilience on the Tibetan Plateau in space and time: A Review. Plants 2021, 10, 480. [Google Scholar] [CrossRef]
- Yang, Y.H.; Fang, J.Y.; Ji, C.J.; Han, W.X. Above- and belowground biomass allocation in Tibetan grasslands. J. Veg. Sci. 2009, 20, 177–184. [Google Scholar] [CrossRef]
- Kang, Y.Y.; Guo, S.R.; Li, J.; Duan, J.J. Effect of root applied 24-epibrassinolide on carbohydrate status and fermentative enzyme activities in cucumber (Cucumis sativus L.) seedlings under hypoxia. Plant Growth Regul. 2009, 57, 259–269. [Google Scholar] [CrossRef]
- Mccready, R.M.; Guggolz, J.; Silviera, V.; Owens, H.S. Determination of starch and amylose in vegetables. Anal. Chem. 1950, 22, 1156–1158. [Google Scholar] [CrossRef]
- Tolsma, A.D.; Read, S.M.; Tolhurst, K.G. Roots of Australian alpine plant species contain high levels of stored carbohydrates independent of post-fire regeneration strategy. Aust. J. Bot. 2007, 67, 959–980. [Google Scholar] [CrossRef]
- Stitt, M.; Zeeman, S.C. Starch turnover: Pathways, regulation and role in growth. Curr. Opin. Plant Biol. 2012, 15, 282–292. [Google Scholar] [CrossRef]
- Reich, P.B.; Luo, Y.; Bradford, J.B.; Poorter, H.; Perry, C.H.; Oleksyn, J. Temperature drives global patterns in forest biomass distribution in leaves, stems, and roots. Proc. Natl. Acad. Sci. USA 2014, 111, 13721–13726. [Google Scholar] [CrossRef] [Green Version]
- Niu, B.; Zeng, C.; Zhang, X.; He, Y.; Shi, P.; Tian, Y.; Feng, Y.; Li, M.; Wang, Z.; Wang, X.; et al. High below-ground productivity allocation of alpine grasslands on the Northern Tibet. Plants 2019, 8, 535. [Google Scholar] [CrossRef] [Green Version]
- Girardin, C.A.J.; Malhi, Y.; Aragão, L.E.O.C.; Mamani, M.; Huasco, W.H.; Durand, L.; Feeley, K.J.; Rapp, J.; Silva-Espejo, J.E.; Silman, M. Net primary productivity allocation and cycling of carbon along a tropical forest elevational transect in the Peruvian Andes. Glob. Chang. Biol. 2010, 16, 3176–3192. [Google Scholar] [CrossRef] [Green Version]
- Moser, G.; Leuschner, C.; Hertel, D.; Graefe, S.; Soethe, N.; Iost, S. Elevation effects on the carbon budget of tropical mountain forests (S Ecuador): The role of the belowground compartment. Global Chang. Biol. 2011, 17, 2211–2226. [Google Scholar] [CrossRef] [Green Version]
- Sulpice, R.; Pyl, E.T.; Ishihara, H.; Trenkamp, S.; Steinfath, M.; Witucka-Wall, H.; Gibona, Y.; Usadela, B.; Poreea, F.; Piquesa, C.M.; et al. Starch as a major integrator in the regulation of plant growth. Proc. Natl. Acad. Sci. USA 2009, 106, 10348–10353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.; Wang, G.; Hu, H.; Wang, Y. The variation of soil temperature and water content of seasonal frozen soil with different vegetation coverage in the headwater region of the Yellow River. China Environ. Geol. 2007, 54, 1755–1762. [Google Scholar] [CrossRef]
- Hu, H.; Wang, G.; Liu, G.; Li, T.; Ren, D.; Wang, Y.; Cheng, H.; Wang, J. Influences of alpine ecosystem degradation on soil temperature in the freezing-thawing process on Qinghai-Tibet Plateau. Environ. Geol. 2009, 57, 1391–1397. [Google Scholar] [CrossRef]
- Walker, D.J.; Romero, P.; Correal, E. Cold tolerance, water relations and accumulation of osmolytes in Bituminaria bituminosa. Biol. Plant. 2010, 54, 293–298. [Google Scholar] [CrossRef]
- Kontunen-Soppela, S.; Lankila, J.; Lahdesmaki, P.; Laine, K. Response of protein and carbohydrate metabolism of Scots pine seedlings to low temperature. J. Plant Physiol. 2002, 159, 175–180. [Google Scholar] [CrossRef]
- Luo, W.; Jiang, Y.; Lü, X.; Wang, X.; Li, M.; Bai, E.; Han, X.; Xu, Z. Patterns of plant biomass allocation in temperate grasslands across a 2500-km transect in northern China. PLoS ONE 2013, 8, e71749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sage, R.F.; Kubien, D.S. Quo vadis C4? An ecophysiological perspective on global change and the future of C4 plants. Photosynth. Res. 2003, 77, 209–225. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Meadow | Species |
|---|---|
| Alpine sandy grassland | Carex moorcroftii Falc. ex Boott (17), Astragalus mongholicus Bunge (13), Leontopodium alpinum Cass (40), Androsace tapete Maxim (15), Aster tongolensis Franch (21), Kobresia pygmaea C. B. Clarke (23), Microula sikkimensis (Clarke) Hems (9), Salvia prionitis Hance (33), Poa annua L. (12) |
| Typical alpine meadow | Same species listed as in alpine sandy grassland, the number of the species is 21, 23, 40, 15, 17, 14, 7, 45, 13, respectively. |
| Swamp meadow | Carex moorcroftii Falc. ex Boott (13), Astragalus mongholicus Bunge (13), Cremanthodium humile Maxim (8), Delphinium grandiflorum L. (13), Allium carolinianum DC. (7), Pedicularis tibetica Franch (14), Saussurea superba J. Anthony (20), Polygonum viviparum L. (7), Polygonum sibiricum Laxm (20) |
| Variables | Typical Alpine Meadow | Swamp Meadow | Alpine Sandy Grassland | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Below | Above | Correlation | Below | Above | Correlation | Below | Above | Correlation | |
| NSC | 12.53 | 8.53 | 0.901 ** | 22.53 | 5.99 | 0.681 * | 20.08 | 4.56 | 0.725 ** |
| SS | 7.64 | 5.58 | 0.994 ** | 31.44 | 15.20 | 0.798 * | 22.03 | 9.56 | −0.049 |
| ST | 18.90 | 12.99 | 0.818 ** | 14.59 | 9.99 | −0.211 | 24.28 | 12.08 | 0.832 ** |
| SU | 21.10 | 10.68 | 0.484 * | 23.99 | 13.12 | 0.450 * | 26.17 | 8.58 | −0.171 |
| FR | 23.72 | 21.50 | 0.978 ** | 48.02 | 7.37 | 0.694 * | 34.40 | 20.60 | 0.363 |
| Variables | Typical Alpine Meadow | Swamp Meadow | Alpine Sandy Grassland | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Below | Above | Ratio | Below | Above | Ratio | Below | Above | Ratio | |
| NSC | 156.6 ± 19.6 | 103.6 ± 8.8 | 1.50 ± 0.07 | 139.7 ± 32.1 | 100.7 ± 6.0 | 1.38 ± 0.18 | 167.3 ± 33.6 | 104.2 ± 4.7 | 1.59 ± 0.19 |
| SS | 82.0 ± 6.3 | 48.7 ± 2.7 | 1.68 ± 0.03 | 70.8 ± 22.3 | 49.0 ± 7.4 | 1.43 ± 0.21 | 102.3 ± 22.5 | 55.4 ± 5.3 | 1.86 ± 0.33 |
| ST | 74.6 ± 14.1 | 54.9 ± 7.1 | 1.35 ± 0.11 | 68.8 ± 10.0 | 51.8 ± 5.1 | 1.34 ± 0.18 | 65.0 ± 15.6 | 48.8 ± 6.0 | 1.32 ± 0.13 |
| SU | 32.3 ± 6.8 | 17.9 ± 1.9 | 1.79 ± 0.15 | 32.1 ± 7.7 | 19.1 ± 2.5 | 1.69 ± 0.25 | 48.9 ± 12.0 | 25.3 ± 2.1 | 1.95 ± 0.42 |
| FR | 18.0 ± 4.3 | 11.3 ± 2.4 | 1.58 ± 0.05 | 24.3 ± 11.6 | 12.1 ± 0.9 | 1.97 ± 0.59 | 29.3 ± 10.0 | 12.9 ± 2.7 | 2.28 ± 0.57 |
| Meadow (M) | Season (S) | Organ (O) | Interactions | |||
|---|---|---|---|---|---|---|
| (M-O) | (S-O) | (M-S) | ||||
| NSC | ns | *** | *** | ns | ** | ns |
| Soluble sugars | * | ns | *** | ns | ns | ns |
| Starch | ns | ** | *** | ns | ns | ns |
| Sucrose | *** | ns | *** | ns | ns | ns |
| Fructose | ns | ** | *** | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Chen, T.; Yun, H.; Chen, C.; Liu, Y. Below-Ground Growth of Alpine Plants, Not Above-Ground Growth, Is Linked to the Extent of Its Carbon Storage. Plants 2021, 10, 2680. https://doi.org/10.3390/plants10122680
Zhang Y, Chen T, Yun H, Chen C, Liu Y. Below-Ground Growth of Alpine Plants, Not Above-Ground Growth, Is Linked to the Extent of Its Carbon Storage. Plants. 2021; 10(12):2680. https://doi.org/10.3390/plants10122680
Chicago/Turabian StyleZhang, Youfu, Tuo Chen, Hanbo Yun, Chunyan Chen, and Yongzhi Liu. 2021. "Below-Ground Growth of Alpine Plants, Not Above-Ground Growth, Is Linked to the Extent of Its Carbon Storage" Plants 10, no. 12: 2680. https://doi.org/10.3390/plants10122680
APA StyleZhang, Y., Chen, T., Yun, H., Chen, C., & Liu, Y. (2021). Below-Ground Growth of Alpine Plants, Not Above-Ground Growth, Is Linked to the Extent of Its Carbon Storage. Plants, 10(12), 2680. https://doi.org/10.3390/plants10122680